酪丁酸梭菌發酵產丁酸研究進展
2021-02-25 09:05:14衡文葉光斌鄒偉
中國調味品 2021年2期
衡文,葉光斌,鄒偉
(四川輕化工大學 生物工程學院,四川 宜賓 644005)
丁酸(butyric acid)又名酪酸,是含4個碳原子的飽和一元羧酸,具有揮發性。丁酸應用十分廣泛:在化工領域可作為原料用于熱塑性材料合成[1];在食品領域可作為風味成分增強食物風味[2-4];在飼料行業可作為新型飼料添加劑,提高牲畜的免疫力水平和營養價值,減少抗生素的使用[5];在醫藥領域可作為輔助用藥對腸道具有修護作用[6-7]。2016年全球丁酸年產量已超過8×104t[8],且呈逐年上升的態勢。目前,丁酸主要的生產方法可分為化學合成法和微生物發酵法。其中,微生物發酵法相較于化學合成法在環境保護、資源再利用等方面更具優勢,更符合可持續發展戰略,受到國內外學者普遍關注。厭氧條件下,丁酸是酪丁酸梭菌(Clostridiumtyrobutyricum)、丁酸梭菌(C.butyricum)、楊木梭菌(C.populeti)等菌株主要代謝產物[9-11]。其中,C.tyrobutyricum因丁酸產量大、產率高、純度高、穩定性好等優勢,被認為是發酵生產丁酸最具商業化潛力的菌株[12-13]。
1 C. tyrobutyricum基本生物學特征
酪丁酸梭菌(Clostridiumtyrobutyricum)隸屬于芽孢桿菌科(Bacillaceae)、梭菌屬(Clostridium),是具有丁酸生產能力的有機化能異養型專性厭氧菌。菌體中間隆起,其直徑約為0.3~2.0 μm×1.5~2.0 μm,端圓,單個或成對出現,菌體呈短鏈,偶見長絲狀菌體,周身鞭毛,能運動,革蘭氏染色呈陽性。平板上菌落表面呈圓形,稍突,邊緣完整,呈煎蛋狀,灰色半透明,表面有光澤,直徑約在1~3 mm。由于菌體中常有圓形或卵圓形的巨大芽孢,對高溫、高鹽、強酸等惡劣環境的耐受性強,在pH 4.5~7.0,35~37 ℃的條件下均能很好地生長[14]。C.tyrobutyricum因發酵乳酸破壞奶酪制品風味而為人所熟知,常分離于動物消化系統、土壤、廢水、青貯飼料、乳制品污染環境樣品中。
2 C. tyrobutyricum丁酸發酵途徑解析
C.tyrobutyricum的主要代謝產物為丁酸和丁醇,常伴有副產物乙酸,也能產生少量乳酸和乙醇[15]。結合文獻和數據庫信息重構C.tyrobutyricum利用不同底物(葡萄糖、木糖、甘油、甘露糖)合成丁酸及其相關副產物的代謝途徑,見圖1。
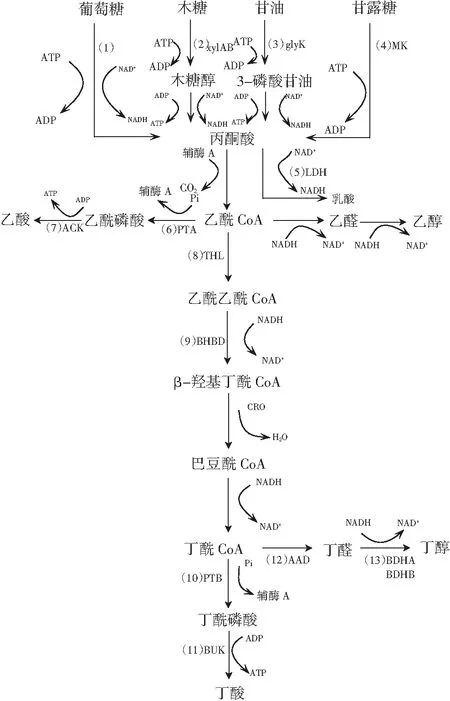
圖1 C. tyrobutyricum發酵產丁酸代謝途徑Fig.1 The metabolic pathway of producing butyric acid by C. tyrobutyricum注:(1)EMP途徑;(2)xylAB:木糖異構酶、木糖激酶;(3)glyK:甘油激酶;(4)MK:甘露糖激酶;(5)LDH:乳酸脫氫酶;(6)PTA:磷酸甘油酯酶;(7)乙酸激酶;(8)THL:硫解酶;(9)BHBD:β-羥丁基輔酶A;(10)PTB:磷酸轉乙酰輔酶;(11)BUK:丁酰激酶;(12)AAD:醇/醛脫氫酶;(13)BDHA/BDHB:丁醇脫氫酶。
以葡萄糖作為底物時[16],當葡萄糖進入代謝途徑,通過EMP途徑將葡萄糖分解生成丙酮酸、NADH和ATP。生成的丙酮酸一部分在乳酸脫氫酶的作用下生成乳酸、NADH;另一部分丙酮酸在丙酮酸脫氫酶的作用下生成中間產物乙酰CoA。乙酰CoA又分別參與乙酸、乙醇及丁酸代謝通路:在乙酸代謝通路中,乙酰CoA在磷酸轉乙酰酶的作用下生成乙酰磷酸,再進一步在乙酸激酶的催化下生成乙酸;在乙醇支路中,乙酰CoA在丙酮酸脫氫酶催化下生成乙醛,再在醇脫氫酶的作用下生成乙醇。丁酸合成途徑首先乙酰CoA在硫解酶的作用下生成乙酰乙酰CoA,再在β-羥基丁酰CoA脫氫酶的作用下繼續生成β-氫羥基丁酰CoA,此時巴豆酸酶發揮作用將β-羥基丁酰CoA轉化為巴豆酰CoA,后在丁酰CoA脫氫酶的作用下進一步生成關鍵前體物質丁酰CoA,前體物質在磷酸丁酰轉移酶、丁酰激酶的催化下最終形成丁酸。以木糖作為底物時[17],木糖底物經戊糖-磷酸途徑分解為果糖-6-磷酸和甘油醛-3-磷酸,然后經過一系列酶的磷酸化、外膜化形成丙酮酸木糖[18]。以甘油作為底物時[19],甘油在甘油激酶的催化下生成α-甘磷酸甘油,再通過脫氫變為磷酸二羥丙酮,再經糖降解轉化為丙酮酸。以甘露糖為底物時[20],在甘露糖激酶的作用下生成6-P-甘露糖,再經異構酶、激酶催化生成丙酮酸。不論是木糖、甘油還是甘露糖,都是通過一系列反應生成中間產物丙酮酸,后續代謝途徑與葡萄糖代謝途徑一致,故不贅述。
3 C. tyrobutyricum利用生物質原料發酵產丁酸
化學合成帶來的環境污染以及不可再生的自然資源消耗殆盡等問題不斷凸顯,迫使人們不得不尋找綠色經濟可再生的原料代替化石燃料生產丁酸。生物質資源作為地球上儲量最豐富的可再生性資源,全球每年通過光合作用生成的生物質資源可以達到 2000億噸,所蘊含的能量是全球能量需求的數十倍[21]。其中以木質纖維素形式儲存的能量能夠占到 60%以上,是地球上最為豐富的有機資源[22],是代替化石燃料生產丁酸的絕佳原料。
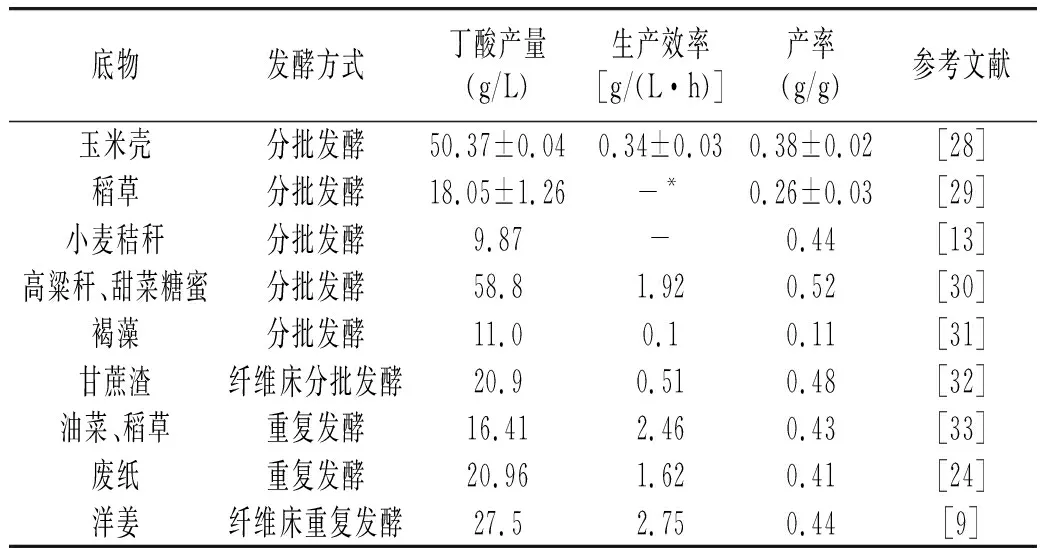
表1 C. tyrobutyricum利用不同生物質發酵產丁酸研究Table 1 Study on production of butyric acid by fermentation of C. tyrobutyricum with different biomass
由表1可知,C.tyrobutyricum在不同發酵方式下對生物質原料均具備較好的利用能力。自由細胞發酵時,由于傳質能力的遞減及產物濃度的反饋抑制作用,C.tyrobutyricum發酵生產丁酸的能力未得到充分展現。利用固定化細胞進行發酵生產能夠一定程度上解決生產能力低下的問題[23-24]。生物質原料雖然來源廣、成本低,但其復雜的細胞結構增加了生物轉化的難度,水解液中酚類化合物對C.tyrobutyricum會造成毒害,這些問題在不同程度影響了C.tyrobutyricum產丁酸的發展。研究發現,利用稀硫酸[25]、γ-戊內酯/1-甲基咪唑二甲基亞磷酸復合體系[26]進行預處理能夠一定程度上破壞生物質復雜結構,目的產物產量分別增加了42.62%、46.8%。另外,采用混合生物質發酵,能一定程度上降低水解液的毒害作用從而提高丁酸產量[27]。這些方法不僅對于C.tyrobutyricum發酵生物質有效,同時也為其他具有生物質發酵能力菌株的研究提供了參考思路。
4 遺傳工程改造C. tyrobutyricum發酵產丁酸
目前,C.tyrobutyricum發酵產丁酸主要受三類因素限制:底物攝取率低;菌株對丁酸、酚類物質耐受程度低;丁酸代謝途徑復雜,代謝流分支較多。研究顯示:依靠遺傳工程技術,對菌株進行定向改造能夠有效打破這些限制,提高丁酸產量。
4.1 提高底物吸收效率
以葡萄糖介導的碳分解代謝物抑制效應(carbon catabolite repression, CCR)是造成C.tyrobutyricum木糖分解率低下的主要原因。過量表達木糖質子——同工酶(xylT)、木糖異構酶(xylA)和木糖激酶(xylB)3個基因,通過分批發酵,產物產量由3.2 g/L提升至12.0 g/L,能有效打破CCR的限制。若在發酵液中加入紫羅堿(viologen BV),人工電子載體加快電子的傳遞,Ct-pTBA的突變體經自由細胞分批發酵,最終產量可達46.4 g/L,產率為0.43 g/g,產率為0.87 g/(L·h)。實驗還發現,該突變體在各種未脫毒木質纖維素生物質水解物中具備優異的葡萄糖和木糖協同利用能力,通過自由細胞分批發酵丁酸終產量達29.7~42.6 g/L的較高濃度[34]。此外,克隆α-隆葡糖苷酶中的agluⅠ和agluⅡ導入C.tyrobutyricum細胞內,可使其獲得分解麥芽糖、淀粉的能力[35]。克隆蔗糖代謝途徑的scrA,scrB,scrK基因,可使C.tyrobutyricum獲得蔗糖分解的能力[36]。此類研究拓展了C.tyrobutyricum可利用底物的范圍。
4.2 提高菌株耐受性
C.tyrobutyricum對環境中丁酸及酚類物質濃度的耐受能力與Ⅰ類熱休克蛋白基因相關性較高,過表達groESL基因能有效提高C.tyrobutyricum對丁酸的耐受能力,而過表達dnaK則會降低其耐受能力。利用過表達groESL基因的菌株進行固定化細胞補料分批發酵丁酸最終產量可達52.2 g/L,較之前提高了15.2%[37]。天然基因groESL的過表達還能提高菌株對水解物中酚類物質的耐受性,丁酸產量較野生型增加了26.5%,達到29.6 g/L[38]。除Ⅰ類熱休克蛋白外,過表達短鏈脫氫酶/還原酶(short-chain dehydrogenase/reductase,SDR)也能增強菌株對呋喃衍生物和酚類化合物的耐受性,當SDR與groESL基因共表達時,丁酸產量還能得到進一步提升[39]。
4.3 調控丁酸代謝流
在丁酸代謝途徑中,磷酸甘油酯酶(phosphoransacetylase,pta)、乙酸激酶(butyrate kinase,ak)、磷酸轉乙酰輔酶(phosphotransbutyrylase,ptb)和丁酸激酶(butyrate kinase,bk)基因起著重要的作用。其中,pta和ak酶將乙酰CoA轉化為乙酸,ptb和bk酶催化丁基CoA生成丁酸。從代謝途徑進行分析,抑制乙酸支路代謝的進行,能促進代謝流更多流向丁酸發酵途徑,見圖1。
Zhu Y等[40]通過滅活pta基因將丁酸濃度提高了60.8%(野生型:20.2 g/L,突變型:32.5 g/L),pta、ak活性分別降低了60%、80%;進一步采用基因敲除技術敲除ak基因,在新型纖維床生物反應器(fibrous bed bioreactor,FBB)體系中,細胞固定化發酵的最終丁酸濃度達到50.1 g/L較高水平[41]。Ptb基因在丁酸形成中十分重要,根據代謝通路圖可合理推測敲除ptb基因,丁酸生成將會降低。然而,Zhang Y等[42]研究發現敲除C.tyrobutyricum中的ptb基因,丁酸產量并無明顯變化。這可能是細胞中其他同工酶代替磷酸轉乙酰酶發揮了作用。此外,過表達EMP途徑中的6-磷酸果糖激酶(pfkA)和丙酮酸激酶(pykA)也可一定程度上促進丁酸的累積[43]。研究發現,不管以何種方式去除乙酸、丁酸途徑中的關鍵基因,并不能完全阻斷對應代謝產物的生成。這表明必須掌握完整的C.tyrobutyricum代謝網絡內通量分布,進行系統全面的分析。目前已有13株C.tyrobutyricum完成全基因組測序[44]。借助于這些基因組信息可以構建C.tyrobutyricum基因組規模的代謝網絡模型,結合相關約束性算法,可以更理性地指導C.tyrobutyricum代謝工程改造工作[45]。
5 生物反應器改良
C.tyrobutyricum通過自由細胞發酵生產丁酸存在產量低、濃度低、產率低等不足[46]。傳統的生物反應器周期長、設備利用率低、發酵中后期產率低等缺陷也不能滿足工業化產丁酸的目的。纖維床生物反應器(fibrous bed bioreactor,FBB)能夠將細胞固定在纖維基質中,并被廣泛應用各種底物發酵生產丁酸,生產效率和產率得到顯著提高[47]。利用FBB法,C.tyrobutyricum經補料分批發酵,丁酸產量達86.9 g/L,為現有報道最高值[48]。王金等[49]創新性地采用四柱FBB,與單柱發酵相比,四柱FBB提高了發酵效率,丁酸產量提高7.19%,產量提高136.7%,補料分批發酵丁酸最終濃度達到61.4 g/L。Huang J等針對丁酸發酵生產的需要,研制了一種新型的內盤狀基體,該基體由不銹鋼絲組成,織物覆蓋在盤狀基體上,優化各項因素后丁酸產率最優值可達0.47 g/g。Luo H等[50]構建了內纖維床反應器(internal fibrous bed bioreactor,IFBB),通過補料分批發酵,丁酸的產量和生產效率分別達到45.38 g/L、0.630 g/(L·h)。在FBB的生產過程中,降低質量梯度需要通過多次循環泵來實現,不僅增加了總投資,pH值也沒有得到及時的控制。而IFBB克服了FBB的不足,以更簡單的施工程序、更好的傳質混合和更低的價格實現丁酸增產的目的,在工業化應用上具有一定優勢。
6 展望
目前,提高C.tyrobutyricum丁酸產量的途徑主要通過提高底物利用率、增強菌體耐受性以及促進丁酸代謝流3個方面進行。其中,江凌等人經補料分批發酵得到丁酸產量最高值86.9 g/L。盡管國內外對C.tyrobutyricum產丁酸已有較為深入的研究,但其代謝與調控機理仍并未得到全面解析。因此,提高C.tyrobutyricum丁酸產量的主要研究方向將以利用遺傳工程技術,借助生物信息學手段,對遺傳信息進行全面解析為目的,從而實現系統科學地指導丁酸發酵。同時,生物反應器的不斷改進和優化也是實現其規模化、產業化生產的關鍵。隨著未來研究過程中對C.tyrobutyricum遺傳信息的進一步揭示以及蛋白質組學、代謝組學等技術的發展與成熟,C.tyrobutyricum的生理代謝機制將被逐漸明確,其用于工業化發酵生產丁酸也指日可待。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中國果業信息(2019年10期)2019-11-13 01:21:34
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國衛生(2016年3期)2016-11-12 13:23:26
