酵母菌厭氧發酵機制的教學探討
2021-03-14 12:26:46李從虎
安慶師范大學學報(自然科學版) 2021年1期
李從虎,吳 彥
(安慶師范大學生命科學學院,安徽安慶246133)
微生物發酵機理是指人類利用微生物的分解代謝與合成代謝活動,將基質合成人們所需要的產品的內在規律[1],它是發酵工程教學的重點內容。然而,微生物發酵機理的知識涉及碳源代謝、氮源代謝、能量代謝以及復雜的代謝控制流系統,知識抽象、復雜,代謝控制理論很難理解和應用。因此,如何將該知識整合、優化,簡明扼要地傳授給學生是發酵工程教學的當務之急。
在眾多微生物中,酵母菌具有安全性高、碳源利用率較高、代謝產物少和在酸性條件下易于生長繁殖的優良特性[2]。因此,酵母菌在食品、醫藥、生物能源和單細胞蛋白等領域得到廣泛應用[3]。酵母菌的厭氧發酵機制是微生物發酵機理的重要內容之一,掌握該知識對于發酵機理知識的融會貫通具有重要的意義。然而,目前的教材編排主要以產品進行分類,這導致學生難以厘清酵母菌的發酵。如果以酵母菌為主旨,將其發酵機制進行有效串聯,從廣度、深度和學科性上都能促進學生對微生物發酵機理知識的掌握。
1 糖酵解(EMP)途徑
EMP途徑是葡萄糖分解代謝的基本途徑之一,是酵母菌進行發酵生產的基本前提[4]。因此,了解和掌握EMP途徑為學生掌握酵母菌的厭氧發酵機制奠定了理論基礎。圖1為EMP途徑的簡明示意圖。
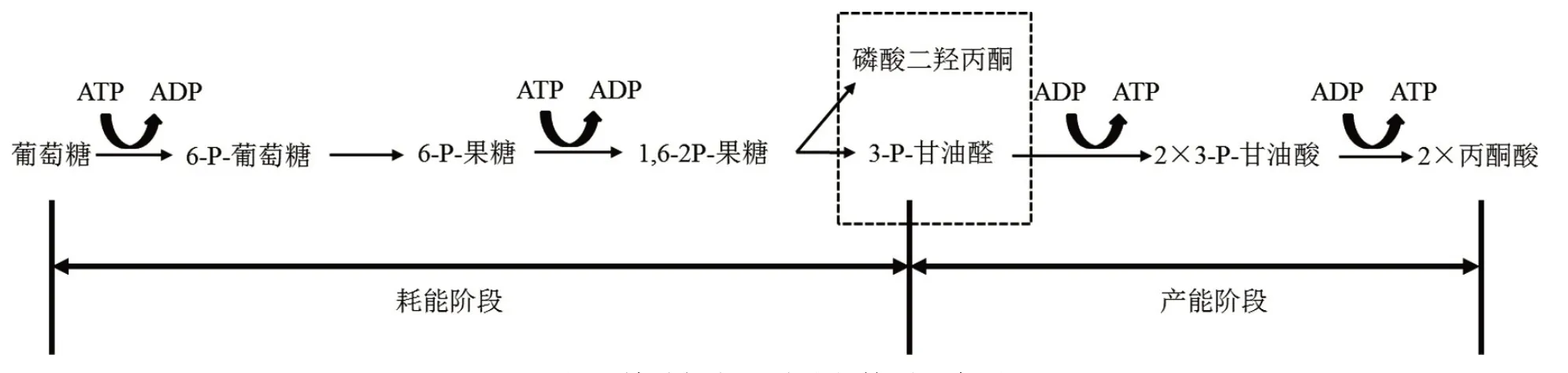
圖1 糖酵解(EMP)途徑簡明示意圖
EMP途徑主要發生在微生物的細胞液中,大致分為兩個階段:第一階段主要是耗能階段,由1分子葡萄糖生成2分子3-P-甘油醛,消耗2分子的ATP;第二階段是產能階段,主要由3-P-甘油醛轉化為丙酮酸的階段,此階段生成4分子的ATP。因此,1分子葡萄糖經EMP途徑凈生成2分子ATP。有氧條件下,丙酮酸進入三羧酸(TCA)循環徹底氧化分解為CO2和水,并生成大量ATP;無氧條件下,丙酮酸進一步被還原成乙醇、甘油、乙酸、乳酸等各種發酵產物,與發酵工業有密切的關系。
己糖激酶、磷酸果糖激酶(限速酶)和丙酮酸激酶是EMP途徑的3個關鍵酶,控制著EMP途徑能否有效進行。EMP途徑的產能雖然較少,但是其關鍵酶的調節仍受到細胞內能量水平的控制。如表1所示,當細胞內產能反應增加時,ATP含量升高,磷酸果糖激酶和丙酮酸激酶的活性被抑制,導致EMP途徑減少;反之,當細胞內耗能反應增加時,ATP被分解為ADP或者AMP,ADP和AMP激活己糖激酶和磷酸果糖激酶,導致6-P-葡萄糖、1,6-2P果糖和3-P-甘油醛的含量增加,這3種中間代謝物含量的增加又激活了丙酮酸激酶的活性,最終促進EMP途徑的增加[1]。此外,6-P-葡萄糖、檸檬酸、NADH和丙氨酸亦分別是己糖激酶、磷酸果糖激酶和丙酮酸激酶的抑制劑,使EMP減少。有報道稱,釀酒酵母進行發酵時,可在1 min內將其細胞的能荷狀態恢復到初始水平[5]。

表1 EMP途徑調節信息
2 酵母菌的酒精發酵
酵母菌的酒精發酵如圖2所示。在掌握了EMP途徑后,我們不難發現酵母菌細胞中酒精的生成來源于EMP途徑中的產物丙酮酸。在丙酮酸脫羧酶(焦磷酸硫胺素為輔酶,并需要Mg2+)的催化下,丙酮酸脫羧生成乙醛和CO2,隨后在乙醇脫氫酶的作用下,乙醛作為受氫體被還原成乙醇。
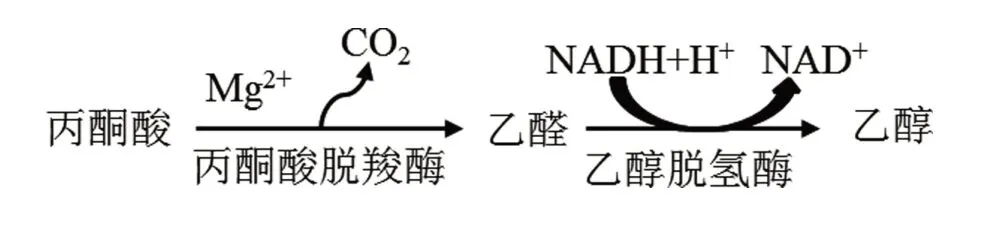
圖2 酵母菌的乙醇生成機制
因此,葡萄糖生成乙醇的總反應式為C6H12O6+2ADP+2H3PO4→2CH3CH2OH+2CO2↑+2ATP。根據此總反應式,1分子葡萄糖在酵母菌細胞內進行分解代謝時,可生成2分子乙醇、2分子CO2和2分子ATP。可見,酵母菌進行酒精發酵生產時,其產能也相對較低。由于葡萄糖和乙醇的相對分子質量分別為180.1和46.05,可計算出乙醇的理論轉化率為[ ( 2×46.05 )/180.1]×100%=51.1%。
在實際生產過程中,由于5%左右的葡萄糖可以用于酵母菌自身組成成分和其他副產物的合成,因此乙醇對葡萄糖的實際轉化率約為理論值的95%。酵母菌的乙醇發酵是酵母菌進行發酵的主體產物之一,其在酒類行業中得以廣泛應用,在教學實際中應結合科技前沿予以補充。如郭燕等人從中高溫大曲中分離出了性能優良的異常威克漢遜酵母Y1、東方假絲酵母Y2以及釀酒酵母Y3和Y4,經過產酒試驗,發現4株酵母的產酒能力從高到低依次為Y3、Y4、Y1、Y2,該研究為提高小曲酒品質和開發大曲中的微生物資源提供了有效路徑[6]。鄭海武等人從釀酒葡萄產區分離得到4株酵母菌,其中WJ1和Q12產酒能力達(10%~11%)vol,與商業酵母菌產酒能力相當,此外還具有明顯的嗜殺性、低泡性和高絮凝性等,符合國標要求,為本土釀酒酵母的多樣性以及葡萄酒的生產研究提供了理論基礎[7]。
3 酵母菌的甘油發酵
3.1 亞硫酸鹽法
正常情況下,酵母菌在厭氧條件下代謝流的主體產物為乙醇,該過程相當徹底。如在進行酵母菌的甘油發酵生產時,加入NaHSO3,乙醛與NaHSO3發生加成反應生成難溶的乙醛加成物,致使乙醇脫氫酶的底物消失,最終導致乙醇無法合成。此時,酵母菌的代謝途徑發生變化,由磷酸二羥丙酮作為氫受體,并在磷酸甘油脫氫酶的催化下生成磷酸甘油,磷酸甘油繼續水解生成甘油,如圖3所示。理論上,1分子葡萄糖可生成1分子甘油。然而,在甘油生產的整個代謝過程中并無ATP的積累,因此,酵母菌無法獲得能量從而導致發酵終止,產物無法生成。為克服這種缺陷,可采取以下措施:(1)厭氧條件下,NaHSO3不能添加太多,保留一部分酒精發酵,以使酵母菌獲得生命所需的能量;(2)有限供氧,使一部分糖代謝進入TCA循環,從而有利于酵母菌甘油發酵,然而由于TCA循環的存在會產生許多中間代謝物,使得甘油的提純較麻煩。為提高釀酒酵母的甘油產量,康毅等人對釀酒酵母D254發酵甘油的條件進行了優化,發現當葡萄糖初始濃度為216 g/L、發酵溫度32℃、pH為3.0時,發酵的甘油產量達0.66 mol/L[8]。因此,如果人為改變條件,控制代謝流的方向,可使產物的形成種類發生改變。這使學生認識到,對內在規律掌握后可熟練進行代謝控制發酵。
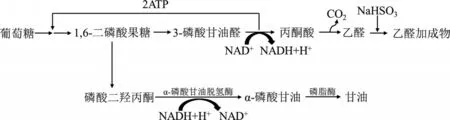
圖3 酵母菌亞硫酸鹽法甘油發酵示意圖
3.2 堿法
酵母菌進行酒精發酵時,如果將發酵液的pH設置在7.6以上時,此時代謝流亦會發生改變。在堿性條件下,此時乙醛不能作為正常的受氫體,因此無法直接被還原成乙醇。然而,2分子乙醛之間可發生歧化反應,相互氧化還原,生成1分子乙醇和1分子乙酸。此外,在另外一條代謝流中,磷酸二羥丙酮作為受氫體,接受3-磷酸甘油醛脫下的氫,進而生成甘油,如圖4所示。因此,2分子葡萄糖在堿法甘油發酵時,主要產物是2分子甘油、1分子乙醇和1分子乙酸[1]。該法與亞硫酸鹽法相似,整個代謝過程并無ATP的積累,因此需采取一定的措施保障酵母菌自身的生命活動,并生產大量的甘油。
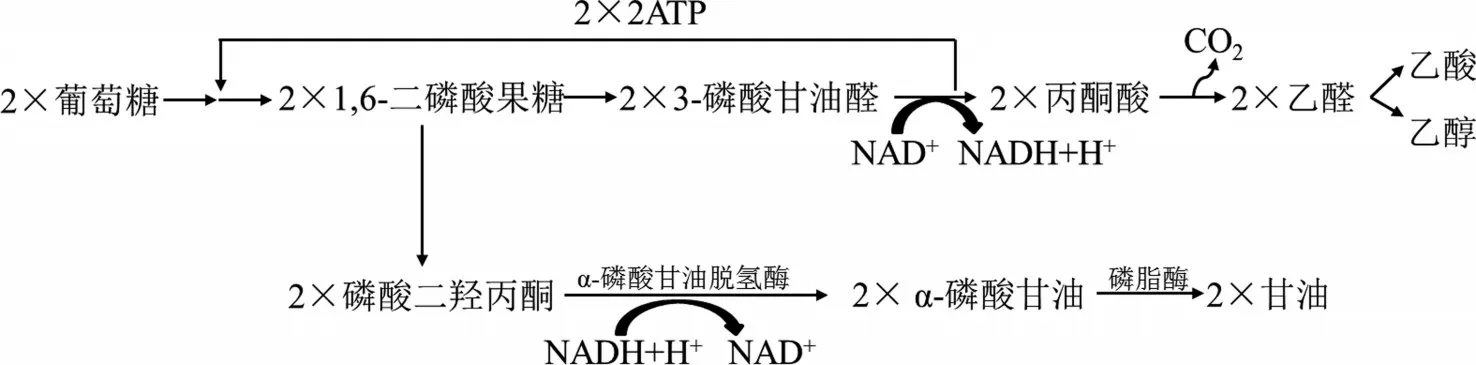
圖4 酵母菌堿法甘油發酵示意圖
4 其他副產物的生成
酵母菌進行酒精發酵時,除了主產物乙醇之外,還生成一些雜醇油、酸類、酯類等副產物,多達40余種,其生成機制如表2所示。這些副產物是構成白酒、黃酒、葡萄酒等酒類風味的重要組成物質,直接影響酒類產品的質量。陳玉香等人通過酵母菌發酵生產紅曲黃酒,發現酵母菌可以代謝合成乙酸異戊酯、丁二酸二乙酯以及乙酸-2-苯乙酯等酯類和苯乙醇、庚醇和丁醇等醇類物質,有效提升黃酒的風味品質[9]。范光森等人從古井貢酒曲中分離出6株酵母菌,其在發酵時均可產生乙酸乙酯、異丁酸己酯、3-甲基-1-丁醇、乙酸、苯甲酸等風味成分,表明酵母菌在白酒釀造過程中可有效增強白酒的風味,提升白酒的品質[10]。

表2 酵母菌發酵時副產物的形成機制
5 結束語
以酵母菌為基本主線,通過人為改變微生物的發酵條件,改變代謝流,可以控制微生物發酵的方向和質量,使有用的發酵產品有效積累,是發酵機理的重要知識點。因此,教師在教學時應盡量簡化代謝流,突出重點,結合科技前沿知識,使抽象的知識具體化、形象化,從而使學生更容易掌握知識點,提升學習品質。
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
中國衛生(2014年12期)2014-11-12 13:12:52
食品工業科技(2014年9期)2014-03-11 18:15:31
