蘋果熱激轉錄因子生物信息學及表達分析
2021-02-12 11:41:56王新亮
西南農業學報 2021年12期
關鍵詞:分析
王新亮
(1.濱州學院學報編輯部,山東 濱州 256603; 2.濱州學院 山東省黃河三角洲生態環境重點實驗室,山東 濱州 256603)
【研究意義】土壤鹽堿化嚴重影響植物的生長,黃河三角洲是中國甚至世界增長最快的河口三角洲之一[1],也是我國沿海鹽堿地高度集中的地區,約有50%的土地屬于鹽堿地[2]。蘋果(Malus×domesticaBorkh.)是世界四大水果(蘋果、葡萄、柑桔和香蕉)之一[3],也是我國種植面積和產量最多的水果,是果農重要的經濟來源,但土壤鹽堿化制約著蘋果種植業的發展[4]。【前人研究進展】熱激轉錄因子(Heat shock transcription factor,Hsf)對熱、干旱、鹽堿等多種非生物脅迫產生響應。研究表明Hsf在真核生物中高度保守[5],植物Hsf家族被分為A、B和C三個亞族[6-7]。植物中的Hsf至少含有 DNA 結合結構域(DBD)、寡聚化結構域(OD)兩個結構域和一個核定位信號基序(NLS),C端激活基序(AHA)是A亞族特有的,大部分 Hsf還含有核輸出信號(NES)結構域[7-8]。【本研究切入點】Hsf是一類廣泛存在于植物體內的轉錄因子,研究發現在番茄、水稻、小麥、擬南芥和棗中分別至少存在26 個[9]、26 個[10-11]、56 個[12]、24 個[5]和21 個[7]Hsf。HsfA3可能在高溫、干旱和鹽脅迫的信號轉導中發揮著重要作用[12-13]。HsfA2能調節番茄花藥熱脅迫保護機制的活性[14]。擬南芥HsfB1和HsfB2b能抑制熱誘導型Hsf的表達,但在熱脅迫條件下,他們同樣能誘導熱休克蛋白基因的表達,從而提高擬南芥的耐熱性[15]。水稻中的OsHsfC1b在ABA介導的耐鹽性中發揮著重要作用,而且它還參與滲透脅迫的響應[16]。【擬解決的關鍵問題】為了解蘋果Hsf的生物信息、表達特性和功能信息,利用蘋果基因組GDDH13 v1.1,通過Hsf結構域特異氨基酸序列檢索和關鍵詞搜索,篩選出36 個Hsf成員,并對其中33個含有完整結構域的成員進行了亞細胞定位、系統進化樹、序列分析、表達分析與功能預測,旨在分析蘋果Hsf的基本信息和生物功能,以期為進一步分析Hsf在調節蘋果響應熱、鹽堿、干旱等非生物脅迫中的分子機制提供參考。
1 材料與方法
1.1 材料與處理
實驗在濱州學院黃河三角洲生態環境重點實驗室苗圃中進行。首先利用0.2% KMnO4浸泡平邑甜茶種子30 min,再用流水洗凈,然后與細沙混和均勻放在 4 ℃的冰箱中層積 60 d左右。露白的種子被種在育苗盤,育苗盤裝有種植土(土、蛭石、沙的體積比為 1︰2︰3),發芽后用1/2 Hoagland 營養液( pH = 6.8 ± 0.2)澆灌,每隔5 d 1次。6~8 片真葉時,平邑甜茶苗用1/2 Hoagland 營養液(含有100 mmol/L NaCl、50 mmol/L Na2SO4、 50 mmol/L NaHCO3,pH =8)處理,葉片和新根分別在第 0、1、3、6 天被收集,除根0 d為2個重復,其他樣品均為3 個重復,經液氮處理后儲存于-80 ℃中。
1.2 RNA 提取與測序
用TRIzol(Invitrogen)法提取樣品總RNA。利用磁珠從總RNA中富集mRNA,再用打斷buffer使mRNA片段化,然后用隨機N6引物反轉錄合成第一條鏈,再形成雙鏈,最后進行PCR擴增;利用熱變性得到PCR產物的單鏈,然后環化形成環狀DNA文庫。用BGISEQ-500平臺(華大基因,深圳)進行測序,原始數據經純化得到高質量序列,用 HISAT 將序列比對到蘋果參考基因組Malus×domesticaGDDH13 Whole Genome v1.1(https://www.rosaceae.org/species/malus/malus_x_domestica/genome_GDDH13_v1.1)[17]。
1.3 蘋果Hsf轉錄因子篩選與鑒定
利用Hsf結構域特異氨基酸序列(Pfam登錄號:PF00447),在蘋果基因組 GDDH13 v1.1蛋白序列中進行BLAST并通過“Heat shock transcription factor”關鍵詞搜索篩選蘋果Hsf候選成員。候選Hsf蛋白通過 Pfam (http://pfam.xfam.org)[18]和NCBI Conserved Domain Database(CDD,https://www.ncbi.nlm.nih.gov/cdd)進行確認。
1.4 Hsf蛋白理化性質、染色體定位與共線性分析
Hsf的理化性質和亞細胞定位分別通過ExPASy(https://web.expasy.org/protparam/)[19]和WoLF PSORT(https://wolfpsort.hgc.jp/)進行分析。
Hsf基因的染色體定位用 TBtools 軟件的 Amazing Gene Location From GTF/GFF 工具分析;共線性分析用 Quick Run MCScanX Wrapper 和Text Merge For MCScanX工具分析,然后使用 Circle Gene View工具可視化。
1.5 Hsf基因的系統進化樹與序列分析
擬南芥Hsf的氨基酸序列從擬南芥信息數據庫(TAIR https://www.arabidopsis.org/)獲得。系統進化樹利用MEGA-X采用Neighbor-Joining (NJ)法構建,bootstrap 設置為 1000。利用MEME 5.3.0版本(http://memesuite.org/tools/meme)分析保守基序(數目設為10,寬度為6~90 aa)[20],通過 TBtools 軟件的Gene Structure View(Advanced)進行基因結構分析并可視化。
1.6 Hsf表達與功能預測
利用平邑甜茶的RNA-seq數據通過Excel繪制Hsf的表達熱圖,因為有些基因在根和葉片中的表達差異較大,所以先分別對根和葉片的Hsf表達量進行標準化后再繪制熱圖。利用AppleMDO數據庫(http://bioinformatics.cau.edu.cn/AppleMDO/)[21]以GDDH13 v1.1 homology withArabidopsis為參考進行基因本體(Gene Ontology,GO)分析,顯著水平設置為0.01。通過在線軟件 STRING[22](https://string-db.org)分析蛋白相互作用網絡,物種來源選擇蘋果(Malusdomestica)。
2 結果與分析
2.1 Hsf轉錄因子的篩選與鑒定
利用Hsf結構域氨基酸序列,在GDR數據庫蘋果基因組(MalusxdomesticaGDDH13 Whole Genome v1.1)進行BLAST并通過關鍵詞共檢索到37個蘋果候選Hsf,然后通過Pfam和NCBI Conserved Domain Database確認36個含有顯著Hsf結構域,其中有3個含有的Hsf結構域不完整,因此本文只對33個含有完整Hsf結構域的成員做了進一步分析。表1列出了這些Hsf基因的定位、編碼蛋白的氨基酸數量、分子量、等電點等理化性質。這些含有完整結構域的Hsf基因在除9號外的各個染色體上均有分布,它們的編碼蛋白的氨基酸數為189~530;分子量為21 703.71~58 972.96;等電點為4.55~8.58。通過亞細胞定位預測發現,除MD00G1095900和MD02G1171800可能定位于葉綠體/細胞核中,MD05G1313700 可能定位于細胞核/線粒體中;其他Hsf蛋白均定位于細胞核中。
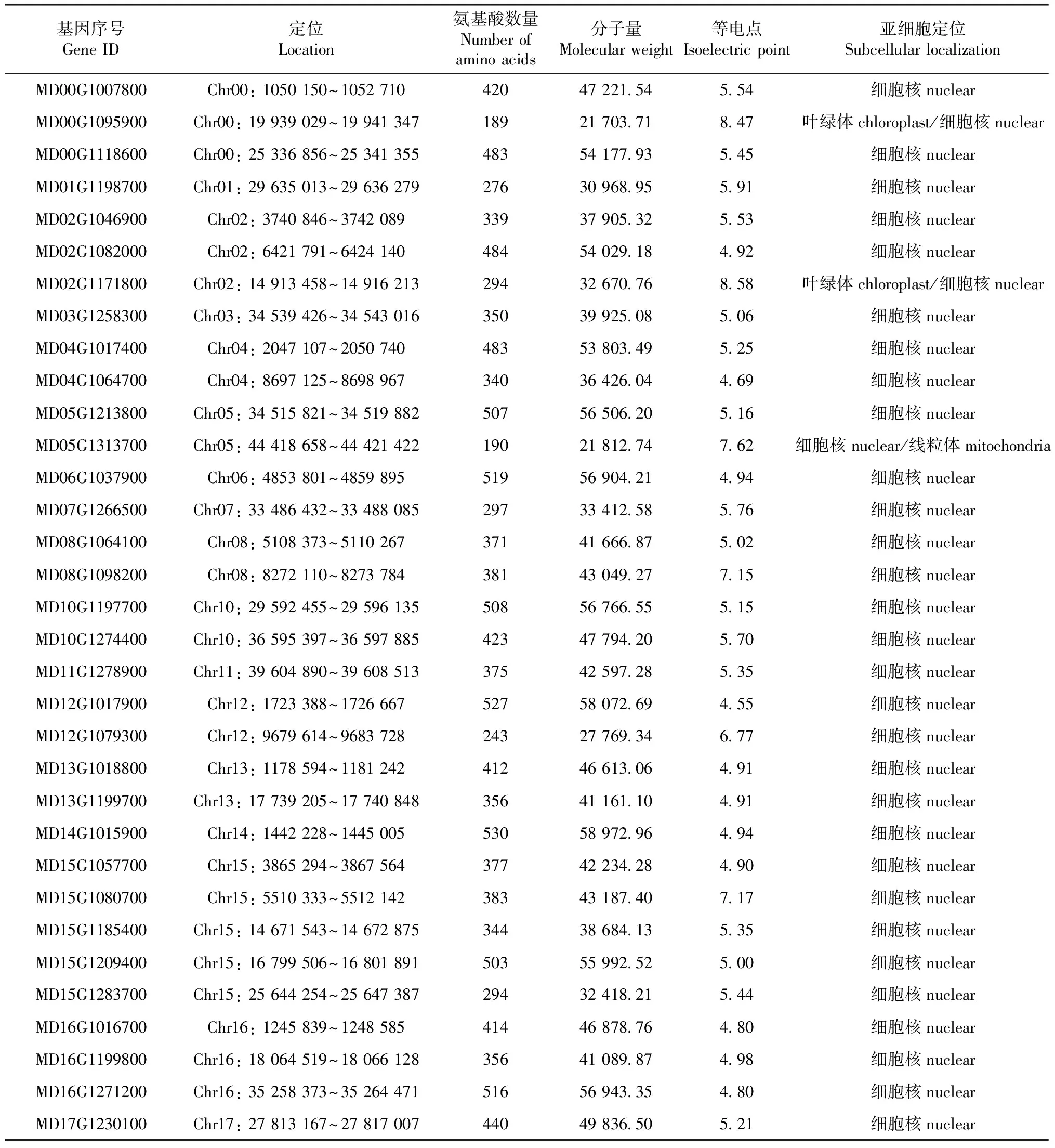
表1 蘋果 Hsf 基因及蛋白信息
2.2 Hsf基因的染色體定位與共線分析
串聯重復(Tandem duplication)、片段重復(segmental duplication)和轉位事件(transposition events)是基因家族表達的主要原因[23]。串聯重復是同一染色體上,200 kb內的片段上存在 2 個或2 個以上同一家族基因[24];而片段重復是不同染色體之間發生的基因重復事件[25]。基因組染色體定位分析顯示,不同染色體上的Hsf基因數量存在差異(圖1-A)。15號染色體含有5個含有完整結構域的Hsf基因,而7條染色體(1、3、6、7、11、14和17號染色體)上各只含有1個Hsf基因。因此,圖1-A顯示蘋果Hsf基因沒有串聯重復,但共線性分析(圖1-B)顯示有 16對共24個蘋果Hsf基因存在片段重復。
2.3 Hsf蛋白系統進化樹、基因結構和保守基序分析
利用MEGA-X軟件構建了蘋果與擬南芥Hsf家族成員的系統進化樹(圖2-A)。根據擬南芥的分組方法將這些Hsf家族成員分成11組,其中蘋果Hsf家族成員在A1組有4 個,A2組有2 個,A3組有2 個,A4組有3 個,A5組有2 個,A6/A7組有4 個,A9組有4 個;B1/B3組有5 個,B2組有3 個,B4組有2 個;C1組有2 個。基因結構分析顯示進化樹同一分支上基因的結構也相似(圖2-B),但成員較多的組中,基因結構差異較大。圖2-C顯示這33個Hsf蛋白均含基序1和基序3,進化樹同一分支上的Hsf蛋白的保守基序的數量和序列相似度也較高。
2.4 平邑甜茶Hsf基因在鹽堿脅迫下的表達分析
由圖3顯示,鹽堿脅迫1、3、6 d后的平邑甜茶幼苗葉和根中Hsf的表達情況,除MD15G1209400在葉片中沒有檢測到外,其他Hsf基因在根和葉中均檢測到一次及以上的表達。在鹽堿脅迫下,多數Hsf基因的表達顯著下調。只有MD05G1213800、MD10G1197700、MD15G1283700在根和葉中的表達同時上調。
2.5 蘋果Hsf蛋白GO分析
為了進一步了解蘋果Hsf的功能,利用AppleMDO數據庫的對這33個Hsf進行了GO功能分析。由表2顯示,共涉及到36個GO條目,其中生物過程25個、分子功能4個、細胞組成 6個。除生物過程中的6個條目(次生代謝過程、分解代謝過程、對內源性刺激的反應、對脅迫的反應、對非生物刺激的反應、對刺激的反應)參與的基因較少,其余條目均有33個Hsf參與。

表2 蘋果Hsf蛋白GO分析
2.6 蛋白相互作用網絡分析
挑選了表達量較高且差異較大的下調基因MD03G1258300和上調基因MD15G1283700,利用STRING 在線軟件構建了蛋白相互作用網絡(圖4),MD03G1258300與熱激蛋白(Hsp40)、超氧化物歧化酶銅分子伴侶(copper chaperone for superoxide dismutase,CCS)、蛋白磷酸酶2C(protein phosphatase 2C,PP2C)、 蔗糖非發酵-1-相關蛋白激酶γ調節亞基(SNF1-related protein kinase regulatory subunit gamma-like PV42a,SnRK1-PV42a)相互作用;MD15G1283700與熱激蛋白(Hsp90、Hsp83)、IAA氨基酸水解酶(IAA-amino acid hydrolase ILRl-like,ILR1)、堿性亮氨酸拉鏈(Basic region/leucine zipper 60,bZIP60)相互作用。
3 討 論
本研究利用Hsf保守結構域和關鍵詞檢索,鑒定了36 個蘋果Hsf家族成員,這與張國俊等[26]鑒定的數目不同,這可能是由于所使用的基因組版本不同引起的。本文只對33 個含有完整Hsf結構域的成員做了進一步分析。這33 個Hsf基因編碼的蛋白質含有189~530 個氨基酸,分子量為21 703.71~58 972.96,等電點為4.55~8.58,與棗和南瓜中Hsf類似[7,27]。
染色體定位和共線性分析顯示蘋果Hsf基因沒有串聯重復,但有16對共24 個蘋果Hsf基因存在片段重復,表明片段重復可能在蘋果的Hsf基因家族進化和擴增中發揮這重要作用。Velasco等[28]報道稱蘋果相對較近的一次基因組重復發生在至少5 千萬年前,導致蘋果染色體由9 條變成為17 條。蘋果基因組重復可能在基因家族擴張中起了重要作用。
系統進化樹可以為同源基因功能的預測提供參考。擬南芥65%以上的熱脅迫上調基因受AtHSFA1(AtHSFA1a、b、d、e)的調控,這 4個基因不僅在熱脅迫中起著重要作用,還調控擬南芥的生長和發育[29]。因此,與AtHSFA1a、b、d、e在同一分支上的MD05G1213800、 MD06G1037900、MD10G1197700和MD16G1271200可能也具有類似的功能。HsfB1和HsfB2b除了能提高擬南芥耐熱能力還具有提高抗病的能力[30]。MD02G1171800、MD04G1064700和MD15G1283700與上述2個基因較近,那么它們可能參與調節蘋果的抗病性。
研究表明Hsf不僅是熱應激防御反應的核心調節因子[31],還能調控植物對干旱、鹽堿等非生物脅迫的響應。本研究中除MD15G1209400在葉片中沒有表達外,其他Hsf基因的表達均受到鹽堿處理影響。鹽堿脅迫下,平邑甜茶大部分Hsf基因的表達下調,只有MD05G1213800、MD10G1197700、MD15G1283700的表達上調。表明蘋果Hsf家族成員可能在調控鹽堿脅迫響應中發揮重要作用。
GO功能分析顯示Hsf共涉及到36個GO條目,除DNA結合等GO條目外,還包括多種生物合成和代謝過程,以及對刺激和脅迫的反應生物過程,表明蘋果Hsf可能不僅在調控脅迫響應中起重要作用,而且調節其生長和發育。對MD03G1258300和MD15G1283700構建了蛋白相互作用網絡,MD03G1258300與Hsp40、CCS、PP2C、SnRK1-PV42a相互作用;MD15G1283700與Hsp90、Hsp83、ILR1、bZIP60相互作用。Hsf和Hsp是熱應激防御反應中的兩個重要成員。植物Hsf通過與‘nGAAnnTCCn’特別結合來調控Hsp基因的表達,Hsp能穩定染色質和膜,促進蛋白質修復,提高植物耐熱性并促進植物熱損傷的恢復[32]。Boyd 等[33]發現CCS1能通過提供一個銅離子及催化每個未成熟的銅—鋅超氧化物歧化酶(copper-zinc superoxide dismutase,SOD1)內二硫鍵的氧化來激活SOD1。PP2C可以通過多種信號轉導途徑來參與植物非生物脅迫響應[34]。在植物中,SnRK1是一類保守的絲氨酸/蘇氨酸激酶,由α-催化亞基和β、γ兩個調節亞基組成,PV42a是SnRK1的γ亞基[35]。SnRK1可以通過蔗糖合成酶(SS)控制碳水化合物代謝,能調節如3-羥基-3-甲基戊二酰—輔酶A還原酶(HMG-CoA還原酶)、磷酸蔗糖合成酶 (SPS)、硝酸還原酶(NR),而且它還在碳和氨基酸信號轉導中發揮重要作用[36]。有研究表明,在玉米原生質體中,HSFTF13(HsfA6b成員)的啟動子是bZIP60的一個靶標。bzip60突變體中,HSFTF13的表達降低并且高溫下的熱應激反應鈍化,Hsp基因在高溫下的表達不能正常上調[37]。因此,Hsf可以通過調節熱激蛋白,活性氧清除酶等多種功能基因來調節植物抗逆反應[38]。
4 結 論
本研究在蘋果基因組GDDH13 v1.1中檢索找到36個Hsf家族成員,然后對其中33個含有完整Hsf結構域的基因做了進一步分析。這些熱激轉錄因子基因編碼的蛋白質含有189~530個氨基酸數,分子量為21 703.71~58 972.96,等電點為4.55~8.58,它們多數定位于細胞核中。染色體定位和共線性分析顯示蘋果Hsf基因沒有串聯重復,但有16對共24個蘋果Hsf基因存在片段重復。在鹽堿脅迫下,多數Hsf基因的表達顯著下調。蘋果Hsf家族成員可能在調控鹽堿脅迫響應中發揮重要作用。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
