長鏈非編碼RNA MAFG-AS1通過靶向miR-143-3p/AKT2軸促進胃癌的進展
2021-04-02 13:32:38陳軍芳岳新榮賀大春
腫瘤 2021年3期
陳軍芳,岳新榮,賀大春
胃癌(gastric cancer,GC)是一種常見的消化系統腫瘤,也是全球第4大最常見的惡行腫瘤[1]。近年來,盡管GC在手術治療、靶向治療和免疫治療方面取得了重大進展,由于大多數GC患者首次診斷位于晚期,治療方案有限,預后仍然很差[2]。由于GC的轉移復發率較高,全球GC患者的5年生存率不足20%[3]。因此,迫切需要為GC的早期診斷和治療尋找新的靶點。
長鏈非編碼RNA(long non-coding RNA,lncRNA)是一種長度超過200個核苷酸的非編碼RNA,近年來其在人類生理病理中的作用引起了越來越多的關注[4]。LncRNAs已被確定為癌癥發展過程的關鍵調節因子,并參與調控腫瘤細胞的增殖、侵襲、分化和凋亡等[5]。多種失調的lncRNAs,包括HOXC-AS3、ARAP1-AS1、TRPM2-AS和GCAT18均被證明與GC的發生和進展有關[6-9]。同屬于lncRNAs家族的lncRNA MAFG-AS1(后續簡稱為MAFG-AS1)已經被證實在多種癌癥組織中發揮促癌作用,如促進肝癌和乳腺癌的增殖和轉移[10-11]。重要的是,MAFG-AS1在GC中表達上調,并與患者不良預后相關[12]。然而MAFG-AS1在GC中的生物學功能和作用機制尚待研究,因此在本研究中將探究MAFG-AS1在GC中生物學功能和可能的作用機制。
本研究首次發現MAFG-AS1在GC組織中上調,并與GC患者的預后不良相關。基于此,進一步探討MAFG-AS1在GC組織中通過調控微(microRNA,miRNA,miR)-143-3p/AKT2(AK mouse plus Transforming or Thymoma 2)軸在GC中發揮的生物學作用及其機制,以期為GC患者的臨床治療提供一條新的思路。
1 材料與方法
1.1 臨床樣本
收集經本院醫院病理確診的39例GC患者的癌及其配對的癌旁正常組織(距原發灶邊緣3~5 cm以上的相應癌旁組織)。所有組織均在手術切除后,立即放入-80 ℃低溫冰箱中保存,用于提取總RNA。所有患者均未接受放療、化療或靶向藥物治療。本研究已獲得醫院倫理委員會批準,所有患者均已簽署知情同意書。
1.2 細胞、試劑及儀器
人胃黏膜正常上皮細胞GES-1和人GC細胞系BGC-823、SGC-7901和MGC-803細胞均購自美國模式菌種保藏中心(American Type Culture Collection,ATCC)。
RPMI 1640培養液購自美國Gibco公司。定點突變試劑盒購自上海玉博生物科技有限公司。熒光素酶報告基因載體pGL3-basic、攜帶有MAFG-AS1的重組過表達質粒及其對照空載體、miR-143-3p-模擬物(miR-143-3p-mimics)及其陰性對照-模擬物(NC-mimics)和特異性針對AKT2基因的siRNA(siAKT2)均購自美國Promega公司。TRIzol試劑和LipofectAMINETM2000試劑購自美國Invitrogen公司,反轉錄試劑盒和SYBR預混劑EXTAQⅡ購自日本TaKaRa公司。CCK-8試劑盒、青霉素、鏈霉素、胰蛋白酶、RIPA蛋白裂解液、BCA蛋白定量試劑盒和電化學顯影試劑盒均購自中國上海碧云天生物技術有限公司,Transwell小室購自美國Corning公司。兔抗人AKT2和GAPDH(內參照)單克隆抗體以及辣根過氧化物酶標記的羊抗兔IgG購自英國Abcam公司。ABI7500型熒光定量PCR儀為美國ABI公司產品。
1.3 生物信息學分析
采用GEPIA(Gene Expression Profiling Interactive Analysis)(http://gepia.cancer-pku.cn/)分析MAFG-AS1在GC組織中的表達情況;使用Kaplan-Meier Plotter數據庫(http://kmplot.com/analysis/index.php)分析MAFG-AS1的表達與GC預后的關系;采用StarBase數據庫(http://starbase.sysu.edu.cn)預測與MAFG-AS1結合的miRNA以及miR-143-3p下游的mRNA。
1.4 細胞培養
人胃黏膜上皮細胞GES-1細胞和人GC細胞系BGC-823、SGC-7901和MGC-803均用含10%胎牛血清和1%青霉素/鏈霉素的RPMI 1640培養液,置于37 ℃、CO2體積分數為5%的培養箱中進行培養。在細胞處于對數生長期時采用0.25%胰蛋白酶消化后傳代,以1×106個/孔的密度接種于6孔板上,待細胞生長穩定后進行細胞轉染。
1.5 細胞轉染
取處于對數生長期的MGC-803細胞,按照LipofectAMINETM2000轉染試劑說明書提供的流程,將空載體(對照組)和MAFG-AS1過表達重組質粒分別轉染MGC-803細胞,轉染48 h后采用實時熒光定量PCR法檢測轉染效率。
1.6 實時熒光定量PCR檢測
組織處理:收集GC組織及相應癌旁正常組織,稱取組織標本約100 mg置于EP管中,加入1 mL TRIzol試劑裂解組織,使用勻漿器充分研磨組織,在低溫下提取組織中的總RNA。細胞處理:收集大約1×106個轉染后的MGC-803細胞,用TRIzol試劑提取細胞總RNA。
采用TRIzol試劑常規提取組織和細胞的總RNA后,檢測其純度后,取部分RNA反轉錄為cDNA。在ABI 7500系統上,采用SYBR預混劑EXTAQⅡ進行實時熒光定量PCR擴增。以GADPH和U6作為內參照。各實驗組目的基因的表達量相對于對照組的表達量變化倍數用2-ΔΔCt法計算,引物序列見表1。
1.7 CCK-8法檢測細胞的增殖能力
將空載體(對照組)和MAFG-AS1過表達重組質粒(MAFG-AS1組)MGC-803細胞接種于96孔板中,每孔1×103個細胞。分別于培養24、48、72和96 h加入10 μL的CCK-8溶液,將培養板在培養箱內繼續孵育4 h,隨后在酶聯免疫酶標儀波長450 nm處檢測各孔細胞的D值。
1.8 EdU染色法檢測細胞的增殖能力
實驗分組同1.7節。取轉染后的MGC-803細胞分別接種于24孔板中,待細胞生長至對數生長期時,每孔加入200 μL濃度為50 μmol/L的EdU染色液,于細胞培養箱內孵育2 h,PBS清洗。隨后,采用4%多聚甲醛溶液固定細胞,室溫孵育10 min。加入200 μL濃度為2 mg/mL的甘氨酸孵育5 min后,搖床上PBS清洗5 min。每孔中加入100 μL含0.5%的TritonX-100的PBS溶液,搖床上脫色孵育10 min,PBS洗滌2次,每次5 min。加入Apollo室溫下避光染色30 min,再用DAPI染色液室溫避光孵育20 min。PBS清洗后在熒光顯微鏡下拍照計數,分析結果。藍色熒光標記細胞總數,綠色熒光標記增殖的細胞,細胞增殖率(%)=綠色熒光細胞數/藍色熒光細胞數×100%。
1.9 Transwell小室實驗
實驗分組同1.7節。取對數生長期的MGC-803細胞,用胰蛋白酶消化后,用含1%胎牛血清的培養液重懸細胞且計數,單細胞懸液的密度為2×106個/mL,取200 μL細胞懸液接種于小室的上室中,小室的下室中則加入600 μL完全培養液。細胞接種24 h后,將小室直接浸泡于甲醇溶液中室溫固定 20 min。待細胞固定后,吸除小室上室中的培養液,用棉簽小心擦去剩余培養液和未穿過小室膜的細胞。將小室放入含1%結晶紫的染色液中,室溫染色10 min,流動水沖洗小室,晾干水分,在光學顯微鏡下(放大倍數為200倍)觀察細胞形態并計數,拍照保存檢測結果。

表1 實時熒光定量PCR引物序列Table 1 The primer sequence of real-time fluorescent quantitative PCR
1.10 雙熒光素酶報告基因檢測
采用定點突變試劑盒對預測的結合位點進行突變。將MAFG-AS1和AKT2 mRNA 3’-非翻譯區(3’-untranslated region,3’-UTR)中含有預測miRNA結合位點的序列片段插入到熒光素酶報告基因載體pGL3-basic中,從而分別構建野生型(wild type,WT)和突變型(mutant type,MUT)熒光素酶報告質粒(MAFG-AS1-WT、MAFG-AS1-MUT、AKT2-WT和AKT2-MUT)。
將MGC-803細胞以1×105個/孔的密度接種于24孔板中,培養至融合度為70%時,收集細胞。采用轉染試劑LipofectAMINETM2000將MAFGAS1-WT、MAFG-AS1-MUT與miR-143-3p-mimics或NC-mimics兩兩組合共轉染MGC-803細胞,驗證MAFG-AS1和miR-143-3p的相關性;將AKT2-WT、AKT2-MUT和miR-143-3p-mimics或NCmimics兩兩組合共轉染MGC-803細胞,驗證miR-143-3p與AKT2的相關性。轉染48 h后,按照試劑盒操作說明檢測各組細胞的熒光素酶活性。
1.11 蛋白質印跡法檢測MAFG-AS1過表達后上調miR-143-3p表達或下調AKT2表達對MGC-803細胞中AKT2蛋白表達水平的影響
實驗分為4組:空載體組(對照組)、MAFG-AS1組、MAFG-AS1+miR-143-3p組(共轉染MAFG-AS1過表達重組質粒和miR-143-3p-mimics)和MAFG-AS1+siAKT2組(共轉染MAFG-AS1過表達重組質粒+siAKT2)。采用RIPA裂解MGC-803細胞,提取細胞中的總蛋白。而后采用BCA法檢測蛋白濃度。取適量蛋白行10% SDS-PAGE分離蛋白,電泳結束后將分離后的蛋白轉移至PVDF膜上;用含5%脫脂奶粉的封閉液封閉處理1 h,然后加入一抗[兔抗人AKT2單克隆抗體(體積稀釋比例為1∶500)]在4 ℃下孵育過夜;用TBST洗膜后,加入二抗[辣根過氧化物酶標記的羊抗兔IgG(體積稀釋比例為1∶1 000)]室溫孵育1 h。加入電化學發光液發光顯影,在凝膠成像系統中進行圖像采集,使用Image J分析蛋白質條帶。
1.12 功能補償實驗
實驗分為4組同1.11節。采用CCK-8法、EdU實驗和Transwell小室實驗檢測MAFGAS1、miR-143-3p和AKT2對細胞增殖和遷移的影響。
1.13 統計學方法
采用SPSS 22.0統計軟件和Graphpad Prism 8對實驗數據進行分析。計量資料數據以表示,兩組間比較采用t檢驗,多組間比較采用方差分析。計數資料數據采用四格表(或百分率)表示,由于N<40,采用確切概率法進行差異分析。以P<0.05為差異有統計學意義。
2 結果
2.1 MAFG-AS1在GC細胞和組織中過表達且與GC患者預后不良相關
為了探究MAFG-AS1在GC細胞中的表達水平,首先通過GEPIA數據庫分析發現,MAFG-AS1在GC組織中高表達(圖1A),差異均有統計學意義(P<0.05)。
隨后,通過實時熒光定量PCR檢測39例GC患者癌及癌旁組織中,以及人胃黏膜上皮細胞GES-1細胞和人GC細胞系BGC-823、SGC-7901和MGC-803細胞中MAFG-AS1的表達水平。結果顯示,與癌旁組織相比,MAFGAS1在GC組織的表達水平明顯上調(P值均<0.001)(圖1B);與人胃黏膜上皮細胞GES-1細胞相比,人GC細胞系BGC-823、SGC-7901和MGC-803細胞中的表達水平均明顯上調(P值均<0.001)(圖1C)。MAFG-AS1高表達與GC患者是否淋巴結轉移(P=0.010 4)和分化程度低(P=0.0150)具有相關性(表2)。
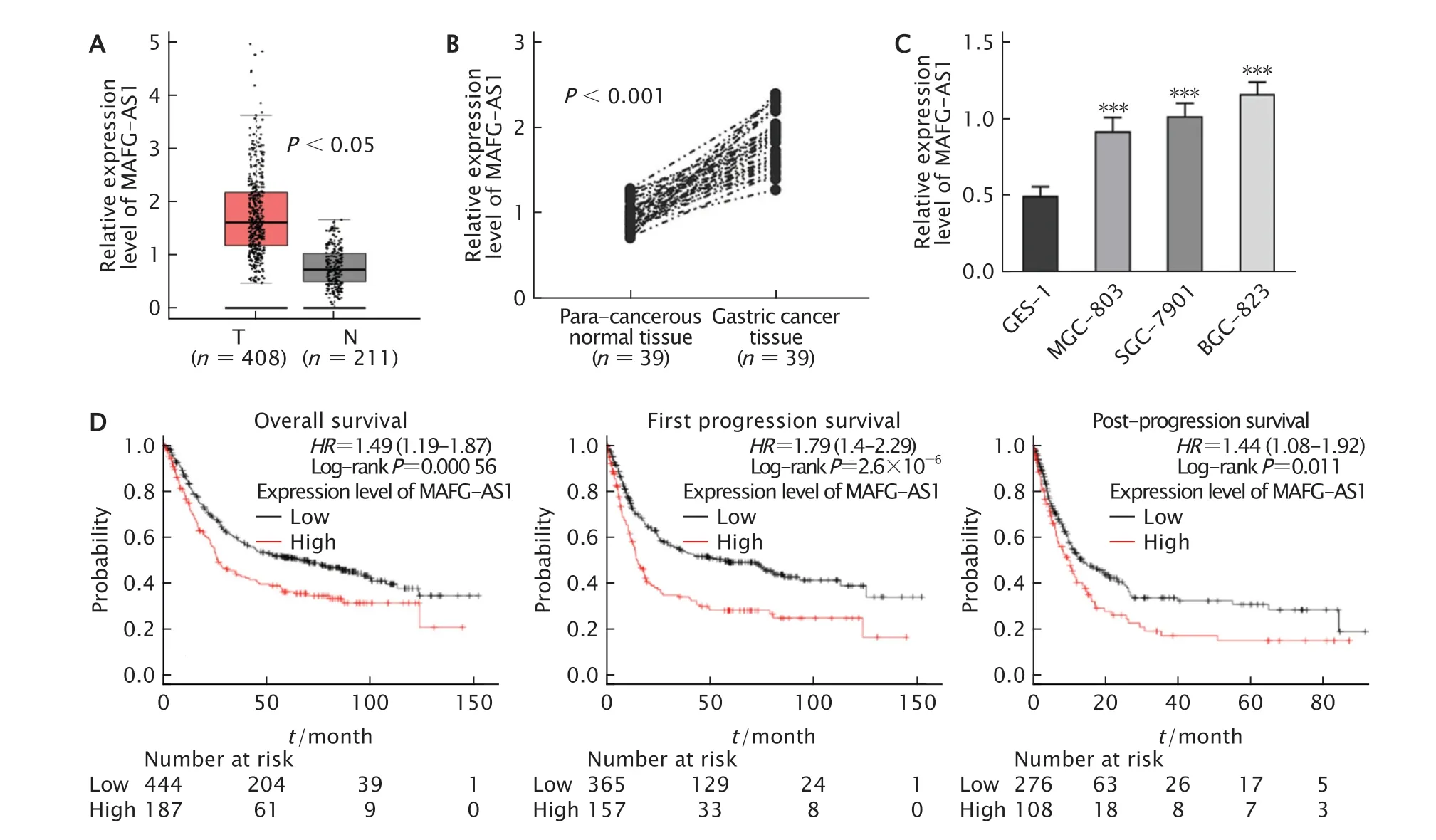
Fig.1 Relationship between the expression of long non-coding RNA (lncRNA) MAFG-AS1 and prognosis of gastric cancer(GC) patients.A:MAFG-AS1 expression from Gene Expression Profiling Interactive Analysis (GEPIA);B-C:MAFGAS1 expression in GC tissues and cells (BGC-823,SGC-7901 and MGC-803 cells,normal gastric epithelial GES-1 cells as the control) was detected by real-time fluorescent quantitative PCR;D:The expression level of MAFG-AS1 was negatively correlated with the overall survival,first progression survival and post-progression survival.T:Tumor tissue;N:Para-cancerous normal tissue;HR:Hazard ratio;**P<0.01,***P<0.001,vs GES-1 cells (n=3).圖1 LncRNA MAFG-AS1在GC組織和細胞中的表達水平及與GC患者生存期的關系
通過Kaplan-Meier Plotter數據庫分析發現,MAFG-AS1高表達與GC患者的總體生存期(overall survival,OS)和首次進展生存期(first progression survival)以及后進展生存期(postprogression survival)呈負相關(圖1D),差異均有統計學意義(P=0.000 56,P=2.6×10-6和P=0.011)。由此可見,MAFG-AS1的表達失調與GC患者預后不良密切相關。
2.2 過表達MAFG-AS1顯著促進GC細胞的增殖和遷移
為了驗證MAFG-AS1在GC中的作用,將MAFG-AS1過表達質粒轉入MGC-803細胞中,隨后采用實時熒光定量PCR法檢測MAFG-AS1表達水平。結果(圖2A)顯示,空載體組(對照組)和MAFG-AS1組中MAFG-AS1的表達量分別為1.0±0.14和3.1±0.33,相較于對照組,MAFG-AS1組中MAFG-AS1的表達水平明顯上調(P<0.001)。
CCK-8法檢測結果(圖2B)顯示,與對照組相比,MAFG-AS1組的細胞活力明顯升高,從48 h時開始,差異具有統計學意義(P值均<0.05)。
EdU實驗檢測結果(圖2C)顯示,對照組和MAFG-AS1組細胞EdU染色陽性細胞所占百分比分別為(29.2±3.3)%和(69.5±7.3)%,MAFG-AS1組細胞的陽性率明顯高于對照組(P<0.001)。
Transweell小室實驗檢測結果(圖2D)顯示,對照組和MAFG-AS1組發生遷移的細胞數分別為(62.4±6.5)個和(152.6±17.3)個,MAFG-AS1組發生細胞的遷移數明顯多于對照組(P<0.01)。
2.3 miR-143-3p是MAFG-AS1的下游靶標
通過StarBase數據庫預測了MAFG-AS1與miR-143-3p存在結合位點(圖3A)。采用實時熒光定量PCR法檢測癌及癌旁組織中miR-143-3p的表達水平,結果(圖3B)顯示,與癌旁組織相比,miR-143-3p表達水平在GC組織中明顯下調(P<0.001)。同時,Pearson相關性分析結果(圖3C)表明,MAFG-AS1與miR-143-3p的表達呈負相關(R2=0.144 8,P<0.05)。
通過雙熒光素酶報告基因系統進一步探究MAFG-AS1與miR-143-3p的關系。結果(圖3D)顯示,上調miR-143-3p表達可明顯抑制MAFG-AS1-WT的熒光素酶活性(P<0.001),而對MAFG-AS1-MUT的熒光素酶活性無明顯影響(P>0.05)。隨后,采用實時熒光定量PCR法檢測發現(圖3E),過表達MAFG-AS1可抑制miR-143-3p的表達水平(P<0.01),而上調miR-143-3p表達水平對MAFG-AS1的表達無明顯影響(P>0.05)。由此可見,GC中MAFG-AS1直接靶向miR-143-3p,并負調節其的表達。
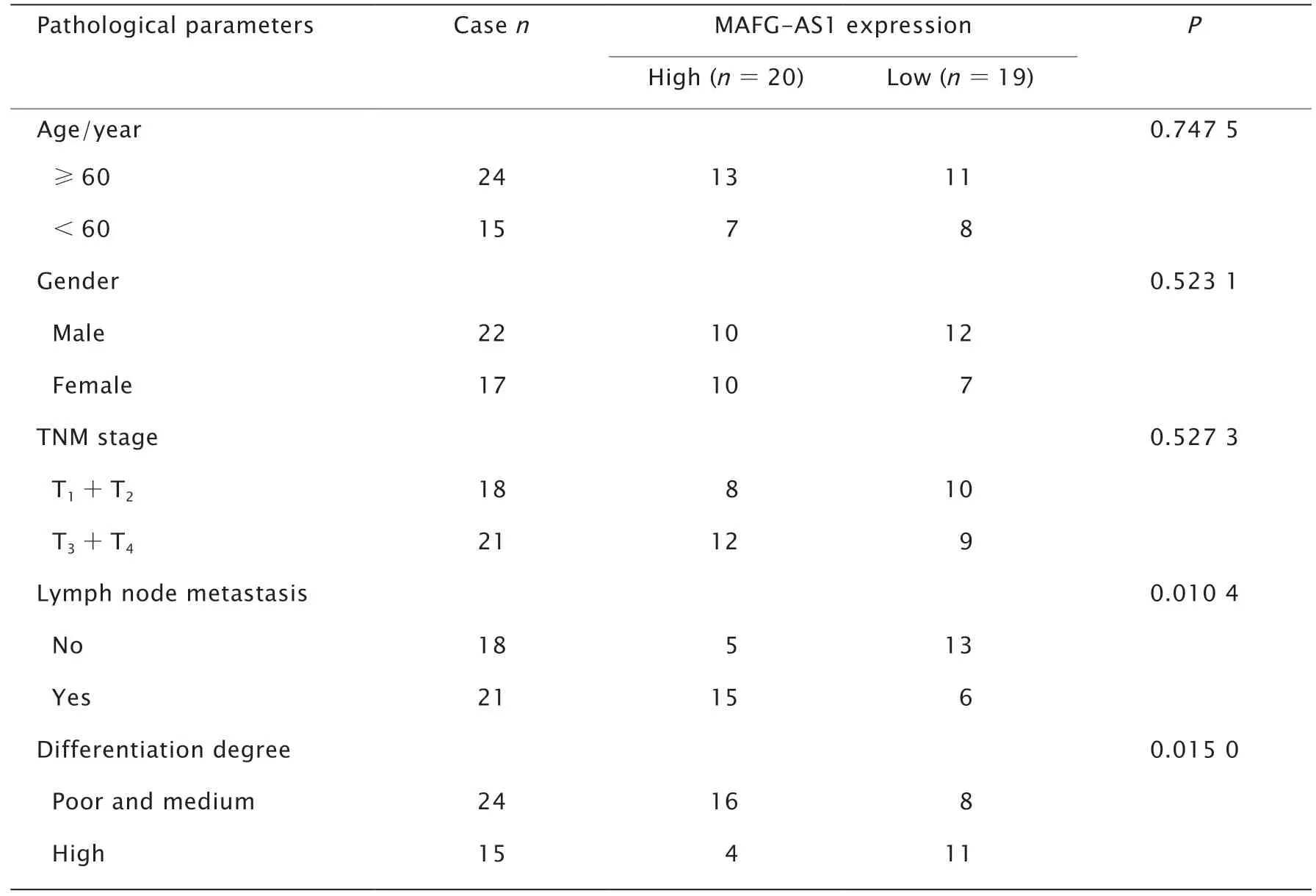
表2 LncRNA MAFG-AS1表達與GC患者臨床病理特征的關系Table 2 Correlation between clinicopathological features and expression of long non-coding RNA(lncRNA) MAFG-AS1 in 39 patients with gastric cancer (GC)N=39,n
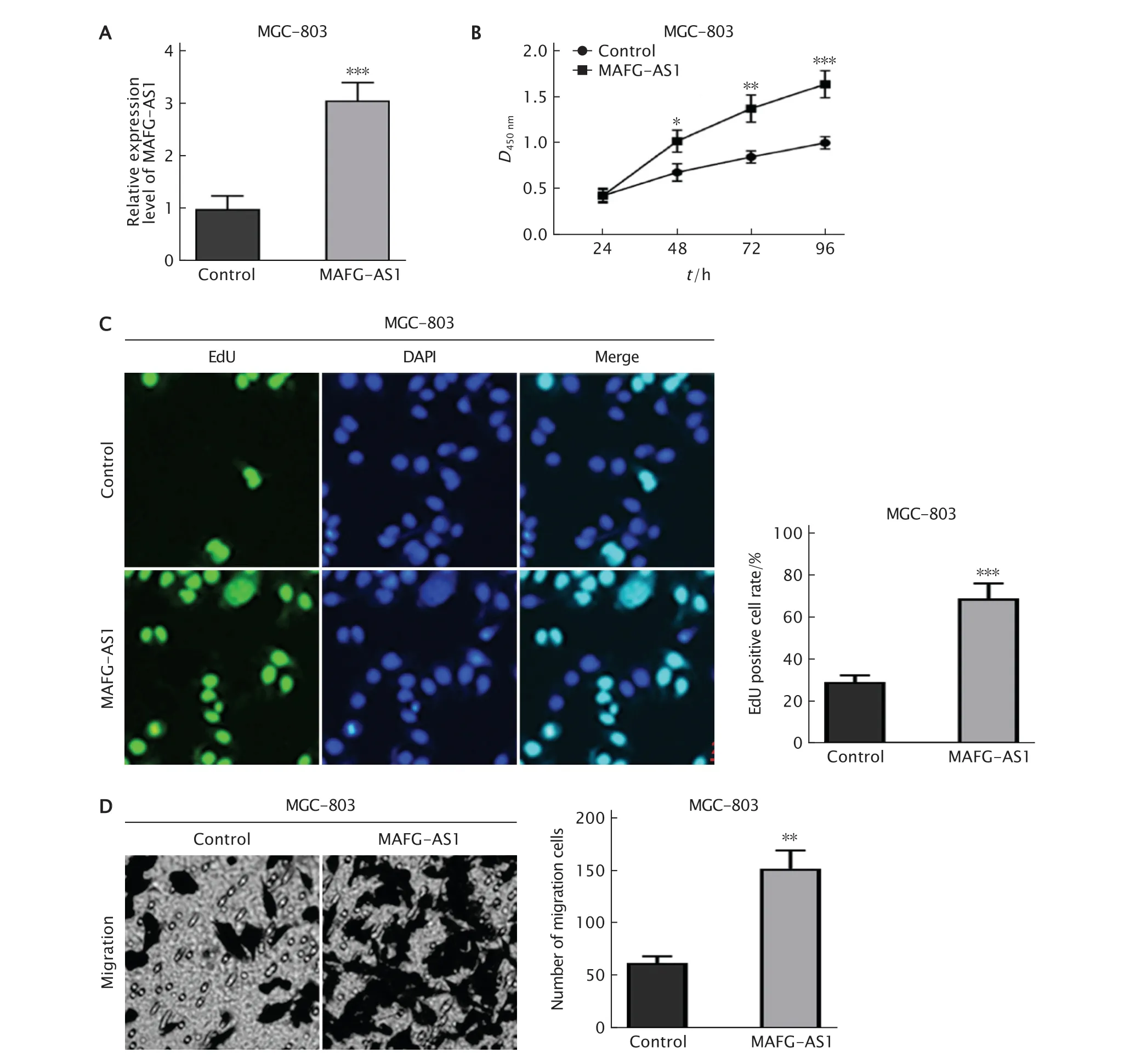
Fig.2 Effects of long non-coding RNA (lncRNA) MAFG-AS1 on proliferation and migration of gastric cancer (GC)MGC-803 cells.A:The transfection efficiency was detected by real-time fluorescent quantitative PCR;B-C:The efffect of MAFG-AS1 overexpression on the proliferation of MGC-803 cells was detected by CCK-8 assay and EdU staining,respectively;D:Transwell assay was conducted to evaluate the effect of MAFG-AS1 overexpression on MGC-803 cells migration.MAFG-AS1 group:MGC-803 cells were transfected with recombinant plasmid expressing MAFG-AS1 by liposome.MGC-803 cells were transfected with empty vector by liposome as the control;*P<0.05,**P<0.01,***P<0.001,vs the control group (n=3).圖2 過表達MAFG-AS1對MGC-803細胞增殖和遷移的影響
2.4 LncRNA MAFG-AS1通過靶向結合miR-143-3p上調AKT2的表達
通過分析StarBase數據庫發現,AKT2與miR-143-3p存在結合位點(圖4A)。隨后,通過雙熒光素酶報告基因實驗檢測發現(圖4B),轉染miR-143-3p-mimics可抑制AKT2-WT組的熒光素酶活性(P<0.01),而對AKT2-MUT組的熒光素酶活性無明顯影響(P>0.05)。
蛋白質印跡法檢測結果(圖4C)顯示,過表達MAFG-AS1可上調AKT2蛋白的表達水平,而上調miR-143-3p表達或敲低AKT2表達均可抑制AKT2蛋白的表達水平,差異均有統計學意義(P值均<0.001)。
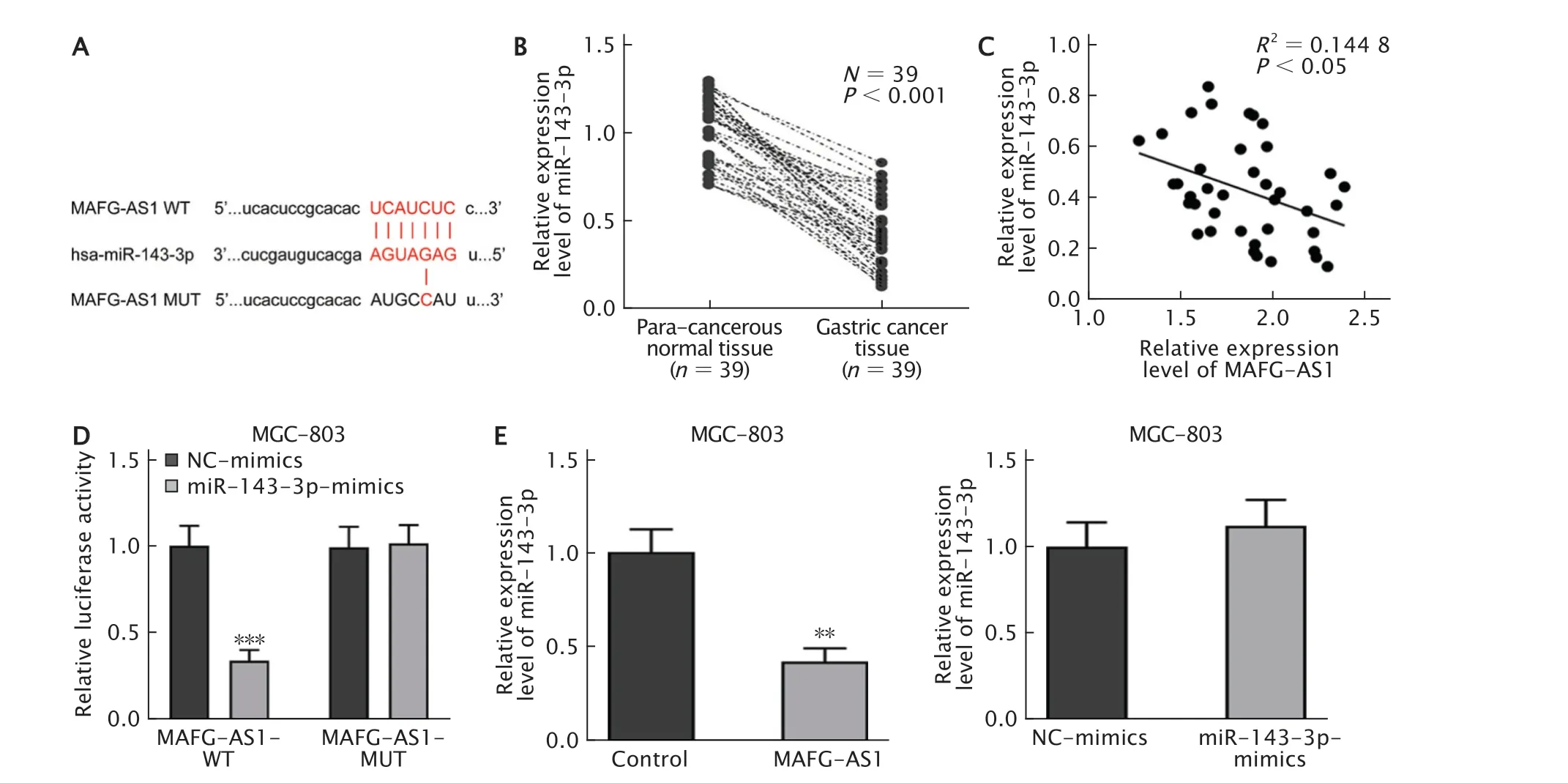
Fig.3 Long non-coding RNA (lncRNA) MAFG-AS1 directly targets miR-143-3p.A:The binding site between MAFGAS1 and miR-143-3p was predicted through the StarBase database;B:The miR-143-3p expression level in gastric cancer(GC) and para-tumor tissues was detected by real-time fluorescent quantitative PCR;C:Pearson correlation analysis was conducted to evaluate the relevance between the expressions of MAFG-AS1 and miR-143-3p.D:Dual-luciferase reporter assay was conducted to detect the relationship between MAFG-AS1 and miR-143-3p;E:MAFG-AS1 and miR-143-3p expression levels in MGC-803 cells were detected by real-time fluorescent quantitative PCR.**P<0.01,***P<0.001 (n=3).圖3 分別采用雙熒光素酶報告基因系統和實時熒光定量PCR法驗證MAFG-AS1和miR-143-3p的靶向關系
采用實時熒光定量PCR檢測癌及其癌旁組織中MAFG-AS1、miR-143-3p和AKT2 mRNA的表達水平,再用Pearson進行相關性分析。結果顯示,miR-143-3p與AKT2 mRNA的表達水平呈負相關(R2=0.364 0,P<0.001)(圖4D),MAFG-AS1與AKT2 mRNA的表達水平呈正相關(R2=0.223 1,P<0.05)(圖4E)。此外通過對StarBase數據庫分析發現(圖4F),AKT2與MAFG-AS1的表達呈正相關(r=0.229,P=7.55×10-6)。由此推測,GC中MAFGAS1通過抑制miR-143-3p表達上調的AKT2的表達水平。
2.5 MAFG-AS1通過調控miR-143-3p/AKT2軸促進MGC-803細胞的增殖和遷移
為了進一步驗證MAFG-AS1是否通過調控miR-143-3p/AKT2軸對MGC-803細胞的增殖和遷移產生影響,分別將空載體(對照組)、MAFG-AS1過表達重組質粒(MAFG-AS1組)、MAFG-AS1過表達重組質粒+miR-143-3p-mimics(MAFG-AS1+miR-143-3p組)和MAFG-AS1過表達重組質粒+siAKT2(MAFGAS1+siAKT2組)分別轉入MGC-803細胞中。
實時熒光定量PCR法檢測結果(圖5A)顯示,與對照組相比,MAFG-AS1過表達后miR-143-3p的表達水平明顯下調(P<0.001);相較與MAFG-AS1組,MAFG-AS1+miR-143-3p組中miR-143-3p的表達水平明顯上調(P<0.001),而MAFG-AS1+siAKT2組中miR-143-3p的表達水平沒有明顯變化(P>0.05)。
CCK-8法檢測結果(圖5B)顯示,與對照組相比,MAFG-AS1組細胞的活力明顯提高(P<0.001),而MAFG-AS1+miR-143-3p組和MAFG-AS1+siAKT2組的細胞活力均低于MAFG-AS1組(P<0.01和P<0.001)。
EdU實驗檢測結果(圖5C)顯示,對照組和MAFG-AS1組EdU染色陽性細胞所占的百分比分別為(31.5±3.5)%和(71.2±7.6)%,MAFG-AS1組細胞的增殖活性較對照明顯提高(P<0.001);MAFG-AS1+miR-143-3p組和MAFG-AS1+siAKT2組細胞的EdU染色陽性細胞所占的百分比分別為(37.5±3.9)%和(33.4±3.6)%,均低于MAFG-AS1組(P值均<0.001)。
Transweell小室遷移實驗檢測結果(圖5D)顯示,對照組和MAFG-AS1組發生細胞的遷移數分別為(57.6±6.3)個和(141.2±14.5)個,MAFG-AS1組發生遷移的細胞數明顯多于對照組(P<0.001);MAFG-AS1+miR-143-3p組和MAFG-AS1+siAKT2組發生遷移的細胞數分別為(64.3±6.7)個和(61.7±6.3)個,均低于MAFG-AS1組(P值均<0.001)。
上述結果表明,MAFG-AS1可能通過抑制miR-143-3p表達促進GC細胞的增殖和遷移。
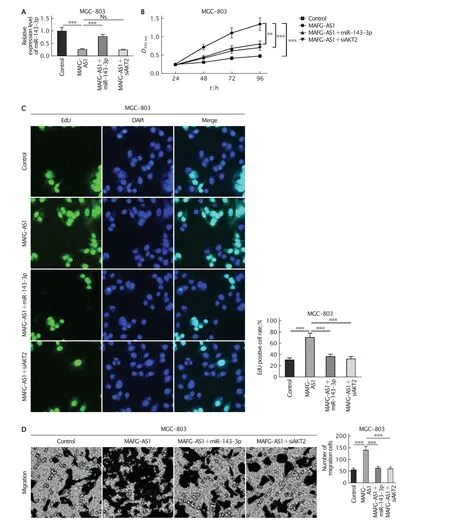
Fig.5 The effects of long non-coding RNA (lncRNA) MAFG-AS1 on proliferation and migration of MGC-803 cells by regulating miR-143-3p/AKT2 axis.A:The transfection efficiency was detected by real-time fluorescent quantitative PCR;B-C:The effect of MAFG-AS1 on MGC-803 cells proliferation was detected by CCK-8 assay and EdU staining,respectively;D:Transwell assay was used to detect the effect of MAFG-AS1 on MGC-803 cells migration.**P<0.01,***P<0.001 (n=3).圖5 MAFG-AS1可能通過抑制miR-143-3p表達促進MGC-803細胞的增殖和遷移
3 討論
LncRNA可分為基因內lncRNA、基因間lncRNA、正義lncRNA及反義lncRNA等多種類型,在多個層面調控基因的表達,如調節其附近蛋白編碼基因的表達、參與調節其他RNA的加工、作為內源競爭性RNA與miRNA結合進而調控靶基因的表達等[13]。近年來,越來越多的研究表明lncRNAs在包括GC在內的多種腫瘤的生長和轉移中發揮關鍵調節作用[14-15]。有研究報道,MAFG-AS1通過抑制miR-147b的表達而上調NDUFA4的表達水平,進而促進結直腸癌的進展[16]。MAFG-AS1通過調節miR-744-5p/MAFG軸促進肺腺癌細胞增殖[17]。而在乳腺癌中,MAFG-AS1通過調節miR-339-5p/基質金屬蛋白酶15(matrix metalloproteinase 15,MMP15)促進腫瘤細胞的侵襲性[18]。GC中MAFG-AS1表達上調,其高表達水平與腫瘤臨床分期提高,浸潤深度加深和患者生存期短密切相關;敲低MAFGAS1表達可抑制GC細胞的增殖、遷移和侵襲[12],這一結果與本研究結果一致。本研究中,通過分析GEPIA數據庫和Kaplan-Meier Plotter數據庫發現,MAFG-AS1在GC中表達失調,并與患者不良預后相關;進一步研究表明,過表達MAFG-AS1可促進GC細胞的增殖和遷移,這表明MAFG-AS1在GC中發揮促癌作用。
MiRNAs是一種在許多生物學進程中調節基因表達的非編碼RNA,miRNAs通過調節靶基因的表達和下游重要信號轉導通路的活性,參與調控癌細胞的增殖、遷移和侵襲、分化和凋亡[19-20]。有研究表明,miR-143-3p可通過靶向結合mRNA的3’-UTR在腫瘤中發揮抑制作用,如miR-143-3p通過靶向結合FOSL2(Foslike antigen 2)抑制骨肉瘤的增殖、遷移和侵襲[19]。類似研究表明,miR-143-3p通過靶向結合K-RAS抑制胰腺導管腺癌的發生[20]。乳腺癌中,miR-143-3p通過靶向絲裂原活化蛋白激酶7(mitogen-activate protein kinase 7,MAPK7)抑制腫瘤的增殖和轉移[21]。而在GC中miR-143-3p表達下調,其表達失調與患者是否淋巴結轉移密切相關,功能上miR-143-3p通過靶向結合AKT2抑制GC細胞的增殖和轉移[22]。本研究中同樣發現,miR-143-3p在GC組織中表達下調,隨后通過驗證發現miR-143-3p通過靶向AKT2在GC細胞的增殖和遷移中發揮抑制作用。
AKT2屬于AKT家族,是一種常見致癌基因,其在調節血管生成、腫瘤生長、細胞轉移和化學抗性中發揮重要作用[23]。有研究提示,AKT2可促進卵巢癌中丙酮酸激酶的表達,從而誘導體外細胞的遷移和侵襲,以及體內腫瘤細胞轉移[24]。在乳腺癌中,AKT2通過調節F-肌動蛋白和波形蛋白的表達,從而促進腫瘤的生長、浸潤和肺轉移[25]。而在GC中,AKT2表達上調,敲低AKT2表達可抑制GC細胞的生長和轉移,促進其凋亡,且AKT2的表達受到miR-143-3p的靶向調控[22]。本研究中,再次驗證提示miR-143-3p負調控AKT2的表達,MAFG-AS1正向調控的AKT2水平,這表明MAFG-AS1通過調控miR-143-3p/AKT2調控軸在GC進展中發揮作用。
總之,本研究中發現MAFG-AS1在GC中的表達水平上調,并預示著GC患者的不良預后。進一步研究表明,MAFG-AS1可促進GC細胞的增殖和遷移。相關機制研究表明,MAFGAS1可通過調控miR-143-3p/AKT2軸在GC的進展中發揮作用。本研究結果為GC發病機制提供了新的見解,將有助于GC的早期診斷和治療。然而,本研究仍有許多不足之處,研究局限于單中心樣本,將來需要更大的樣本來證實這一結論。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
