AMETH_3452基因在擬無枝酸菌中的系統發育分析
2021-04-01 07:07:34唐標吳靜鄭華軍趙維楊華
浙江農業科學 2021年4期
關鍵詞:分析
唐標,吳靜,鄭華軍,趙維,楊華*
(1.浙江省農業科學院 農產品質量安全與營養研究所,浙江 杭州 310021; 2.上海人類基因組研究中心,上海 201203;3.中國科學院 深圳先進技術研究院,廣東 深圳 518055)
擬無枝酸菌(Amycolatopsis)大部分的菌株都是從土壤中分離,部分從海洋、沙漠、沉積物、臨床樣本等分離,歸屬于放線菌綱[1-2]、假諾卡氏菌目、假諾卡氏菌科[3-4]。近年來,該屬菌種數量逐漸增加,迄今為止已有82個有效發表的典型菌種(https://lpsn.dsmz.de/genus/amycolatopsis)。該屬的菌株可通過gyrB、recN和16S rDNA基因分劃為多個分支[5-6]。該屬菌種分為2個主要分支,東方擬無枝酸菌分支(Amycolatopsisorientalissubclade,AOS)和甲基擬無枝酸菌分支(Amycolatopsismethanolicasubclade,AMS)[5-8]。其中,AOS的菌株常被發現可以生產抗生素,例如A.orientalis生產萬古霉素[9]、Amycolatopsismediterranei生產利福霉素[10-11]等。AMS的代表菌株A.methanolica是一個兼性甲基營養放線菌,未發現產生抗生素,且次級代謝物較少[8-12]。AMS中的菌種能夠在45 ℃以上的條件下生長,屬于嗜熱放線菌[12-13]。后經過gyrB-recN-16S的串聯核酸序列,將該屬的35個菌種分為A~F分支[6]。然而,由于采用不同的序列及方法,獲得的系統發育模式并不相同,因此,該屬菌種的系統發育地位仍需進一步探究[14]。
全基因組數據目前已經被越來越多的用來分析原核細菌之間的進化關系,包括一些傳統方法很難區分的放線菌[15]。基因組序列提供了更多的潛在分子標記用于系統發育。前期的研究在A.methanolica中發現了放線菌綱中普遍存在的特有基因AMETH_3452,定位在擬無枝酸菌屬環形基因組的復制終點附近[8]。但是該基因的生理特征和應用前景尚不明確。
本研究對AMETH_3452基因進行了蛋白表達分析,對可能存在的保守位點進行了解析,利用AMETH_3452蛋白序列對擬無枝酸菌屬內76個菌株的同源序列進行了系統發育分析,為該基因的功能解析和潛在的檢測方法應用提供了基礎。
1 材料與方法
1.1 菌株培養和基因組抽提
菌株A.methanolica239來自本實驗室保藏,使用高天瓊脂培養基(20 g·L-1淀粉,0.5 g·L-1L-天冬氨酸,1 g·L-1硝酸鉀,0.5 g·L-1三水合磷酸氫二鉀,0.5 g·L-1氯化鈉,0.5 g·L-1七水硫酸鎂,1 g·L-1氯化鈣和15 g·L-1瓊脂粉,pH 7.5)在30 ℃條件下復蘇培養2 d,隨后轉接至ISP2液體培養基(0.4%酵母浸出物,1%麥芽浸出物和0.4%葡萄糖)在30 ℃條件下培養2 d。使用細菌基因組提取試劑盒(天根生化科技有限公司),按照說明書提取基因組DNA。
1.2 質粒構建與蛋白表達
載體pET-28a(5.3 kb,kan+)用NdeⅠ/HindⅢ雙酶切、去磷、膠回收純化后,以A.methanolica239基因組為模板,用高保真聚合酶phanta(南京諾唯贊生物科技股份有限公司)進行PCR擴增(F:5′-AGCCATATGGTGCGCTCCCACA GCTATGAC-3′,R:5′-CGCAAGCTTTCAGCCTTCCG GGTCGGTCAC3′)后純化。用NdeⅠ/HindⅢ雙酶切回收產物進行黏性末端連接,轉化大腸桿菌DH10B菌株,37 ℃培養,篩選得到陽性轉化子。將構建成功的質粒和對照質粒pET-28轉化到大腸桿菌Rosetta(DE3)中,37 ℃過夜培養后獲得單克隆。接種單克隆至5 mL液體LB培養基中,30 ℃過夜培養,按1%比例轉接到50 mL的LB液體培養基中,30 ℃培養至D600約為0.8時加入終濃度為0.2 mmol的IPTG,調整搖床溫度至16 ℃過夜培養。離心收集菌體,超聲裂解細胞,分離上清和沉淀,其中沉淀用50 mL TE緩沖液重懸,最后分別取樣進行蛋白電泳。
1.3 基因與蛋白序列分析
AMETH_3452蛋白二級結構預測使用PSIPRED server[16](http://bioinf.cs.ucl.ac.uk/psipred/);生理特性預測使用Expasy(http://web.expasy.org/protparam/);蛋白質跨膜區預測使用TMHMM,(http://www.cbs.dtu.dk/services/TMHMM/);信號肽的預測使用SignalP[17](http://www.cbs.dtu.dk/services/SignalP)。AMETH_3452基因在擬無枝酸菌中的同源序列參考之前的報道[8],對位分析使用Clustal W,去除非對位堿基后,使用BioEdit進行保守區可視化。
1.4 系統發育分析
AMETH_3452的同源蛋白序列通過在GenBank的NR庫上BLAST檢索直系同源蛋白,選擇所有全長匹配(匹配長度>95%)的擬無枝酸菌的直系同源蛋白。使用Clustal W進行比對,通過MEGA X軟件[18]基于最大似然法(Maximum-likelihood method)構建系統發育樹,進化樹分支模式的穩定性分析采用bootstrap法,重復次數為1 000。外類群使用Pseudonocardiadioxanivorans菌株的同源蛋白。
2 結果與分析
2.1 AMETH_3452蛋白和編碼基因的序列特征
通過對18個擬無枝酸菌不同種的基因序列對比發現,該基因的全長在792~861 bp,不同的基因序列同源性在79.23%~99.02%。通過比對分析后可以看出,在300~500 bp的位置序列較為保守(圖1中A),可以作為該屬的分子標記,用于核酸檢測。該蛋白序列通過預測后發現不穩定系數為35.66(<40),說明蛋白性質穩定,親水性為-0.177,說明可溶性不好。預測到TMHs(跨膜螺旋)個數為0,即不存在跨膜區,未預測到信號肽區域。在115位氨基酸之前主要的二級結構以β-折疊為主,而在之后,α-螺旋增加(圖1中B)。另外,在蛋白的75~115位預測到結構域,可能與水解酶相關。
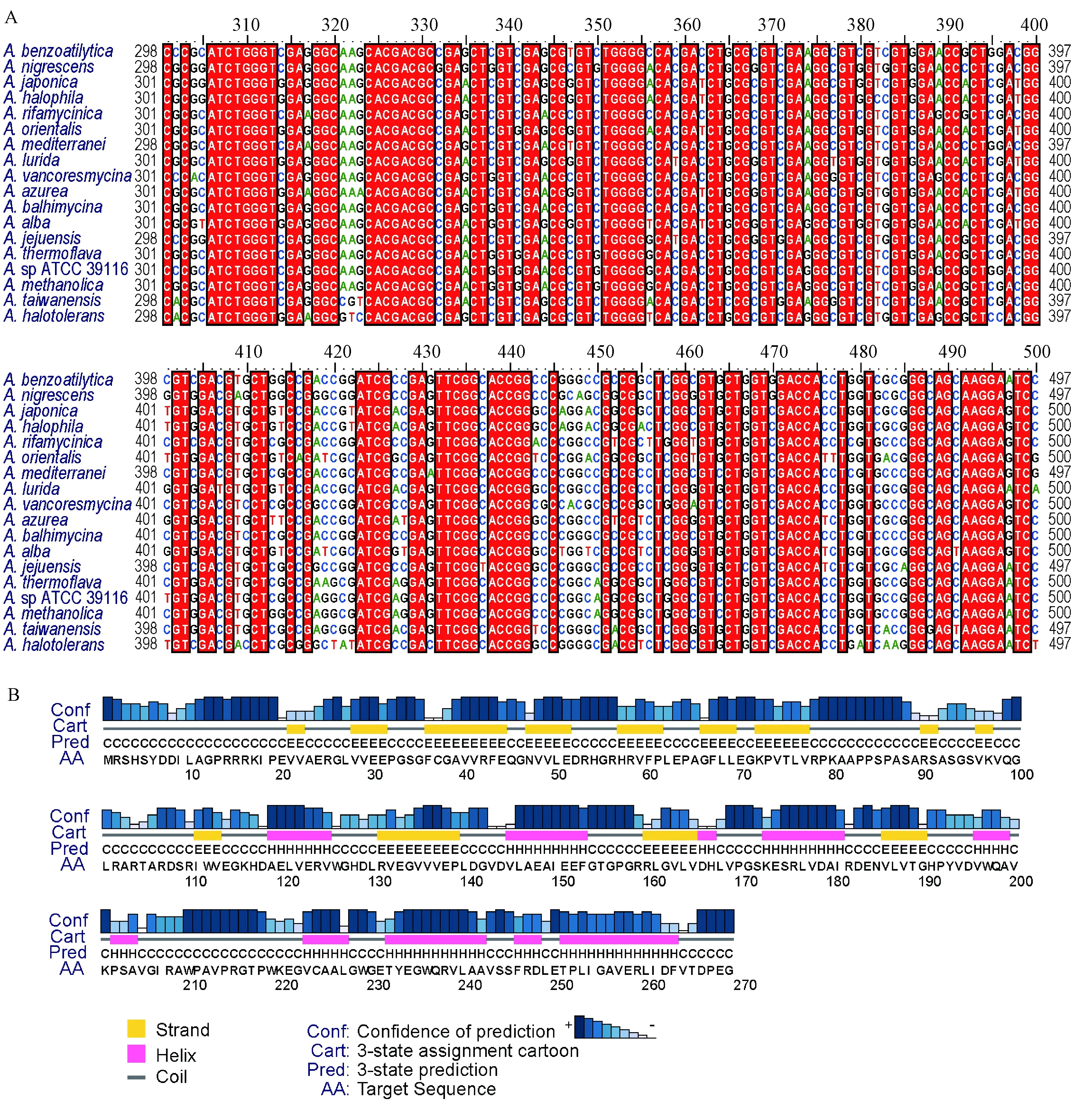
圖1 AMETH_3452蛋白二級結構預測與AMETH_3452基因保守性分析
2.2 AMETH_3452表達載體構建與蛋白表達
AMETH_3452基因擴增后,通過NdeⅠ/HindⅢ雙酶切后黏性末端連接將基因序列構建在表達載體pET-28a上,形成表達質粒pTC17260,總長度為6 121 bp。AMETH_3452蛋白大小為29.2 KD,該蛋白和His標簽融合表達(圖2中A)。表達宿主加入IPTG誘導,裂解細菌通過蛋白電泳后發現,AMETH_3452蛋白有表達,并位于正確的可見條帶上(圖2中B)。在目標產物的下方,仍有較明顯的蛋白條帶產生,可能存在其他基因被誘導表達的情況。另外,全細胞破碎后電泳,發現表達出的蛋白以包涵體的形式富集在沉淀中,但是仍有較多存在于上清中,為后期蛋白純化提供了有利條件。
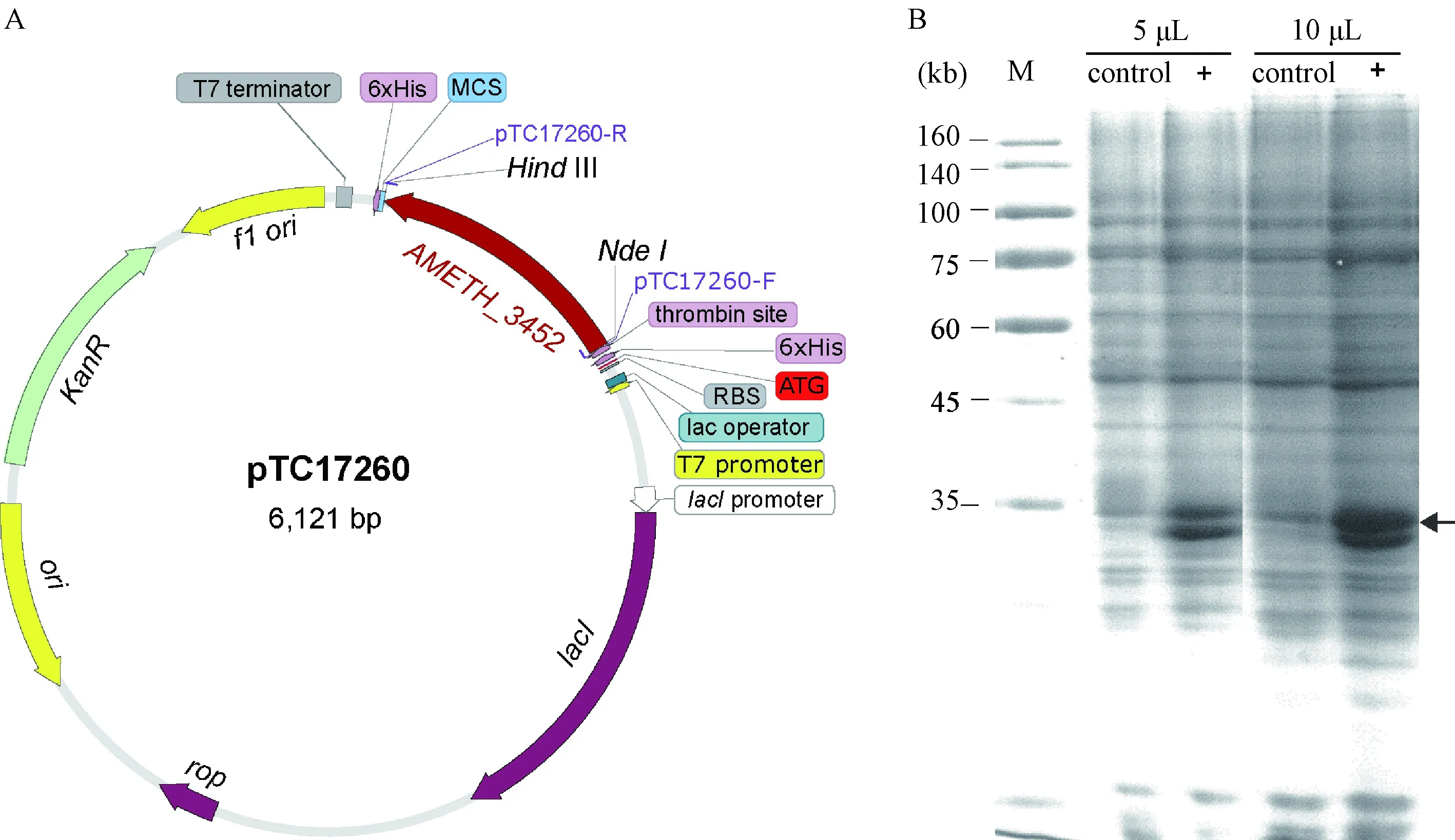
圖2 AMETH_3452蛋白表達質粒構建與表達宿主上清液蛋白電泳
2.3 AMETH_3452蛋白序列的系統發育分析
將AMETH_3452蛋白在NCBI數據庫比對后發現,共檢索到76條擬無枝酸菌的直系同源蛋白,與AMETH_3452的同源性均大于77.82%。聚類分析后(圖3)可知,Amycolatopsiseurytherma[13]、Amycolatopsissp.ATCC 39116、Amycolatopsismethan-olica、Amycolatopsisdeserti、Amycolatopsisgranulosa、Amycolatopsisviridis、Amycolatopsisendophytica[19]菌株在同一個分支,為AMS(甲基擬無枝酸菌分支)。另外有49個菌株聚類在AOS分支(東方擬無枝酸菌分支)上。而AOS又可以分成穩定的2個分支,自展值分別是83和99,可靠性較高。其中一支命名為AMEDS(地中海擬無枝酸菌分支),以A.mediterranei為代表,包括19個菌株;另一支為AORIS(東方擬無枝酸菌分支,屬于AOS的小分支),以A.orientalis為代表,共包括30個菌株。此外,從系統發育樹的根部可以看出,Amycolatopsisarida與其他菌株距離較遠。該菌株在2018年被Imen Nouioui從Yuhushielladeserti重新命名為A.arida[15]。此外從整體上看,多個種名未命名的菌株均聚類在進化樹中,與已有種名的菌株序列都有差異,同樣也說明該蛋白序列作為系統發育分析標記具有較強的分辨率和準確性。
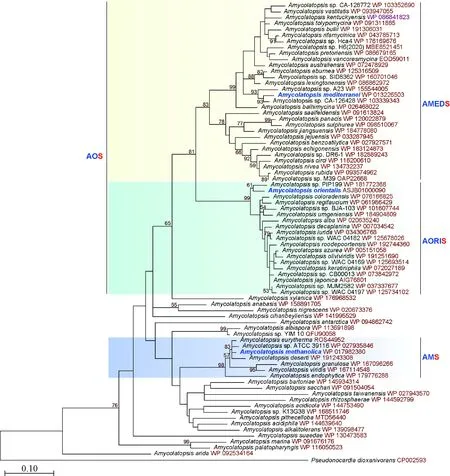
圖3 AMETH_3452蛋白序列在擬無枝酸菌中系統發育分析
3 討論
16S rDNA序列廣泛應用于細菌的系統發育分析,但由于部分細菌基因組中存在多個不同的16S rDNA序列拷貝,給系統發育樹分支結構造成了困擾[20-21]。本文利用一個廣泛存在于放線菌的具有269個氨基酸的保守蛋白AMETH_3452在擬無枝酸菌中進行了系統發育分析,得到了穩定的分支結構,與前人報道的系統發育樹具有一定相似性[5,6,14]。使用該蛋白序列在AOS中將菌株聚類,可明確將其分成2個分支AMEDS和AORIS。由于該蛋白序列的獲得主要依據NCBI數據庫中的全基因組序列,本文采用的序列相比16S rDNA的普適性具有一定劣勢,沒有窮盡該屬所有的菌株。
在擬無枝酸菌中,AMETH_3452基因序列保守性較高,下一步可以作為分子檢測的標記應用。此外,AMETH_3452蛋白具有一定的可溶性,未來可進一步研究其生理功能。鑒于該蛋白的保守性,下一步擬通過在放線菌其他目中進行更廣泛的系統發育分析,驗證該蛋白作為分子標記的適用性和效果。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經界(學術版)(2015年20期)2015-12-23 09:20:13
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
