微小RNA-429過表達對乳腺癌細胞增殖侵襲的影響
2021-02-27 05:58:50喻建牟文輝向英豪王波涌
河北醫藥 2021年1期
喻建 牟文輝 向英豪 王波涌
乳腺癌是女性常見的惡性腫瘤,具有較高的發病率與死亡率[1]。研究顯示,細胞增殖及轉移速度快是乳腺癌發生發展的主要特點[2],因而探究乳腺癌發病機制進而尋求治療新靶點成為研究熱點。研究表明,微小RNA-429(miR-429)在非小細胞癌等多種惡性腫瘤中呈低表達,可參與細胞增殖、凋亡及侵襲等生物學行為[3]。研究表明,Wnt/β-鏈蛋白(β-catenin)經典通路可參與乳腺癌發生及發展過程[4]。研究顯示,miR-429可通過抑制Wnt/β-catenin信號通路進而抑制上皮性卵巢癌細胞增殖及侵襲[5]。然而關于miR-429與乳腺癌發生及發展的研究筆者所見較少。因此,本研究旨在通過miR-429過表達分析其對乳腺癌細胞增殖、侵襲及Wnt信號通路的影響,進一步揭示乳腺癌發生機制提供理論依據。
1 資料與方法
1.1 一般資料 收集2017年8月至2018年6月在利川市人民醫院進行手術治療的76例乳腺癌患者的癌組織標本為乳腺癌組,另選取距癌組織邊緣>5 cm的對應癌旁組織標本為癌旁組。患者經病理證實為Ⅰ~Ⅱ型,手術切除組織迅速置于10%甲醛中。患者年齡28~79歲,平均年齡(52.6±7.6)歲。
1.2 主要試劑與儀器 乳腺癌細胞MCF-7(TCHu74)購自中國科學院典型培養物保藏委員會細胞庫。DMEM細胞培養液與10%胎牛血清(fetal bovine serum,FBS)均購自美國Gibco公司;MTT檢測試劑盒購自美國Sigma公司;LipofectamineTM2000轉染試劑盒購自美國Invitrogen公司;miR-429陰性對照質粒購自上海吉瑪制藥有限公司;Trizol Reagent核酸分離試劑購自日本Takara公司;FastQuant RT Kit(RNA反轉錄試劑盒)與SYBR Green試劑盒均購自北京天根生化科技有限公司;兔抗β-鏈蛋白(β-catenin)、磷酸化糖原合成酶激酶-3β(p-GSK-3)一抗均購自美國Bioworld公司;羊抗兔、羊抗鼠二抗均購自美國ROCKLAND公司;ki67、基質金屬蛋白酶-2(MMP-2)、基質金屬蛋白酶-9(MMP-9)、細胞周期相關蛋白1(CyclinD1)、原癌基因c-Myc抗體均購自美國Abcam公司;Matrigel基質膠購自上海前塵生物科技有限公司;Transwell小室購自美國BD公司;BCA試劑盒、蛋白提取試劑盒均購自碧云天生物技術研究所;ECL化學發光劑試劑盒購自上海翊圣生物科技有限公司。多功能酶標儀購自美國Molecular Devices公司;實時熒光定量PCR儀購自美國ABI公司;D30型核酸蛋白測定儀及凝膠成像儀購自德國Eppendorf公司;TE70X型半干轉膜儀購自上海帝博思生物科技有限公司。
1.3 方法
1.3.1 細胞穩定轉染及轉染效率檢測:取凍存MCF-7細胞,恒溫水浴鍋解凍(37℃)1~2 min,置于含10%胎牛血清及100倍稀釋的青鏈霉素雙抗混合液的DMEM培養基,放入培養箱中培養(5%CO2、37℃),待細胞生長匯合度達到約80%時,用0.25%胰蛋白酶液消化。傳代培養后取對數生長期細胞進行后續實驗。將LipofectaminTM 2000轉染試劑稀釋輕輕混勻,室溫孵育5 min,另將miR-429 mimics、陰性對照分別稀釋于500 μl無血清培養基中使終濃度為20 μmol/L,室溫孵育5 min,將稀釋的脂質體與稀釋的miR-429 mimics混合,輕輕混勻,室溫孵育20 min,分別加入相應培養孔,另設不添加轉染試劑作為空白組。6 h后棄去培養基,更換含10%胎牛血清的RPMI 146培養基。轉染48 h后將培養板置于倒置熒光顯微鏡下觀察,鏡下隨機選擇3個視野,分別計數細胞總數及法綠色熒光的細胞數,計算轉染效率,試驗重復3次,取平均值。當轉染效率80%時,加入嘌呤霉素進行壓力篩選,獲得穩定轉染細胞株。
1.3.2 實時定量聚合酶鏈反應 (quantitative real-time PCR,qRT-PCR)檢測miR-429表達:取出超低溫冰箱內保存乳腺癌組、癌旁組組織以及空白組、miR-429 mimics、miR-429 NC組各組細胞參照Trizol Reagent核酸分離試劑盒說明書提取細胞總RNA并反轉錄為cDNA。qRT-PCR反應條件:95℃ 30 s,95℃ 15 s,60℃ 30 s,共39個循環;95℃ 15 s,60℃ 60 s,95℃ 15 s。miR-429以U6為內參基因,采用2-ΔΔCt法計算miR-429相對表達量。見表1。

表1 qRT-PCR引物序列
1.3.3 MTT法檢測細胞增殖:收集對數期的MCF-7細胞,加入100 μl細胞懸液(密度為1×105個/ml),每組設置4個平行孔,培養24 h后棄原培養液,每孔分別加入20 μl MTT溶液,繼續培養4 h,棄上清,分別加入DMSO溶液(100 μl)震蕩,采用酶標儀分別檢測細胞0、24、48、72、96 h時的吸光度值(570 nm波長),重復3次,取平均值。細胞增殖抑制率(%)=(空白組吸光值-試驗組吸光值)/空白組吸光值×100%。
1.3.4 平板克隆形成實驗:分別取各組處于對數生長期的MCF-7細胞,胰蛋白酶(0.25%)消化后制備單個細胞懸液并接種于6孔板(細胞密度500個/ml),放入5%CO2、37℃培養箱中培養直至出現肉眼可見的克隆細胞,PBS清洗,甲醇(100%)固定15 min,結晶紫液(0.1%)染色15 min,清洗后自然晾干。顯微鏡下觀察細胞克隆數目并計算克隆形成率,克隆形成率(%)=(克隆數/接種細胞數)×100%。
1.3.5 Transwell法檢測細胞侵襲能力:細胞侵襲實驗:Transwell小室上室底膜表面鋪Matrigel膠(40 μl 2.0 mg/ml),室溫孵育30 min,收集各組MCF-7細胞,采用無血清DMEM培養液調整MCF-7細胞濃度(2×105個/ml),首先在Transwell小室的上室加入100 μl細胞懸液,其次在下室加入含10%胎牛血清的DMEM培養液(600 μl),放入培養箱繼續培養24 h,棄上室培養液,采用無水甲醇固定,30 min后擦去上室未透過膜的細胞,結晶紫(0.1%)染色20 min,將其置于倒置顯微鏡下觀察計數。
1.3.6 蛋白免疫印跡法(Western Blot,WB)檢測ki67、MMP-2、MMP-9、細胞核β-catenin、細胞質β-catenin、CyclinD1和c-myc蛋白表達情況:收集各組MCF-7細胞,加入RIPA裂解液提取總蛋白并采用BCA法檢測各蛋白濃度,其中核蛋白提取參照Active Motif公司細胞核提取蛋白說明書。分別取20 μg蛋白上樣于SDS-PAGE分離蛋白,將各蛋白轉至PVDF膜,放入脫脂奶粉(5%)溶液室溫封閉2 h,分別加入各蛋白稀釋比為1∶1 000一抗,4℃孵育過夜,TBST清洗,加入稀釋比為1∶5 000二抗,室溫孵育1 h,采用ECL法檢測免疫復合物表達,其中核β-catenin蛋白以Lamin B為內參,其他蛋白均以β-actin為內參,應用Chemi Imager 5500V2.03軟件分析各蛋白條帶,并采用圖像分析系統計算個條帶整合光密度值,蛋白相對表達量=目的蛋白條帶光密度值/內參照條帶光密度值。
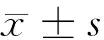
2 結果
2.1 癌旁組和乳腺癌組中miR-429表達水平比較 與癌旁組(1.05±0.20)比較,乳腺癌組的(0.51±0.11)顯著降低,差異有統計學意義(t=20.624,P<0.05)。
2.2 陰性轉染組、MiR-429 mimics組轉染效果檢測倒置熒光顯微鏡下觀察陰性轉染組、miR-429 mimics組細胞轉染效率均>90%。miR-429 mimics組MCF-7細胞中miR-429表達水平顯著高于空白對照組和陰性轉染組(P<0.05)。見圖1,表2。

圖1 倒置顯微鏡下觀測細胞轉染效率(丫啶橙和碘化丙啶染色×100)

表2 3組miR-429表達水平比較
2.3 CCK-8法檢測空白組、陰性轉染組、miR-429 mimics組MCF-7細胞增殖情況 隨著培養時間的延長,miR-429 mimics組MCF-7細胞增殖抑制率顯著升高(P<0.05),不同時間點miR-429 mimics組MCF-7細胞增殖抑制率均顯著高于空白對照組和陰性轉染組,差異有統計學意義(P<0.05)。見表3。
表3 miR-429過表達對MCF-7細胞增殖抑制率的影響
2.4 平板克隆法檢測空白對照組和陰性轉染組、miR-429 mimics組MCF-7細胞增殖情況 與空白對照組相比,陰性轉染組細胞克隆形成率無統計學意義(P>0.05),miR-429 mimcs組克隆細胞形成率顯著降低,差異有統計學意義(P<0.05)。見表4。

表4 3組MCF-7細胞克隆形成率
2.5 Transwell法檢測空白對照組、陰性轉染組、miR-429 mimics組MCF-7細胞侵襲能力 miR-429 mimics組MCF-7細胞侵襲數量均顯著低于空白對照組和陰性轉染組(P<0.05)。見表5,圖2。
表5 miR-429過表達對MCF-7細胞侵襲能力的影響

2.6 空白對照組、陰性轉染組、miR-429 mimics組增殖、侵襲及Wnt通路相關蛋白表達 Western Blot法檢測細胞增殖相關蛋白Ki67、細胞侵襲相關蛋白MMP-2與MMP-9及Wnt信號通路相關蛋白Wnt、GSK-3β、β-Catenin蛋白表達,結果顯示,miR-429 mimics組ki67、MMP-2、MMP-9、Wnt、GSK-3β、β-Catenin蛋白表達均顯著低于空白對照組和陰性轉染組(P<0.05)。見表6,圖3。
表6 MCF-7細胞中ki67、MMP-2、MMP-9、Wnt、GSK-3β、β-Catenin蛋白表達

圖3 MCF-7細胞中ki67、MMP-2、MMP-9、Wnt、GSK-3β、β-Catenin蛋白表達
3 討論
乳腺癌病情發展迅速,目前臨床主要采用手術治療方法,由于乳腺癌細胞增殖、侵襲及浸潤能力強,導致其極易發生轉移[6,7]。因而抑制乳腺癌細胞侵襲對控制其發生、發展及改善患者預后均具有重要意義。miRNA已成為乳腺癌研究重點[8],miR-429是微小RNA-200(miR-200)家族的重要成員,其位于人染色體1p36.33,研究表明miR-429在多種惡性腫瘤中異常表達并可參與腫瘤細胞增殖及侵襲等生物學過程[9,10],但有關miR-429在乳腺癌中研究較少。因此,本研究分析miR-429與乳腺癌發生的關系,并初步探討其可能作用機制。
本研究結果顯示,乳腺癌組織中miR-429表達水平顯著低于癌旁組織(P<0.05),提示miR-429在乳腺癌細胞中呈低表達。倒置熒光顯微鏡鏡下觀察以及qRT-PCR檢測發現miR-429 mimics組miR-429表達水平明顯升高,提示轉染效果佳可用于后續研究。CCK8試驗顯示顯示miR-429過表達組MCF-7細胞增殖抑制率明顯高于空白對照組、陰性轉染組,而MCF-7細胞克隆形成率、細胞侵襲數量均明顯降低,提示miR-429過表達可降低乳腺癌細胞增殖侵襲能力,但具體機制尚不清楚。
本研究結果顯示miR-429 mimics組MCF-7細胞核內β-catenin表達明顯低于空白對照組、陰性轉染組,細胞質內β-catenin表達明顯升高,說明miR-429過表達阻止β-catenin向細胞核內轉移進而抑制EMT發生,類似研究亦證實[11-13],提示miR-429可通過抑制Wnt/β-catenin信號通路進而抑制乳腺癌細胞侵襲。Wnt信號通路其異常活化可促進細胞增殖、分化及侵襲進而導致多種腫瘤發生及發展,癌基因激活或抑癌基因失活均可激活Wnt/β-catenin信號通路進而激活下游靶基因CyclinD1、c-Myc等導致乳腺癌發生及發展,而抑制Wnt/β-catenin信號通路可抑制乳腺癌細胞增殖及侵襲[14-16]。Wnt/β-catenin信號通路激活可抑制GSK-3β活性從而避免β-catenin被其磷酸化導致β-catenin向細胞核內轉移,研究表明β-catenin向細胞核內轉移現象是Wnt/β-catenin信號通路激活的標志[17,18]。然而,腫瘤細胞增殖及侵襲受多種基因調控,研究表明ki67與腫瘤細胞增殖有關并可作為增殖期腫瘤的重要標志物,而MMP-2與MMP-9可作為惡性腫瘤細胞侵襲的標志物[19,20]。本研究結果顯示miR-429 mimics組ki67、MMP-2、MMP-9蛋白表達水平均明顯降低,進一步分析Wnt信號通路相關蛋白表達情況,結果顯示miR-429 mimics組GSK-3β、Wnt蛋白表達水平均明顯低于空白對照組和陰性轉染組,說明miR-429過表達后可明顯抑制Wnt信號通路相關蛋白表達,提示miR-429可能通過抑制Wnt信號通路進而抑制乳腺癌細胞增殖及侵襲。
綜上所述,miR-429過表達可降低乳腺癌細胞增殖、侵襲能力,其可能通過抑制Wnt信號通路發揮作用,可為乳腺癌分子診斷及靶向治療提供理論基礎。本研究存在不足之處:miR-429與Wnt信號通路的具體作用機制及其與其他可能信號通路的關系有待進一步研究。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
電子制作(2018年11期)2018-08-04 03:25:42
祝您健康(2018年5期)2018-05-16 17:10:16
