草-溞生物組合修復農田周邊富營養化水體氮、磷研究
2021-02-22 08:46:38張春雪王晗陽魏孝承鄭向群
湖北農業科學 2021年1期
張春雪,王晗陽 ,2,魏孝承 ,鄭向群 ,楊 波
(1.農業農村部環境保護科研監測所,天津 300191;2.天津城建大學,天津 300384)
人類的生產和生活加速了氮、磷等營養鹽向水環境的排放,造成藻類、浮游生物數量的快速增加,使水體的透明度、溶解氧等大幅度下降。一些藻類代謝產物使得水質迅速惡化,進而造成魚類、大型水生植物等生物大量死亡,最終導致水環境生態平衡被嚴重破壞[1-3],尤其是池塘等封閉水體,由于水體長時間處于停滯狀態,缺少交換過程,因而其自凈能力較弱,一旦受到外源氮、磷的輸入,就會加重其富營養化情況[4]。池塘富營養化會導致藻類生長過剩,從而造成藻類水華,對其他水生生物包括對大型水生植物和水生動物的生長、繁殖能力等產生抑制作用[5],最終引起水生生物的大量死亡[6]。
據2018年度《中國水資源公報》[3]顯示,目前中國大多數水體環境面臨著嚴重的富營養化問題。在所調研的121個湖泊和1 097個水庫中,有73.5%的湖泊和30.4%的水庫目前都處于不同程度的富營養化狀態。富營養化導致水華現象頻發,近年來太湖、巢湖、滇池[4]以及天津市的于橋水庫[5]均發生過不同程度的藍藻水華。藍藻水華會引起水生態系統結構和功能退化,致使中國水質性缺水日益嚴重,危害人體健康和生態安全,且對當地經濟造成巨大的損失[6]。因此,對藍藻水華的抑制和富營養化水體的修復展開研究具有重要的現實意義。
關于水體富營養化和水華的治理方法通常分為外源控制法和內源控制法兩大類[7]。外源控制法主要指通過控制農業面源污染,減少工業廢水的排放等[8],從水體外源減少氮、磷等營養鹽的輸入,從而實現控制水體富營養化的方法。但由于中國農業面源污染來源分散、復雜,溯源困難,并且不同于點源污染,其排放通常具有隨機性[9],因而對其管理和控制產生難度。此外,一些水體流域的污水管網建設落后,導致外源控制的效果不穩定。內源控制指通過各類措施從水體內部著手,降低水體中氮、磷等營養鹽的濃度,從而抑制浮游植物過量生長[10]。目前常見的內源控制方法有物理法、化學法以及生物法。生物法的原理是通過生物之間的補食作用及種間競爭等方式達到抑制藻類生長、恢復水生生態系統穩定性、提高水體自凈能力的方法[11]。目前常見的生物法主要包括微生物修復法、生物操縱法、水生植物修復法以及聯合生物修復法。生物修復作為一種新興的環境修復方法,與傳統物理化學法相比,具有低成本、環境友好、二次污染小等優點。生物修復已成為當前環境修復的研究熱點,近年來被廣泛應用于土壤、地表水及地下水等環境的污染治理中。
近年來,一些學者開始考慮將生物操縱與水生植物這2種理論聯合,研究其對水華抑制和富營養化水體的修復作用,在生態修復實踐中,采用2種或2種以上水生生物聯合修復富營養化水體的技術受到廣泛關注。李萍等[12]以背角無齒蚌(Anodonta woodiana,俗稱河蚌)和苦草(Vallisneria natans)為研究對象,構建中型生態系統,采用二者單一及聯合修復富營養化湖泊水體,表明兩者的協同作用可以明顯降低水體葉綠素及營養鹽濃度,此外還發現背角無齒蚌可以促進苦草的生長,試驗結束時二者共存的系統中苦草的株高比單培養時增高24.9%。郭穎[13]分別采用濾食性底棲動物河蚌、田螺與沉水植物菹草(Potamogeton crispus)構建復合表面流人工濕地,發現其對氮、磷等營養鹽去除效果顯著,尤其是河蚌-菹草系統表現出了更好的污染物去除效果,對總氮(TN)、總磷(TP)的去除效果分別達82.80%和98.54%,顯著高于其他系統,但河蚌在濾水過程中會產生“假糞”,其中包含大量氨、尿素、尿酸、可溶性磷等物質,且氨氮(NH4+-N)占總排泄量的60%~70%,因此在河蚌-菹草濕地系統中,河蚌是通過擾動、強化基質中微生物的作用來去除水中NH4+-N,其本身沒有起到凈化NH4+-N的作用。霍元子等[14]向上海市滴水湖修復區投放大型溞,利用大型溞濾除水體懸浮物和藻類,提高水體透明度,之后再移栽伊樂藻、輪葉黑藻等沉水植物,通過長期監測發現,水體中TN、NH4+-N和TP等水質指標的濃度均顯著低于未實施修復的對照區,水體長期維持在國家Ⅱ到Ⅲ類地表水標準內。馬進等[15]將沉水植物苦草和大型溞聯合使用,建立草-溞處理系統修復富營養化池塘水體,結果表明,經過草-溞處理系統處理后的水中氮、磷營養鹽大幅度下降,水體透明度顯著提升;除此之外,經過41 d的處理之后,草-溞聯合處理中苦草濕重增長率(740%)顯著高于單一使用苦草時的增長率(470%),說明大型溞對苦草的生長有促進作用。
現有研究中對生物操縱已有一定的成果,但對生物操縱是否能夠有效抑制產毒藍藻,濾食性動物在修復水體時的添加密度是否會影響其對水華的抑制作用,水生植物修復和生物操縱能否完全相輔相成,此類生物修復過程中對水體浮游細菌群落結構會造成何等影響等問題,仍需進一步討論和研究。
1 材料與方法
1.1 試驗水體來源
原水取自天津市寧河區某農田周邊的富營養化池塘,其水質參數:TN為(7.03±0.35)mg/L,TP為(0.28±0.03)mg/L,葉綠素 a含量為(272±13)μg/L,經計算,綜合營養狀態指數為79.81,參考富營養化評級標準[15],判斷其為重度富營養化水質。
將上述水體用13號浮游動物網濾除水中原有的大型浮游動物,并用滅菌后的BG-11(4%)培養液按1∶1稀釋。BG-11培養液組成見表1。稀釋后,其TN、TP濃度與原水的TN、TP濃度一致;而原生浮游植物和細菌群落稀釋為原水含量的一半,為浮游植物和細菌群落保留一定的生存空間,以此作為本試驗的供試水體,考察試驗設置對富營養化池塘中原生浮游植物和細菌群落的影響。試驗水體初始水質為pH 8.77、溶解氧(DO)9.28 mg/L、透明度17.20 cm、TN 7.38 mg/L、TP 0.31 mg/L、NH4+-N 0.21 mg/L、硝氮(NO3--N)6.59 mg/L、葉綠素 a 68.59 μg/L、浮游植物密度2.80×107cells/L、藍藻門密度2.52×107cells/L、微囊藻毒素2.77 μg/L。
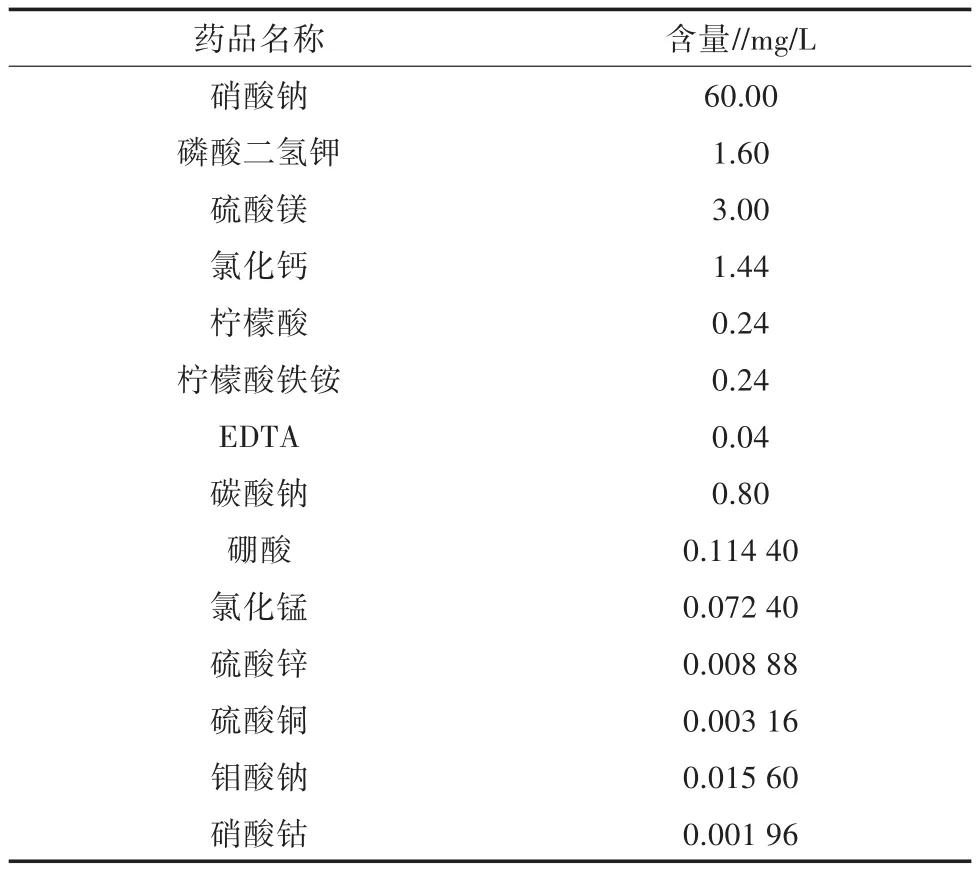
表1 BG-11(4%)培養液組成
1.2 供試動植物
本著高攝食量、低排泄量且不會造成生物入侵的原則,選用大型溞(Daphnia magna)作為本試驗的供試動物。大型溞溞種購自中國科學院沈陽應用生態研究所,試驗前在試驗室內擴大培養,將其置于盛有供試水體的1 L錐形瓶中,在人工氣候箱內馴化培養60 d以上,馴化溫度為25℃,光照度為1 200 lx,光暗比12 h∶12 h,每日喂食20 mL斜生柵藻藻液,隔日換水。試驗前通過孔徑2 mm的水溞濾網過濾選取個體尺寸一致、活潑健康的子代,在純水中清養24 h,使其盡量排空腸道后用于試驗。
綜合考慮水生植物的經濟性和環境友好性,供試植物選擇粉綠狐尾藻(Myriophyllum aquaticum)和金魚藻(Ceratophyllum demersum)作為本試驗的供試植物。粉綠狐尾藻和金魚藻購于天津市某花鳥魚蟲市場,試驗前在4%的BG-11培養液中馴養15 d,使其適應試驗環境,再選取生長良好且大小相似的植株用于試驗。
1.3 試驗裝置
試驗裝置(下稱試驗桶)為33個PVC圓桶,容積為20 L。試驗前使用75%乙醇滅菌,并用去離子水反復沖洗(圖1)。

圖1 試驗裝置
1.4 試驗設計
采用日光燈模擬春、夏時期的日光光照,光照度為(1 200±100)lx,光暗比為12 h∶12 h。將供試水體充分混勻后分裝至各個試驗桶內,初始水位均為20 L,剩余水體用于各指標檢測,以此作為試驗0 d時的初始值。在每個試驗桶內設置相同的恒溫加熱棒,溫度均設置在25℃,并用溫度計監測水溫,以確保試驗水溫恒定,試驗時間為20 d。試驗設置均基于預試驗的結果優選而得,共計9個處理:CK,不投加動植物對照;處理A、B分別投加高、低2種不同密度的大型溞;處理C、D分別投加相同濕重(60 g)的狐尾藻、金魚藻;處理E、F、G、H為高、低密度大型溞和與2種植物兩兩組合即草-溞聯合,每組均設有3個平行試驗桶,具體設置見表2。
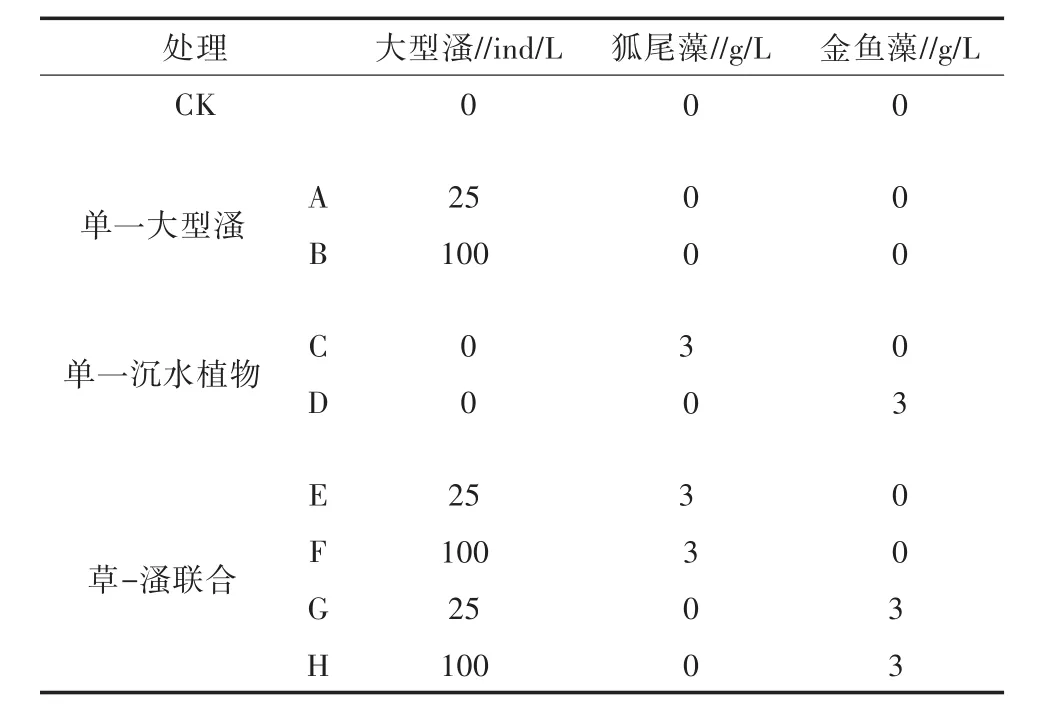
表2 試驗設置
1.5 樣品的采集與測定
1.5.1 樣品的采集 在試驗2、5、10、15、20 d時,測定水體pH、DO、TN、TP、NH4+-N、NO3--N。采樣時間為上午8:00—9:00,為減少試驗誤差,在每次取樣后記錄水位,在下次取樣前采用蒸餾水補足蒸發量[16-18]。
1.5.2 樣品的測定 ①pH、DO。采用雙輸入多參數數字分析儀(HACH HQ40d,USA)對試驗水體原位測定,在各平行試驗桶底部0~20 cm、20~40 cm處分別測定后取平均值。②TN、TP、NH4+-N、NO3--N濃度。取水樣20 mL,營養鹽指標采用流動分析儀測定。TN濃度采用堿性過硫酸鉀消解-紫外分光光度法測定;TP濃度采用鉬銻抗分光光度法測定;NH4+-N濃度采用納氏試劑光度法測定;NO3--N濃度采用酚二磺酸光度法測定。③大型溞密度。使用玻璃棒攪拌混勻水體,取1 L水樣采用3%的甲醛溶液固定并濃縮至50 mL,采用1 mL計數框于顯微鏡100×下定量計數,每個樣品均計數3次后取平均值。④植物濕重。將各試驗桶中的水生植物取出,用濾紙吸干表面的水分后,采用電子天平稱重,每個樣品稱3次取平均值。
理化指標的消減率計算公式為:
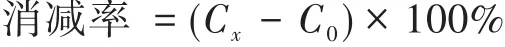
式中,C0為某指標的初始濃度,Cx為該指標在第x天的濃度。
1.6 數據統計分析
各項指標采用Origin 9.0制圖,數據均以3個平行試驗桶的平均值±標準差形式表示。采用SPSS 24.0進行統計分析,通過單因素方差分析(one-way/ANOVA)檢驗指標的差異顯著性,圖基檢驗(HSDtest)用于單因素方差分析后的兩兩比較,顯著水平為0.05。
2 結果與分析
2.1 對水體pH、DO的影響
2.1.1 對pH的影響 試驗期間各處理水體的pH變化如圖2所示。由圖2可知,不同處理對水體pH的影響不明顯,pH均在8.2~9.3波動,各組pH差異均不顯著(P>0.05),整體處于偏堿性狀態。研究表明,水體處在偏堿性環境中有利于微生物的硝化作用、NH4+-N的揮發和TP的去除,但pH過高會影響植物對營養鹽的吸收作用[19-21]。大型溞的呼吸作用會向水中釋放CO2降低水體pH,但沉水植物的光合作用可向水中釋放O2,緩解了大型溞對pH的降低作用。
2.1.2 對DO的影響 試驗期間水體DO濃度變化如圖3所示。由圖3可知,DO濃度隨時間變化存在組間差異(P<0.05)。與CK相比,處理A、B中的DO濃度隨時間變化波動較大。試驗期間,CK的DO濃度波動不明顯,維持在8.4~9.9 mg/L;處理A、B在0~5 d時均發生較大幅度的下降,5 d時分別降至7.21、7.19 mg/L;10~15 d時,處理A、B的DO濃度變化不穩定,15 d時分別降至6.34、5.47 mg/L;試驗結束(20d)時,處理A、B的DO濃度有所回升,分別升至6.52、5.89 mg/L,但仍顯著低于CK(P<0.05)。處理C、D與CK之間存在顯著差異(P<0.05),試驗結束時,表現為D(5.72 mg/L)<CK(9.09 mg/L)<C(10.03 mg/L),兩兩比較(HSDtest)表明,處理C的DO濃度顯著高于處理D(P<0.05),這是由于狐尾藻較金魚藻具有龐大的根系系統[22],其強大的釋氧作用可使水中DO濃度顯著提升。處理G、H的DO濃度顯著低于處理E、F和CK(P<0.05);處理E、F與CK則無顯著差異(P>0.05)。推其原因,一方面是對照浮游植物大量繁殖,采樣時均在光照期,浮游植物通過光合作用產生了大量氧氣;另一方面是大型溞的呼吸耗氧量高于金魚藻的釋氧;在試驗10 d之后,處理G、H的DO濃度開始回升,試驗結束時分別恢復至7.63、7.12mg/L。這是由于后期金魚藻生長加快,葉莖組織逐漸龐大,此時金魚藻的釋氧高于大型溞的耗氧所致。

圖2 不同處理對水體pH的影響
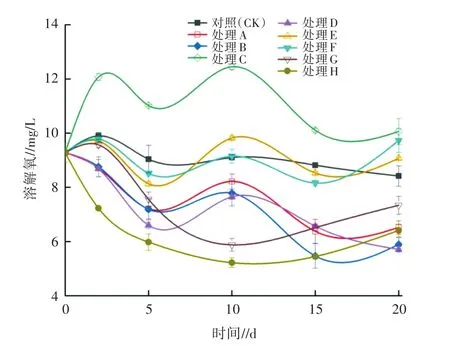
圖3 不同處理對水體DO的影響
2.2 對水體氮素營養鹽的影響
2.2.1 對TN的影響 對試驗期間各處理水體中TN濃度進行監測,其變化規律如圖4所示。由圖4可知,各處理水體中TN濃度隨處理時間延長均呈下降趨勢,且均在20 d時降至最低。處理A、B水體中TN濃度與CK均有所差異。試驗0~10 d時,處理A、B中的TN濃度均高于CK,10 d之后,TN濃度迅速下降。試驗結束時,各處理和對照的TN濃度由低到高依次為 C(1.65 mg/L)、E(1.88 mg/L)、F(2.35 mg/L)、D(3.18 mg/L)、G(3.59 mg/L)、H(3.86 mg/L)、A(4.57mg/L)、B(4.80 mg/L)、CK(5.22 mg/L),此時對應的TN消減率為CK(29.27%)<B(34.96%)<A(38.08%)<H(47.69%)<G(51.36%)<D(56.91%)<F(68.16%)<E(74.53%)<C(77.64%),且處理C、E、F對TN的消減率均顯著高于D、G、H、A、B、CK。
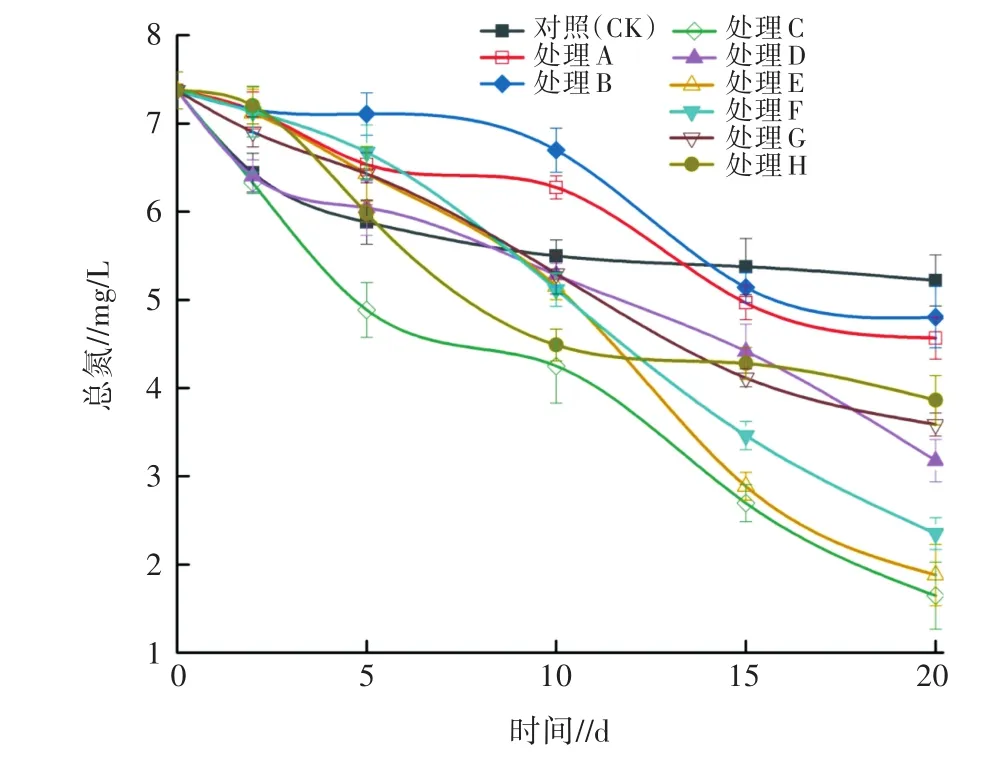
圖4 不同處理對水體TN的影響
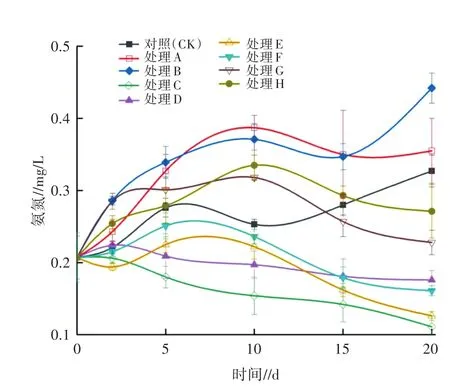
圖5 不同處理對水體NH4+-N的影響
2.2.2 對NH4+-N的影響 對試驗期間各處理水體中NH4+-N的濃度進行監測,其變化規律如圖5所示。由圖5可知,試驗期間,各處理NH4+-N濃度呈現不同的趨勢,C、D、E、F在20 d時有所下降,其余各處理則呈上升趨勢。試驗結束時,各處理水體中的NH4+-N濃度由低到高依次為C(0.11 mg/L)、E(0.13 mg/L)、F(0.18 mg/L)、D(0.19 mg/L)、G(0.24 mg/L)、H(0.28 mg/L)、CK(0.34 mg/L)、A(0.36 mg/L)、B(0.44 mg/L),僅處理A和B中的NH4+-N濃度高于CK,并達到統計學上的顯著水平(P<0.05)。處理C和E對NH4+-N的消減作用最明顯,且處理E(38.10%)較處理F(14.29%)消減作用更為顯著(P<0.05)。
2.2.3 對NO3--N的影響 對試驗期間各處理水體中NO3--N的濃度進行監測,其變化規律如圖6所示。由圖6可知,各處理NO3--N濃度隨處理時間呈下降趨勢,試驗結束時,NO3--N濃度由低到高依次為 E(1.62 mg/L)、F(1.89 mg/L)、C(1.91 mg/L)、D(2.69 mg/L)、G(3.76 mg/L)、H(3.84 mg/L)、A(3.98mg/L)、B(4.08 mg/L)、CK(4.42 mg/L),所對應的NO3--N消減率由高到低依次為E(75.42%)、F(71.32%)、C(71.02%)、D(59.18%)、G(42.94%)、H(41.73%)、A(39.61%)、B(38.09%)、CK(32.93%);各處理對NO3--N的消減率均顯著高于對照(P<0.05);兩兩比較(HSDtest)顯示,處理E、F對NO3--N的消減率均顯著高于G、H。由“2.2.1”可知,處理E、F對TN的消減率也均顯著高于G、H,說明不同密度大型溞與狐尾藻的聯合(處理E、F)對TN和NO3--N的消減作用較其與金魚藻聯合處理時更優。

圖6 不同處理對水體NO3--N的影響
2.3 對水體磷素營養鹽的影響
對試驗期間各處理水體中TP濃度進行監測,其變化規律如圖7所示。由圖7可知,試驗期間,處理G、H的TP濃度總體呈先上升后下降的趨勢;處理E、F的TP濃度呈持續下降趨勢。各處理TP濃度均在試驗結束時降至最低,此時,各處理和對照之間的TP濃度從低到高依次為C(0.06 mg/L)、E(0.09 mg/L)、D(0.11 mg/L)、F(0.12 mg/L)、G(0.14 mg/L)、A(0.15 mg/L)、H(0.16 mg/L)、B(0.18 mg/L)、CK(0.20 mg/L),此時各處理對應的TP消減率由高到低依次為C(80.65%)、E(70.97%)、D(64.52%)、F(61.29%)、G(54.84%)、A(51.61%)、H(48.39%)、B(41.94%)、CK(35.48%)。其中,處理E、F的TP消減率顯著高于CK(P<0.05),處理 G、H 則與 CK 差異不顯著(P>0.05)。結果表明,大型溞和狐尾藻聯合處理對TP的消減作用明顯高于大型溞和金魚藻聯合處理。處理C、D對TP消減率顯著高于CK,且處理C對TP的消減率顯著高于處理D(P<0.05)。
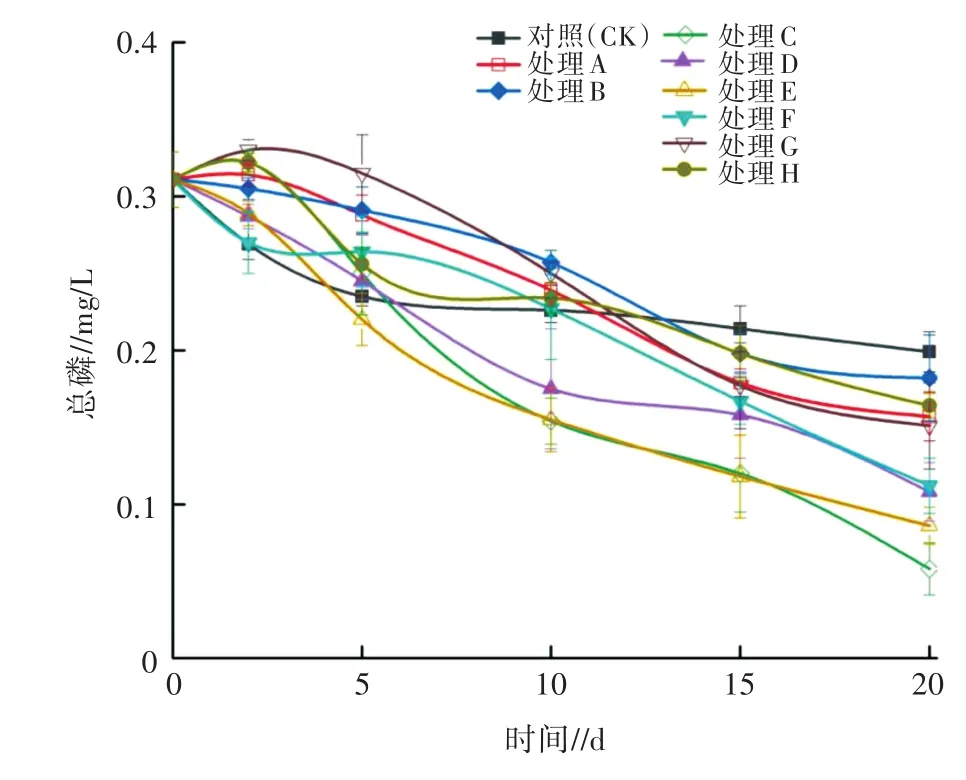
圖7 不同處理對水體TP的影響
3 討論
大型溞對水體pH的影響較小;受呼吸作用影響,水體DO濃度隨溞濃度增大而降低。狐尾藻和金魚藻對水體pH影響較小。試驗期間,金魚藻的DO濃度低于對照;而狐尾藻由于其強大的根系釋氧作用可以顯著提升水體的DO濃度。草-溞聯合處理對水體pH的影響較小。相比于對照,大型溞+狐尾藻對水體DO濃度的影響較弱。試驗0~10 d時,由于金魚藻釋氧作用低于大型溞的呼吸作用,致使大型溞+金魚藻處理水體DO濃度顯著降低;10 d后金魚藻葉莖壯大,DO濃度回升。
大型溞可以通過攝食水中的浮游植物、顆粒物來同化營養鹽,因而對TN、TP、NO3--N等營養鹽均有一定的消減作用。與此同時,對照中浮游植物大量繁殖也對N、P營養鹽有一定消耗。因此,大型溞的添加對營養鹽的消減與對照相差不顯著。此外,大型溞作為動物,其機體對N、P營養鹽的需求有限,未被吸收的N、P等物質將會以糞便的形式排出體外,從而引起水環境中N、P營養鹽的回升。由于高密度大型溞排泄作用較大,在同等試驗條件下,導致其N、P營養鹽的濃度較低密度處理略高。研究表明,大型溞暴露在含有一定濃度產毒藍藻的水環境中,其生長周期會縮短至 13~15 d[23],低于試驗周期所設定的時間(20 d)。此外,由于受到種群競爭因素的影響,試驗過程中,大型溞群體均有死亡現象,死亡個體向水中釋放NH4+-N,從而造成試驗過程中NH4+-N濃度的升高。
沉水植物在生長過程中可吸收大量的營養鹽,因此本試驗中狐尾藻和金魚藻對TN、TP、NO3--N、NH4+-N等的消減作用均較為明顯。狐尾藻具有增長較快、對營養鹽富集量大的特點,此外,還具有冠層優勢,可以浮在水面充分利用光照條件快速生長,金魚藻則是整個浸沒在水體中,因而生長速度較慢,試驗結束時狐尾藻組的平均濕重為(72.84±5.87)g,高于金魚藻組的(63.52±9.43)g。狐尾藻根系龐大,輸氧作用強,可以加強根系微生物的硝化-反硝化作用,而金魚藻屬于無固定根類型的植物,只能靠莖葉吸附水體營養鹽,因此本試驗結果中狐尾藻對各項營養鹽的消減作用均高于金魚藻。
相比于單一使用溞處理時對N、P營養鹽無顯著消減作用的現象,聯合處理可明顯降低營養鹽的濃度。草-溞聯合對水體營養鹽的消減作用具有協同性,大型溞+狐尾藻對營養鹽的消減作用均優于大型溞+金魚藻,說明在營養鹽的消減上,沉水植物起到主要作用。本試驗結果中,低密度(25 ind/L)大型溞+狐尾藻對于TN、NO3--N、TP的消減作用與高密度(100 ind/L)大型溞+狐尾藻差別較小,說明大型溞對于營養鹽的攝食和同化作用有限,試驗中大型溞的排泄作用和死亡個體向水中釋放了NH4+-N,但沉水植物的對營養鹽的吸收作用,改善了NH4+-N上升的現象。研究表明,狐尾藻有親氨性,對水中NH4+-N有較強的吸收作用[24],這解釋了處理E、F的NH4+-N濃度在試驗后期下降的原因。
4 小結
近年來,水體富營養化現象日趨嚴重,已成為亟需解決的環境問題之一。本研究基于生物操縱理論和水生植物修復理論,結合目標水體現狀,采用大型溞作為濾食性動物的代表,狐尾藻和金魚藻作為沉水植物的代表,在實驗室可控條件下,模擬春、夏水華易發生時期的光照和溫度條件,考察單一生物及其組合對富營養化水體的修復效果,為生物操縱和水生植物修復富營養化水體的應用提供數據支持。試驗結果表明,大型溞、狐尾藻、金魚藻以及草-溞聯合對水體pH均無顯著影響;各處理均對水體DO有一定的改善效果;草-溞聯合處理相比于單一溞處理可有效提升水中的DO濃度,其中大型溞+狐尾藻效果更優。大型溞對水體TN、TP和NO3--N消減作用不顯著,投加密度對營養鹽的消減作用影響較弱;NH4+-N濃度隨大型溞投加密度的增高而上升;狐尾藻和金魚藻對TN、TP、NO3--N、NH4+-N的消減作用顯著,其中狐尾藻消減作用更優。草-溞聯合處理均可顯著消減水體中的TN、TP、NO3--N濃度,且明顯優于單一溞處理時的抑制能力,其中以低密度大型溞+狐尾藻的消減作用最為顯著。金魚藻和狐尾藻均能夠抑制因大型溞排泄和死亡致使的NH4+-N濃度上升,且狐尾藻的抑制作用更強。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
科學大眾(2021年9期)2021-07-16 07:02:54
當代陜西(2021年2期)2021-03-29 07:41:24
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
