人類與獼猴大腦的同源腦區(qū)研究綜述
2021-03-19 06:27:28王千山柴靜文李斌強李海芳
太原理工大學學報 2021年2期
王千山,王 玥,柴靜文,李斌強,李海芳
(太原理工大學 信息與計算機學院,太原 030024)
理解大腦結(jié)構(gòu)、發(fā)掘大腦工作模式,對解構(gòu)人類思維、意識、認知等方面具有重大意義。探索大腦的結(jié)構(gòu)和功能是21世紀最具挑戰(zhàn)性的前沿學科[1]。2013年美國和歐盟分別提出BRAIN和HBP大腦研究計劃,日本于2014年啟動了“腦智計劃”,我國在《國家中長期科學和技術(shù)發(fā)展規(guī)劃綱要(2006-2020)》將“腦科學與認知”列入基礎(chǔ)研究8個科學前沿問題之一。大腦研究計劃的重要目的是揭示大腦的工作原理和疾病發(fā)生機制,帶動相關(guān)學科及其上下游產(chǎn)業(yè)的共同發(fā)展。
人體為對象的各類實驗受到嚴格的限制。國際通行的《紐倫堡法典》《赫爾辛基宣言》以及我國《涉及人的生物醫(yī)學研究倫理審査辦法(試行)》中都對人體實驗設(shè)置了各類規(guī)范和限制。最新頒布的《民法典》中對人體實驗提出明確的法律規(guī)定。在開展人體實驗的過程中,不僅要關(guān)注倫理問題,實驗對象的生命安全更應(yīng)放在首位。因此侵入式的創(chuàng)傷性實驗的開展,要經(jīng)過反復(fù)論證,并在動物模型上試驗通過后再評估能否進行人體實驗。而且考慮到示蹤藥物對于人體的影響,諸如CT、PET等非侵入式的實驗也不能在受試者上反復(fù)進行。此外,民眾普遍對輻射較為敏感,因此即便是對人體影響最小的磁共振技術(shù),受試者往往也不愿配合進行多次實驗。人體實驗難以開展的情況很大程度上限制了腦科學的發(fā)展。
動物大腦研究是研究人類大腦的重要途徑,借助動物大腦可以研究人類大腦的工作機制和腦疾病的發(fā)展規(guī)律。獼猴在中國具有得天獨厚的資源優(yōu)勢,隨著體細胞克隆猴和轉(zhuǎn)基因獼猴模型的相繼突破,獼猴作為焦點模式實驗動物的地位進一步強化。雖然非人靈長類動物具有和人類相似的大腦組織結(jié)構(gòu)和保守特征,但在生物演化過程中仍出現(xiàn)了形態(tài)、連接、功能等方面的差異[2]。因此將動物實驗的結(jié)果推廣到人類還需要進一步研判。
同源腦區(qū)是指具有相同演化先祖、擁有相近或相同功能的腦區(qū),能夠為跨物種研究提供相對可靠的推測依據(jù),為大腦組織方式研究提供獨特解讀依據(jù)[2]。現(xiàn)代醫(yī)學研究得到的人類和獼猴已知腦圖譜對于大腦區(qū)域的劃分多達數(shù)百個,但尚不存在某兩個圖譜能夠?qū)⑷四X和獼猴腦區(qū)完整對應(yīng)。且目前人類與獼猴的大腦同源腦區(qū)研究中,缺少對已證明的同源腦區(qū)的全面總結(jié)和綜合研究記錄。對同源腦區(qū)進行梳理總結(jié)對制作精確的人類和獼猴腦區(qū)映射關(guān)系圖譜、探究大腦工作機制和腦疾病的發(fā)展規(guī)律具有重大意義。
本文收集整理2000年以來涉及人類和獼猴同源腦區(qū)選取的相關(guān)研究資料。尋找大腦分區(qū)數(shù)目明確、適用于計算機分析、有望為人類和獼猴大腦同源區(qū)研究提供較好幫助的腦圖譜或腦模板;選取被明確證實的同源腦區(qū)或在跨物種腦區(qū)功能研究中使用的腦區(qū),并根據(jù)其所涉及的具體功能進行歸類總結(jié)。目的是回顧同源腦區(qū)的研究過程,總結(jié)已有的同源腦區(qū)和其研究進展,為新的同源腦區(qū)的研究找到理論支撐,為大腦高級認知功能研究奠定理論基礎(chǔ),以期推動人腦工作機制和腦疾病發(fā)展規(guī)律的相關(guān)研究。
1 兩物種腦圖譜匹配
人類腦圖譜發(fā)展至今數(shù)量眾多,代表性的是以MNI152標準空間作為腦模板[3],先后誕生的諸如哈佛-牛津圖譜[4]、AAL模板[5]、Brainnetome人腦連接組圖譜[6]以及Julich腦圖譜[7]。相較于人類腦圖譜,獼猴腦圖譜數(shù)量較少。伴隨影像學的發(fā)展,非人靈長類動物的腦影像數(shù)據(jù)的不斷積累,基于MRI的非人靈長類動物的腦模板、腦圖譜陸續(xù)被提出。特別是如PRIME-DE此類項目,提供了大量的多平臺多中心的獼猴非人靈長類動物腦影像數(shù)據(jù)集[8]。
由于人類和獼猴大腦間已探知的同源組織數(shù)量和精細度嚴重不足,因此跨物種的大腦研究仍停留在低層次腦區(qū)的表征和比較層面,這樣就限制了腦區(qū)同源性研究的發(fā)展。目前,一些研究團隊已開始此方面的初步探索,如華盛頓大學醫(yī)學院Van Essen實驗室基于形態(tài)標記約束的表面配準過程實現(xiàn)兩個物種在形態(tài)學上的體素拓撲映射[9],牛津大學納菲爾德腦功能磁共振成像中心嘗試提供一些連接模式上存在最佳對應(yīng)關(guān)系的位點[10-12]。由此可見,構(gòu)建能夠逼近人類和獼猴真實同源關(guān)系的大腦體素級映射關(guān)系的腦圖譜有著十分重要的價值。
獼猴腦圖譜研究中,主要以FOX和LANCASTER[13]、PAXINOS et al[14]、SALEEM和LOGOTHETIS[15]提出的猴腦圖譜標記作為標準,為后續(xù)腦圖譜開發(fā)提供重要的分區(qū)依據(jù)。此后,2009年DUBACH et al[16]繪制了NeuroMaps三維獼猴腦圖譜,2011年FREY et al[17]采用人腦MNI空間構(gòu)建方法,將Paxinos繪制的獼猴圖譜標記配準到MNI空間從而提出猴腦MNI腦模板。基于MNI猴腦模板ROHLFING et al[18]在2012年制作了INIA19腦模板,并將NeuroMaps獼猴圖譜手工繪制其上。同年,VAN ESSEN et al[9]基于表面圖集,在早期D99腦模板上繪制了F99的皮質(zhì)表面圖譜。2015年,為更好地使用連接度量分析,Duke大學的KOLSTER et al[19]使用彌散磁共振數(shù)據(jù),結(jié)合Paxinos獼猴圖譜和MNI標準空間,繪制基于彌散數(shù)據(jù)的CIVM MRI/DTI atlas獼猴腦圖譜。DONAHUE et al[20]于2016年同期發(fā)布基于彌散數(shù)據(jù)的Yerkes19獼猴腦圖譜。2017年,REVELEY et al[21]在早期D99腦模板的基礎(chǔ)上,結(jié)合前人的工作,繪制了獼猴3D腦圖譜。2018年,SEIDLITZ et al[22]結(jié)合此前工作,將LOGOTHETIS繪制的圖譜中248個腦區(qū)配準其上發(fā)布了NMT腦圖譜。隨后JUNG et al[23]在其NMT的基礎(chǔ)上引入恒河猴皮層層次圖集(CHARM),制作了NMTv2腦圖譜。本文檢索大量腦圖譜并研究其圖譜屬性,從而給出兩類人類和獼猴大腦圖譜的最佳匹配結(jié)果,兩物種大腦圖譜的匹配流程如圖1所示。
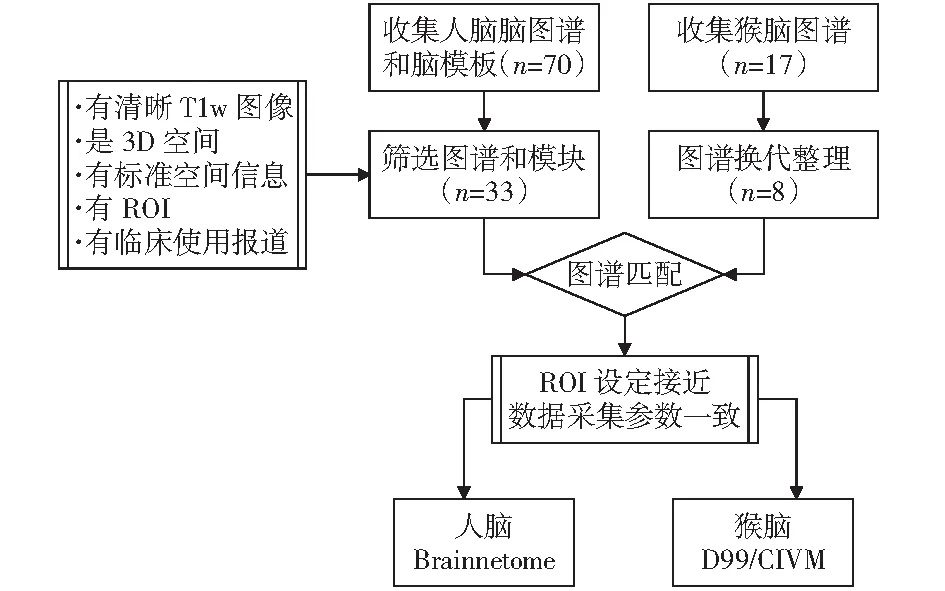
圖1 選取匹配圖譜
1.1 大腦皮層組織圖譜
基于形態(tài)學標記約束的配準方法是構(gòu)建人類和獼猴大腦對應(yīng)關(guān)系的重要途徑。該方法的準確性依賴于大腦皮層組織圖譜的精細程度和配準軟件的適應(yīng)能力。在實際比對后發(fā)現(xiàn),人類腦圖譜在FSL、FreeSurfer、SPM等軟件中,使用各自軟件內(nèi)置的腦圖譜均可以產(chǎn)生較好的配準效果。在AFNI中使用D99腦模板或FreeSurfer中使用F99腦模板能夠得到相對較好配準結(jié)果。通過對腦區(qū)位置和數(shù)量進行詳細比對,發(fā)現(xiàn)人腦Brainnetome連接組圖譜提供的246個區(qū)域、Julich-Brain Atlas標記的248個區(qū)域能夠同2017年更新的猴腦D99腦模板的254個區(qū)域構(gòu)成一定的對應(yīng)關(guān)系。
1.2 彌散磁共振圖譜
彌散張量成像(DTI)技術(shù)能夠較好的觀察大腦內(nèi)部白質(zhì)纖維走向。通過使用彌散磁共振技術(shù)已經(jīng)圍繞人腦中運動皮質(zhì)區(qū)域建立起感覺運動區(qū)模板(SMAAT).圍繞猴腦展開的圖譜研究,目前還未有同人腦中SMAAT圖譜相似的白質(zhì)纖維束圖譜。Brainnetome連接組圖譜中提供了基于彌散態(tài)的圖譜信息,其中的246個區(qū)域能夠同Duke大學的CIVM MRI/DTI 圖譜提供的241個區(qū)域在一定程度上形成對應(yīng)關(guān)系。為猴腦計算制作全腦的連接組圖譜,勢必能夠為構(gòu)建人腦和獼猴大腦對應(yīng)關(guān)系提供有力的支持。
2 已知的同源腦區(qū)
基于圖論的復(fù)雜網(wǎng)絡(luò)分析極大地推進了對于人類腦網(wǎng)絡(luò)組織模式的認識。構(gòu)建的網(wǎng)絡(luò)中的節(jié)點數(shù)和網(wǎng)絡(luò)復(fù)雜度,很大程度上依賴于腦圖譜對于腦組織的區(qū)域劃分結(jié)果[24]。特別在MARS et al[25]提出基于連接指紋圖的方式進行跨物種比較后,同源腦區(qū)的選定變得更加重要。
雖然人類與非靈長類動物的大腦結(jié)構(gòu)具有相似性,但在人類的進化過程中,出現(xiàn)了諸如大腦半球偏側(cè)化、腦組織體積膨脹、功能轉(zhuǎn)移等情況[26],導(dǎo)致跨物種比較中不能僅以大腦組織的宏觀形態(tài)標記劃分感興趣區(qū)域,以腦區(qū)的同源性為根據(jù),進行興趣區(qū)域的篩選,有助于更精確地分析大腦結(jié)構(gòu)和工作原理。
同源腦區(qū)的定義至今不夠完善,研究者對同源組織進行過一些總結(jié),如:華盛頓大學醫(yī)學院Van Essen實驗室總結(jié)利用55對大腦皮層同源腦區(qū)用于表面配準[27],牛津大學納菲爾德腦功能磁共振成像中心預(yù)定義出22對同源腦區(qū)來比較人類和獼猴前額葉皮層位點的功能連接模式[10],隨后的研究中MARS et al[28]僅通過預(yù)定義5對同源腦區(qū)研究了輔助運動區(qū)(SMA)、前輔助運動區(qū)(PreSMA)以及Area9、10等腦區(qū)的大腦同源性。中國電子科技大學生命科學與技術(shù)學院的WANG et al[29]預(yù)定義了17對同源腦區(qū)來比對人類和獼猴的PCun結(jié)構(gòu)。這些研究都是帶著特定目的粗略總結(jié),詳細的同源組織還需要專門系統(tǒng)總結(jié)。
同源區(qū)的研究主要通過電刺激效應(yīng)分析、細胞構(gòu)筑、解剖分析、連通性分析等四類方法開展[30-34]。如CASPERS et al[35-37]在2006-2013年間,先后受體細胞構(gòu)筑、解剖分析、功能連接等方法,驗證了人類和猴類大腦中頂葉部分中與認知相關(guān)的部分腦區(qū)之間的同源關(guān)系。JOHANSEN-BERG、MORECRAFT et al、SALLET et al先后使用解剖分析和細胞構(gòu)筑的方法驗證了人類和猴類大腦中輔助運動區(qū)(SMA)的同源關(guān)系[38-40]。MARS et al發(fā)表的基于結(jié)構(gòu)連接的同源性研究方法在近年逐漸成為研究關(guān)注熱點[28]。
本文根據(jù)人類大腦的一般工作過程,將同源腦區(qū)依照同感覺過程、行動過程、言語過程、認知過程四類處理過程的相關(guān)性進行分類。腦區(qū)同源關(guān)系確定過程如圖2所示,通過查閱相關(guān)腦區(qū)的研究報道,篩選出至少包含3種同源區(qū)研究方法相關(guān)記載的腦區(qū)作為有效的同源腦區(qū),為同源腦區(qū)研究提供相關(guān)的理論基礎(chǔ)。
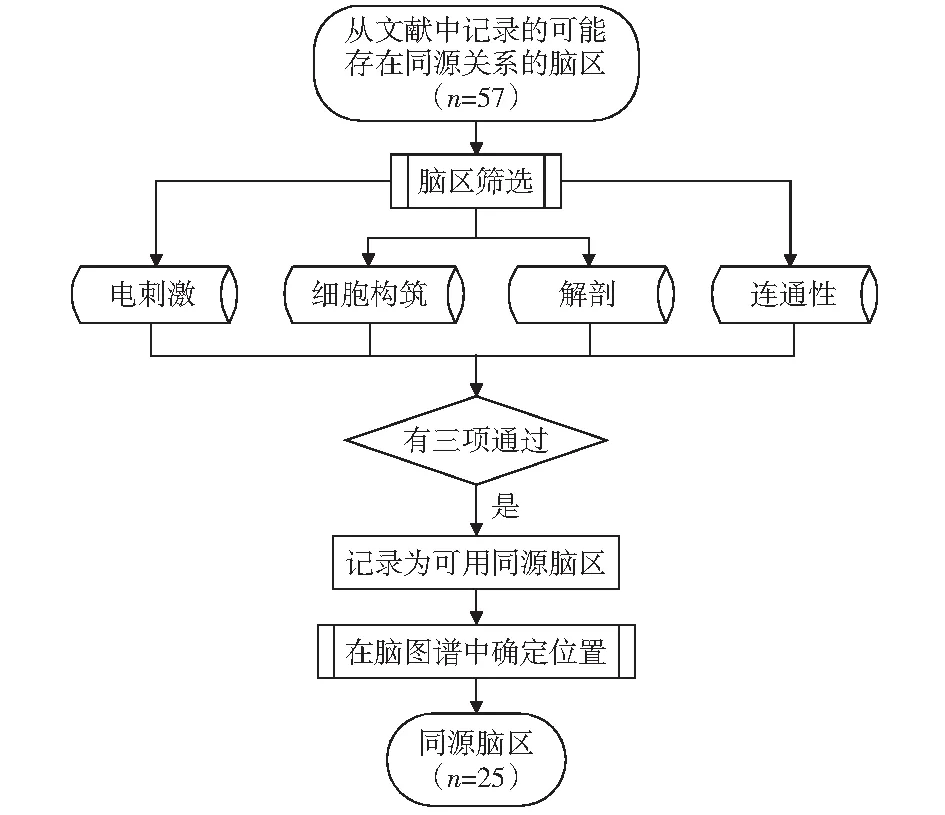
圖2 同源腦區(qū)選取
2.1 感覺過程相關(guān)
目前腦區(qū)同源性研究主要集中在以視覺為代表的感覺過程相關(guān)腦區(qū)。靈長類動物通過視覺獲取了大量的外部環(huán)境信息,這些信息經(jīng)過加工整理,驅(qū)動人體產(chǎn)生各類具體反應(yīng)。
1) 人類的外側(cè)枕葉皮質(zhì)(LOC)同獼猴LOC部分同源[31-34]。
2) 人類大腦中位于海馬旁回附近的嗅周皮質(zhì)區(qū)域(Perirhinal)同獼猴35、36腦區(qū)同源[41-43]。
3) 人類顳極(TP)皮質(zhì)同獼猴TG區(qū)同源[32,43-45]。
4) 人類頂內(nèi)溝腹側(cè)(VIP)同獼猴VIP同源[45]。
5) 人類輔助運動前側(cè)(preSMA)同獼猴的F6區(qū)域同源[25]。
2.2 動作過程相關(guān)
動作相關(guān)的同源腦區(qū)研究在眾多腦區(qū)同源性研究中最早開始。動物都具有一定的行為動作能力,是區(qū)別于植物的主要特征。動作是包含表情變化和動作變化,以及由肢體動作變化導(dǎo)致的本體在空間上能夠發(fā)生的位置變化。而動作的完成,主要是由中樞神經(jīng)系統(tǒng)有規(guī)律的控制、協(xié)調(diào)各個肌肉群配合完成的。動作作為一種基本能力,在漫長的演化過程中得到了長期的保留,相關(guān)腦區(qū)的同源性較容易得到證實。
1) 人類和獼猴同位于前連合冠狀面后面的內(nèi)側(cè)額上回的輔助運動區(qū)[38-40]。
2) 人類和獼猴的初級運動皮質(zhì)區(qū)(M1)[46-47]。
3) 人類和獼猴的初級體感皮質(zhì)區(qū)(S1)[38]。
2.3 言語過程相關(guān)
語言的復(fù)雜結(jié)構(gòu)使得其表達范圍比任何已知的動物交流系統(tǒng)都要廣泛。語言包含視覺、聽覺、觸覺等多個方面,言語系統(tǒng)的表達雖然在不同生物間有很大差異,但依舊可以找到其表達過程的共有特征。
1) 人類額中回前部和額上溝的交界區(qū)(aMFG+SFS)與獼猴9/46區(qū)[39,48-50]。
2) 人類額上回下延至旁扣帶回區(qū)域(sSFG+Cingulum)與獼猴8B/8Bm區(qū)[39,51-52]。
3) 人類額下回腹側(cè)(vIFG)與獼猴44v區(qū)[53-56]。
2.4 認知過程相關(guān)
認知是人類獲取并加工信息的過程。這一能力是人類與其他靈長類動物的主要差異。在該研究領(lǐng)域中,研究人員既希望找到更多的同源特性論證,以輔助藥物、精神疾病研究,同時也急切的希望找到更多的差異區(qū)域,借此解開人類智慧的奧秘。因而此類區(qū)域集中了大量的研究工作。與認知相關(guān)的同源腦區(qū)有:
1) 人類頂下小葉前部和島葉外側(cè)區(qū)域(PFop)同獼猴大腦的PFop/7op區(qū)域[26,35-37]。
2) 人類頂下溝前部(aIPS)同獼猴大腦的頂內(nèi)溝前部(AIP)[26,35-37].
3) 人類頂下溝后部(pIPS)同獼猴大腦的頂下溝后部(pIPS)[26,35-37].
4) 人類頂下小葉頂部(PG)同獼猴大腦的頂下小葉的PG/7a區(qū)[26,35-37]。
5) 人類扣帶回(PC)后部同獼猴大腦扣帶回后部(pPC)[38,57-59].
6) 人類扣帶回皮質(zhì)的后皮質(zhì)區(qū)域(rsplC)同獼猴23ab區(qū)[38,57-59]。
7) 人類前扣帶回膝部(pgACC)同獼猴pgACC區(qū)[10,60-61]。
除上述區(qū)域外,還有一些結(jié)構(gòu)清晰、功能明確的腦區(qū),在早期研究中得到明確同源論證結(jié)果。此類腦區(qū)有腹側(cè)紋狀體(ventrStr)、尾狀核頭部(Caud)、殼核(putamen)、海馬體(hippoc)、杏仁核(amygd)、下丘腦(hypoth)、腹側(cè)被蓋區(qū)(VTA)[62-65].
本文梳理出的25對同源腦區(qū)在MNI標準空間中的位置如圖3所示,人腦和獼猴同源腦區(qū)在圖譜中名稱的對應(yīng)關(guān)系詳見表1.
通過梳理文中涉及的同源腦區(qū)相關(guān)研究工作發(fā)現(xiàn),人類Perirhinal皮質(zhì)同獼猴35、36腦區(qū)的同源研究中,尚未使用纖維追蹤判斷連通性的方式論證其同源性;人類和獼猴的初級運動皮質(zhì)區(qū)(M1)同源研究中,盡管人類已經(jīng)構(gòu)建了SMAAT圖譜以描述人類大腦各運動單元的白質(zhì)纖維連接情況,但兩者的連通性分析仍未進行。人類額下回腹側(cè)與獼猴44v區(qū)域同源研究關(guān)聯(lián)到Broca區(qū)的研究分析,已在細胞構(gòu)筑和連通性分析方面進行了詳細論述,但功能連接分析沒有進行。人類扣帶回(PC)后部同獼猴大腦扣帶回后部同源性研究情況也存在類似功能連接分析尚未論述的情況。
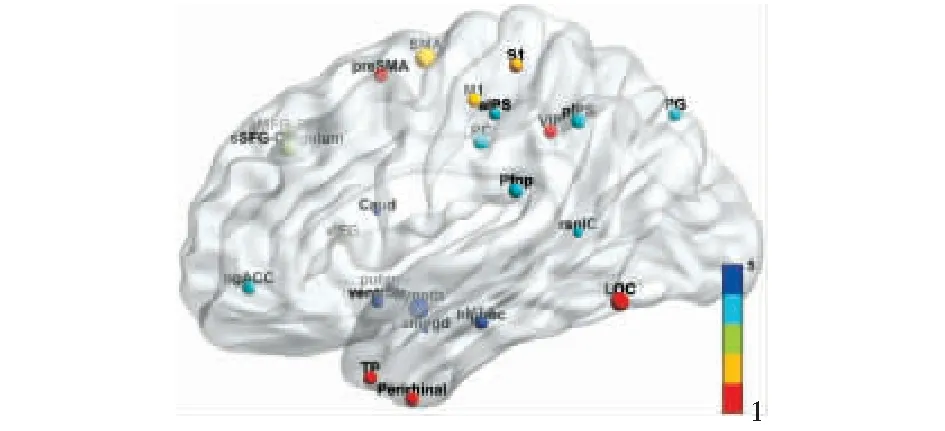
圖3 同源腦區(qū)位置示意

表1 已知同源腦區(qū)的人猴對應(yīng)關(guān)系表
目前仍有大量的腦區(qū)同源性有待證實。猴腦腦區(qū)在PAXINOS[14]和LOGOTHETIS et al[15]提出的猴腦圖譜中均被手工標記了200個以上的大腦腦區(qū)。如認知控制和語言功能相關(guān)的45區(qū)、47/12區(qū);社會認知和面部識別的vmPFC,14m區(qū);對于獎懲敏感的FPm區(qū)域的研究;參與到空間注意、語言和高級運動加工的腦區(qū)如PF,PG,IPL,Broca等區(qū)域的同源性研究都有待更多的科學工作者揭示其中原理。同時,大量有望能夠為藥物靶點治療提供更多可能的腦區(qū)的功能同源性分析仍亟待開展。
值得注意的是,人的動作、言語、認知過程均不是孤立發(fā)生的,各類的行為是在基于對感覺通路獲得的大量信息進行有效處理后,經(jīng)過復(fù)雜決策過程后進行的生理活動。特別是認知過程,要整合加工各類型信息,并在不同腦區(qū)間進行復(fù)雜的信息傳遞,而不是簡單的在某個腦區(qū)內(nèi)直接完成的。這就導(dǎo)致感覺通路上的各個腦區(qū)同大量其他活動相關(guān)的腦區(qū)都存在一定程度的聯(lián)系。即使是對面孔的識別也存在由基本視覺區(qū)域到多個不同功能腦區(qū)的投射。因此在分析各種類型的腦區(qū)活動時,都應(yīng)將感覺過程相關(guān)的同源腦區(qū)作為重要的信息輸入?yún)⒖家氲较嚓P(guān)的計算和分析。
3 總結(jié)
目前國內(nèi)開展的腦區(qū)同源性研究已得到很多成果,但在人類與非人靈長類動物方面還在發(fā)展階段。多圖譜的腦區(qū)識別方法能夠為同源腦區(qū)研究提供更多信息,本文中提及的25對同源腦區(qū)在D99獼猴腦圖譜中僅能有效的對應(yīng)13個腦區(qū),其他腦區(qū)在INIA19、CIVM圖譜中予以尋找完整。
國內(nèi)人類腦連接組圖譜研究居國際領(lǐng)先地位,轉(zhuǎn)基因獼猴模型制作也屬前列,但仍缺乏高清的非人靈長類腦圖譜。隨著新型7T磁共振設(shè)備的推廣[66],大量3T/7T磁共振數(shù)據(jù)勢必大量產(chǎn)生,獼猴腦影像數(shù)的不斷積累,可促使獼猴腦連接組圖譜的產(chǎn)生,對非人靈長類圖譜研究起重要推動作用。構(gòu)建能夠清晰反映人類和獼猴大腦體素級映射關(guān)系圖譜能夠有效推動大腦工作機制探究和腦疾病的研究進展。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
大科技·百科新說(2021年6期)2021-09-12 02:37:27
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
好孩子畫報(2020年5期)2020-06-27 14:08:05
科技傳播(2019年22期)2020-01-14 03:06:54
意林·全彩Color(2019年6期)2019-07-24 08:13:50
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導(dǎo)刊(2016年34期)2016-04-11 00:49:44
奧秘(2015年2期)2015-09-10 07:22:44
