生草對北方山地桃園害蟲和天敵昆蟲數量動態的影響
2021-02-21 11:55:46張鵬飛
鄉村科技 2021年30期
關鍵詞:耕地
張 瑞 張鵬飛
(1.貴州省綠色食品發展中心,貴州 貴陽 550001;2.邢臺學院生物科學與工程學院,河北 邢臺 054001)
在果園內長期使用化學農藥,不僅會使害蟲產生抗藥性,殺傷害蟲的天敵昆蟲,引起害蟲再次猖獗,而且還會造成果品農藥殘留嚴重超標。隨著人們生活水平的日益提高,人們對高質量水果質量的要求也越來越高。因此,生產出綠色、高質量的水果成為當前迫切的需要。果園生草是一種病蟲害生物防治技術,基本原理是通過在果園內有選擇性地的種草,為園內的害蟲天敵昆蟲提供更加豐富的獵物、花蜜食物以及更適宜的越冬、越夏、棲息和避難場所,從而招引引來更多的昆蟲天敵,進而減施甚至不施化學農藥[1-3]。此外,采用生草技術還可改善地面小氣候環境,增加土壤墑情及有機質含量,最終有利于生產出高品質的水果[4-5]。
果園生草在國內外早有較多的研究,但多集中在蘋果園、梨園和棗園內,有關山區桃園生草的報道研究不是很多[6-10]。生草后果園內的生態系統,尤其是害蟲與其天敵昆蟲之間的關系發生了較大變化。因此,生草的種類以及果園所處的小氣候環境對生草效果有著重要影響。以往報道的桃園生草研究涉及的草種主要為白三葉(Trifoliumrepens)、夏至草(Lagopsissupina)等[11-12],而關于在北方淺山區桃園內混合種植黑麥草(Loliummultiflorum)、白三葉和紫花苜蓿(Medicagosativa)防治害蟲未有研究。本文對桃園內混合種植黑麥草、白三葉和紫花苜蓿防治害蟲進行了研究,為北方山區桃園害蟲的生物防治提供了一定的科學依據。
1 材料與方法
1.1 試驗地
試驗基地位于河北省石家莊市贊皇縣西部淺山區的花果山桃園,桃園總面積約達133.33余hm2,全年平均氣溫在10~20℃。試驗區域位于同一溝域內,面積約3.33hm2。參試驗桃樹的主要品種是為油蟠桃,樹齡4a,樹高2~3m,行株距為4m×2m,長勢良好。蚜蟲和葉蟬是該桃園的主要害蟲,調查期間正值葉蟬發生期。試驗地不施任何農藥。
1.2 試驗方法
1.2.1 生草試驗:于2019年4月,在生草地(圖1,處理組)樹下按3∶2∶2的比例以混合撒播的方式混種黑麥草、白三葉和紫花苜蓿3種牧草種子,清耕地(圖1,對照組)內不種植任何植物,并及時清除雜草。生草地設置了3個重復地塊,清耕地設置了2個重復地塊,每個重復地塊約2668m2。生草地與清耕地試驗區域的至少間隔200m。生草成坪后施肥以磷鉀肥為主,每667m2施肥10~20kg。在春夏季節,草的生長量較大,應適當增加氮磷鉀肥的用量,干旱時及時澆灌。生草成坪前要及時進行人工除草;生草成坪后,由于其抑制雜草的能力增強,一般不再進行除草。在草高30cm時開始刈割,全年刈割3~5次。刈割時,白三葉留1~2個分枝,禾本科草需留心葉。每次刈割時不可一次性割完,應順行留一部分,等割過的草長出后再刈割。生草地與清耕地內其他農事操作均保持一致。

圖1 生草地與清耕地
1.2.2 樹下地面昆蟲調查:采用五點取樣法在地面植被上調查昆蟲情況,每個采樣點的面積約60m2。調查時邊前進邊掃網,共掃10個來回、20網次,掃網寬度1m左右,掃網調查由專人進行。對于不明確的物種,捕獲后用75%的酒精浸泡,帶回室內進行鏡檢計數。每隔7~10d調查1次,調查多在9:00左右開始。調查區間從2019年6月29日開始到2019年9月29日結束。
1.2.3 樹冠昆蟲調查:在上述的每個采樣點內,選取1棵桃樹進行樹冠昆蟲調查,所選桃樹與地面調查區域有一定間隔。樹冠調查與地面調查同時進行。調查時環繞桃樹進行連續掃網(網長76.4cm,直徑32.8cm),共掃網10個來回、20網次,掃網調查由專人進行。其他同地面調查。
1.3 數據處理
利用GraphPadPrism6.0數據處理軟件對收集的數據進行處理,繪制種群動態圖(見圖2、圖3)。。

圖2 樹下植食性昆蟲與捕食性天敵的數量動態
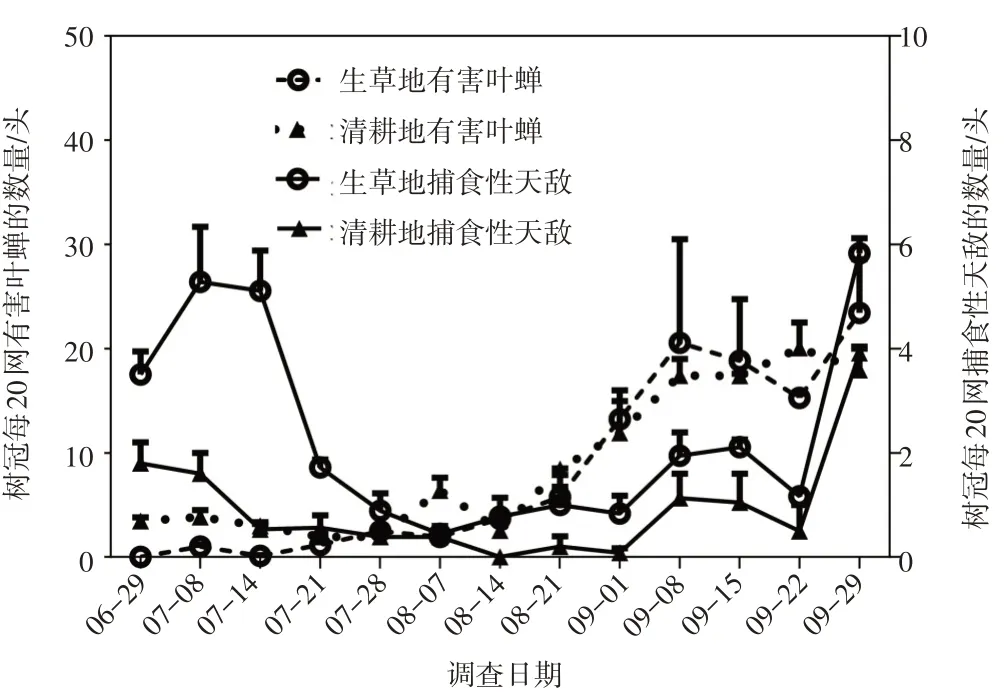
圖3 樹冠有害葉蟬與樹冠捕食性天敵的數量動態
2 結果與分析
2.1 樹下植食性昆蟲與捕食性天敵的種群動態
如圖2所示,對桃樹無害的植食性昆蟲條沙葉蟬(Psanrmotettixstrintus)在生草地的數量明顯多于清耕地。生草地內條沙葉蟬主要出現在7—8月份,并在7月中旬達到峰值,之后一直處于較低水平。對桃樹無害的植食性昆蟲苜蓿斑蚜(Therioaphistrifolii)在生草地從8月中旬開始出現,之后逐漸增多,清耕地內的苜蓿斑蚜數量明顯少于生草地。生草地內的捕食性天敵異色瓢蟲(Harmoniaaxyridis)、龜紋瓢蟲(Propylaeajaponica)、日本通草蛉(Chrysoperlanipponensis)和東亞小花蝽(Oriussauteri)混合種群的數量一直高于清耕地,并在7月下旬和9月底達到峰值。這與植食性昆蟲條沙葉蟬和苜蓿斑蚜分別在7月中旬和9月下旬達到峰值高度一致,表明生草地植食性昆蟲增多很可能是捕食性天敵增多的重要原因。
2.2 樹冠害蟲與樹冠捕食性天敵的種群數量動態
如圖3所示,生草地樹冠上的捕食性天敵異色瓢蟲、龜紋瓢蟲、日本通草蛉和東亞小花蝽混合種群的數量一直高于清耕地,并且種群數量的波動趨勢基本與樹下(圖2)一致。上述結果表明,生草地樹冠天敵昆蟲增多很可能與樹下天敵昆蟲增多有關。經初步鑒定,本地區桃樹上的主要害蟲為塔葉蟬族(Zyginellini)的葉蟬,這些樹冠上有害葉蟬的數量在清耕地總體高于生草地(圖3)。生草地內樹冠捕食性天敵的數量在7月上中旬處于較高水平,而此時樹冠上葉蟬的數量很低,之后樹冠混合天敵的數量也開始下降,至7月底降至最低水平;進入8月份,葉蟬的數量開始上升,到9月上旬達最高峰,此時天敵數量也開始上升,并在9月底也達到了最高峰,顯然,捕食性天敵與葉蟬間呈現出較為典型的互為消長的動態關系(圖3)。同樣在清耕地,天敵與葉蟬間的動態關系基本與生草地一致(圖3)。上述結果顯示,樹冠天敵對樹冠有害葉蟬具有一定的控制作用。
3 討論
生草地樹下捕食性天敵數量的增加很可能與地面的植食性昆蟲增多有關。條沙葉蟬的寄主多為一些禾本科雜草,對果樹無害,是果園內捕食性天敵的獵物之一。本試驗結果顯示,條沙葉蟬在生草地較多,在清耕地幾乎沒有。生草地中的條沙葉蟬主要出現在7—8月份,這可能是生草地樹下捕食性天敵在7月份顯著高于清耕地的主要原因之一。苜蓿斑蚜以苜蓿為寄主,對桃樹無害,也是園內捕食性天敵的獵物。上述結果顯示:清耕園內幾乎未發生苜蓿斑蚜;而在生草地,苜蓿斑蚜從8月中旬出現后,數量逐漸增多,這可能是樹下捕食性天敵在9月份升高的主要原因之一。
本文研究結果顯示,樹冠上捕食性天敵種群動態變化趨勢與樹下的高度一致,均在7月份和9月份相對較高,在其他時間較低,表明樹下天敵的數量可能對樹冠上的有直接影響。瓢蟲、草蛉和小花蝽是園內的主要捕食性天敵,這些天敵昆蟲均具有較強的擴散能力,易在樹冠與樹下間進行遷徙。因此,樹下的天敵數量增多后可遷徙至樹上,從而而使樹冠天敵增多。諶愛東等的研究表明,在蘋果園生草可顯著增加樹下以及樹冠上蚜蟲天敵的數量,并且樹冠天敵數量與樹下天敵數量之間呈正相關關系,與本文研究結果基本一致[13]。
本文結果顯示,在生草地和清耕地內,當葉蟬的數量較低時,天敵數量逐漸下降,而當葉蟬數量開始上升時,捕食性天敵緊隨其后,數量也在逐漸上升,二者呈現出較為明顯的互為消長的動態關系。昆蟲生態學相關理論顯示,捕食者與被捕食者之間的關系符合Lotka-Voterra方程,也即,隨著捕食者種群的增長,被食者種群逐步下降,當被食者種群降至某一定低值時,捕食者種群的數量因饑餓而下降,使被食者種群得以恢復,至當被食者種群密度升至某個較高密度時,捕食者種群的數量又得以上升,如此往復,循環不息[14]。顯然,本文中的捕食性天敵與樹冠葉蟬屬于捕食者與被捕食的關系,加之清耕地樹冠上的葉蟬數量又總體高于生草地,因此,樹冠上的捕食性天敵對樹冠有害葉蟬具有控制作用。
4 結論
綜上所述,桃園混種牧草后,首先在樹下吸引了對桃樹無害的植食性昆蟲條沙葉蟬和苜蓿斑蚜,其次又吸引了捕食性天敵昆蟲,這些捕食性天敵昆蟲又有一部分遷徙到了樹冠上而使樹冠上捕食性天敵增多,最終對樹冠上的害蟲起到了更好的控制作用。
猜你喜歡
中國化肥信息(2022年8期)2022-12-05 20:01:27
今日農業(2022年13期)2022-11-10 01:05:49
浙江人大(2022年4期)2022-04-28 21:37:09
北京測繪(2021年12期)2022-01-22 03:33:36
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年14期)2021-10-14 08:35:16
當代陜西(2019年10期)2019-06-03 10:12:12
小學生必讀(低年級版)(2018年9期)2018-12-13 01:17:06
浙江國土資源(2015年5期)2015-05-09 08:19:30
中國農資(2014年13期)2014-02-06 16:06:06
