白蟻菌圃中產纖維素酶菌株的鑒定與酶活研究
2021-04-20 06:00:47韋張其王潔慧劉國慶
安徽農業科學 2021年6期
韋張其 王潔慧 劉國慶
摘要 [目的]提高生物質秸稈降解速度,能夠更加充分地利用微生物資源。[方法]以黑翅土白蟻菌圃為菌源,采用平板稀釋法初篩分離得到降解纖維素的目標菌株,再復篩得到最優菌株FUN-ce4,然后對其進行ITS序列分析鑒定以及酶穩定性研究。[結果]菌株FUN-4歸屬于栓孔菌屬(Trametes)。經對酶活性研究,菌株FUN-4在溫度為60 ℃、pH為4、發酵時間為6 d時酶活相對較高;而溫度為40 ℃、最適pH為5時酶活相對較穩定。[結論]白蟻菌圃中存在活性較高的纖維素降解菌株,可以成為高活性木質纖維素降解酶的重要來源。
關鍵詞 白蟻菌圃;纖維素酶;分離;鑒定;酶活
Abstract [Objective]To improve the degradation rate of biomass straw and make full use of microbial resources.[Method]The target strains degrading cellulose from termite combsare was isolated by plate dilution method,and then the optimal strain FN-4 was rescreened from the target strains,and ITS sequence analysis was conducted combining with the research of the enzymatic activity.[Result]The strain FN-4 was identified as Trametes.The enzymatic activity research experiment showed that the enzymatic activity of strain FUN-4 was relatively high when the temperature was 60 ℃,the pH was 4,and the fermentation time was 6 days.When the temperature was 40 ℃ and the pH was 5,the enzyme activity was relatively stable.[Conclusion]There are high activity cellulose degrading strains in termites fungus gardens,which can be an important source of high activity lignocellulosic degrading enzymes.
Key words Termite combsare;Cellulase;Screening;Identification;Enzymatic activity
白蟻生物系統顯示了高效生物轉化木質纖維素的能力[1]。白蟻之所以能食木是因為它能靠其體內的微生物群將木材中纖維素和半纖維素消化成能夠被機體吸收的物質[2]。在白蟻的蟻巢中存在“菌圃”這一特殊的結構,其主要組成成分是未被完全消化的植物,白蟻和菌圃真菌之間較好的協同作用能夠高效利用木質纖維素[3]。
目前,關于白蟻共生微生物降解木質纖維素研究主要集中在白蟻腸道菌功能,并分離出其中的產酶基因進行基因組分析[4-8],而關于菌圃真菌降解纖維素的研究較少。該研究以黑翅土白蟻菌圃為研究對象,篩選出高產纖維素酶真菌并研究其纖維素酶活性及穩定性,從而發掘具有應用價值的纖維素降解真菌。
1 材料與方法
1.1 材料
1.1.1 供試菌株。黑翅土白蟻及其菌圃,購于江西省白蟻活體實驗室。
1.1.2 培養基。
①羧甲基纖維素-剛果紅培養基:CMC-Na 2.0 g,(NH4)2SO4 2.0 g,MgSO4·7H2O 0.5 g,K2HPO4 1.0 g,NaCl 0.5 g,剛果紅 0.4 g,瓊脂 20.0 g,蒸餾水1 000 mL,pH自然。②PDA培養基:200 g馬鈴薯,葡萄糖2 g,瓊脂2 g。③發酵產酶培養基:秸稈粉 5.0 g,蛋白胨 2.0 g,KH2PO4 2.0 g,FeSO4·7H2O 0.4 g,NaCl 0.5 g,蒸餾水1 000 mL,pH自然。各培養基按配方配制后,加熱攪勻,分裝至錐形瓶中,加塞、包扎,121 ℃滅菌20 min左右后取出錐形瓶搖勻,冷卻后貯存備用。
1.1.3 主要試劑及設備。
試劑:剛果紅染色液、氯化鈉脫色液、檸檬酸-磷酸氫二鈉緩沖液、DNS試劑等。
主要設備:分析天平、高速離心機、立式壓力蒸汽滅菌鍋、紫外可見光分光光度計、電熱恒溫培養箱等。
1.2 方法
1.2.1 菌圃中降解纖維素的菌株篩選。
取0.5 g菌圃碾碎,放入50 mL無菌離心管中,加入適量無菌水,于8 000 r/min的轉速下離心15 min。取 1 mL 靜置后的上清液分別稀釋至10-1、10-2、10-3、10-4和10-5倍,涂抹至羧甲基纖維素-剛果紅培養基平板上,30 ℃培養7 d。選取生長良好且透明水解圈較大的菌株接種于PDA培養基上28 ℃培養5 d,至在每個平板上都可看到單一形態的菌落。選取單菌落打孔接種于羧甲基纖維素培養基中,30 ℃培養3 d后,量取1 mg/mL的剛果紅染液15 mL染色15 min,用1 mol/L NaCl溶液脫色后,根據透明圈的大小,選出酶活最大的菌株(暫命名為FUN-4)進行分子鑒定和酶活性分析。
1.2.2 菌圃中降解纖維素的菌株鑒定。
1.2.2.1 形體觀察。
將純化菌株用黏片法,粘取菌絲于載玻片上,用顯微鏡和電鏡觀察其形態特征,鑒定參照文獻[9-10]提供的方法進行。
1.2.2.2 ITS序列分析鑒定[11]。FUN-ce4菌株基因組DNA采用E.Z.N.A.Stool DNA Kit試劑盒提取。
PCR反應體系:PhusionHot Start Flex 2× Master Mixart Version 12.5 μL、Forward Primer(1 μmol/L)2.5 μL、Reverse Primer(1 μmol/L)2.5 μL、模板DNA 50 μL、加ddH2O 25 μL。
PCR反應程序:預變性98 ℃ 30 s;變性98 ℃10 s,退火54 ℃ 30 s,延伸72 ℃ 45 s,35個循環;最后延伸72 ℃ 10 min;保持4 ℃。
PCR擴增產物通過2%瓊脂糖凝膠電泳進行檢測,并對目標片段進行回收,回收采用AMPure XT beads回收試劑盒。將PCR產物送至杭州聯川生物測序。將測序結果提交GenBank數據庫進行Blast,從GenBank數據庫中搜索相關菌株的ITS相應序列,采用MEGA 5.0鄰接法建進化樹,進行分離菌株菌種的鑒定。
1.2.3 菌株纖維素酶活研究。
1.2.3.1 粗酶液的制備。
將FUN-4菌株接種于液體PDA培養基中,30 ℃搖床180 r/min培養3 d。按10%接種量接入裝有100 mL發酵產酶培養基的250 mL三角瓶中,30 ℃搖床180 r/min培養。分別在2、3、4、5、6、7 d取粗酶液測定纖維素酶活。纖維素酶是一組酶的總稱,是一種復合酶,主要由外切β-葡聚糖酶、內切β-葡聚糖酶和β-葡萄糖苷酶等組成[12-13],因此纖維素酶活分別由微晶纖維素酶活、羧甲基纖維素酶活和濾紙條酶活反映。
1.2.3.2 纖維素酶活的測定。
(1)羧甲基纖維素酶活力[14]。取100 μL 粗酶液加入1% CMC溶液(溶于pH為4.5的檸檬酸-磷酸氫二鈉緩沖液),50 ℃條件下反應30 min(以100 ℃條件下反應30 min的酶液作為滅活對照組),然后加入1 mL DNS試劑充分搖勻并沸水煮5 min,冷水冷卻后在OD540下測定吸光度。
(2)濾紙酶活力[15]。取500 μL粗酶液加入1 500 μL緩沖液和1 cm×6 cm的濾紙(需卷成),在恒溫水浴鍋中預熱5 min,50 ℃條件下反應60 min(以100 ℃條件下反應60 min的酶液作為滅活對照組),然后加入2 mL DNS試劑充分搖勻并沸水煮5 min,冷水冷卻后在OD540下測定吸光度。
(3)微晶纖維素酶(Avicel)活力。取100 μL粗酶液加入400 μL緩沖液和1%微晶纖維素在恒溫水浴鍋中預熱5 min,50 ℃條件下反應60 min(以100 ℃條件下反應60 min的酶液作為滅活對照組),然后加入1 mL DNS試劑,充分搖勻并沸水煮5 min,冷水冷卻后在OD540下測定吸光度。
酶活力單位定義[16]:50 ℃恒溫條件下,以水解反應中1 min 催化底物水解形成1 μmol還原糖的酶量為1個酶活單位(U)。
1.2.4 FUN-ce4菌株產纖維素酶穩定性研究。
在不同pH緩沖液體系和不同溫度下測定酶活力和殘余活力,分析其對酶促反應和酶穩定性的影響,以達到最佳產酶條件。
2 結果與分析
2.1 菌圃中降解纖維素的菌株篩選
2.1.1 純化。
從5個梯度的羧甲基纖維素-剛果紅培養基中選取6個生長良好且具有較大透明圈的菌株進行純化,得到4株菌株,分別暫命名為FUN-1、FUN-2、FUN-3、FUN-4(圖1)。
觀察到這些菌落均生長較快且菌落較大,質地疏松,呈棉絮狀和絨毛狀,不透明,顏色有純白、乳白、黃色、綠色。菌落與培養基連接比較緊密,不易挑取。
2.1.2 復篩。
挑取純化后的菌株單菌落接種于羧甲基纖維素培養基,經剛果紅染液染色后NaCl溶液脫色,得到FUN-1、FUN-2、FUN-3、FUN-4各菌株的剛果紅水解圈(圖2)。
測量各菌株的菌落直徑和透明圈直徑,比較菌株活性(表1)。通過分離純化篩選最終得到酶活較強的菌種FUN-ce4,并進行分子學鑒定和酶穩定性分析。
2.2 菌種的分類鑒定
2.2.1 菌株形態鑒定。鏡檢圖中可以看到有比較粗壯的菌絲,許多偏透明的孢子,長長的分生孢子梗頭部有一個頂囊(圖3),菌落形態和鏡檢結果可以初步確定為霉菌。
2.2.2 ITS分子鑒定。
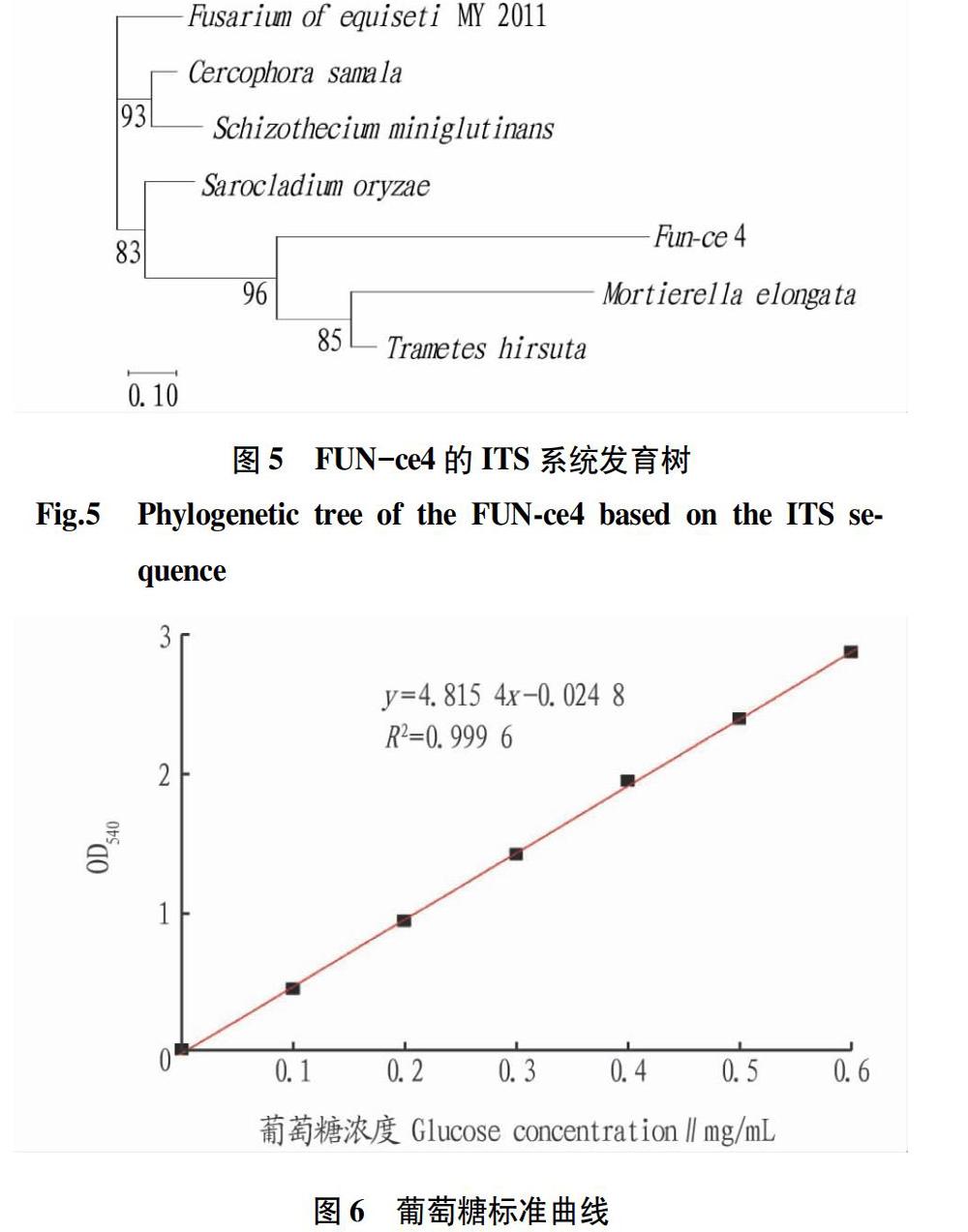
將擴增后的ITS進行凝膠電泳檢測。由圖4可知,序列2為所挑選菌株目的條帶片段。由PCR擴增得到的目的片段長度約353 bp。在GenBank的Blast中對該序列進行同源性比較(圖5),結果顯示FUN-4菌株ITS基因序列與毛栓菌(Trametes hirsuta)同源性為96%,初步鑒定為栓菌屬。
2.3 菌株纖維素酶活的測定及纖維素酶穩定性研究
2.3.1 葡萄糖標準曲線制作。
該葡萄糖標準曲線的R2=0.999 6,擬合程度較好,得到的回歸方程為y=4.815 4x-0.024 8(圖6)。可用該方程計算對應的葡萄糖含量,代入方程中計算酶活。
2.3.2 發酵時間對酶活力的影響。
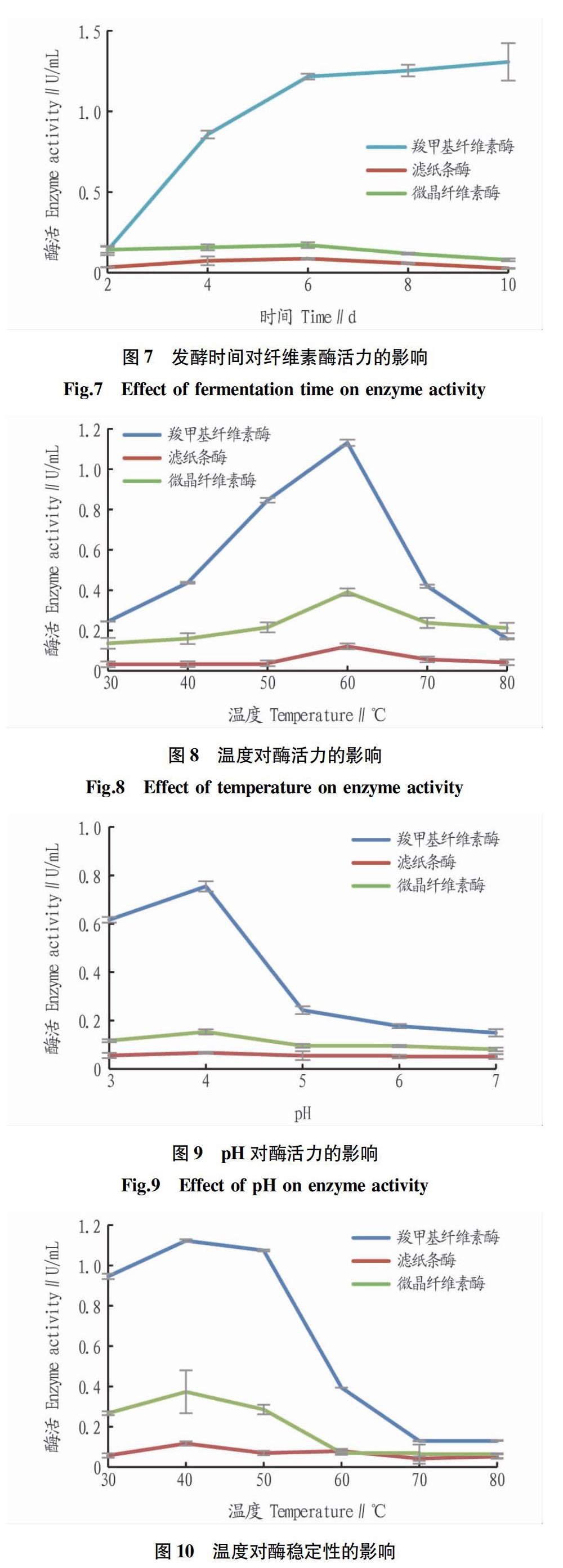
羧甲基纖維素酶活隨著時間延長逐漸增大,但在第6天之后酶活升高不明顯(圖7),考慮到時間成本,選擇第6天為最優發酵時間;濾紙條酶活和微晶纖維素酶活較低,兩者相差較小,且在第6天達到最大值,可能是由于羧甲基纖維素酶抑制了兩者的活性。因此,選擇第6天為發酵時長。
2.3.3 溫度和pH對酶活力的影響。羧甲基纖維素酶、濾紙條酶、微晶纖維素酶的酶活均在60 ℃時達到峰值,分別為1.020、0.094和0.246 U/mL。在30~50 ℃,濾紙條酶幾乎不顯示活性,80 ℃時微晶纖維素酶活有超過羧甲基纖維素酶活的趨勢(圖8)。羧甲基纖維素酶、濾紙條酶、微晶纖維素酶的酶活隨緩沖液pH升高均先增大后減小,在pH為4時達到最高值,分別為0.755、0.067、0.153 U/mL,但濾紙條酶活隨緩沖液pH升高變化不明顯(圖9)。
2.3.4 溫度和pH對酶活穩定性的影響。
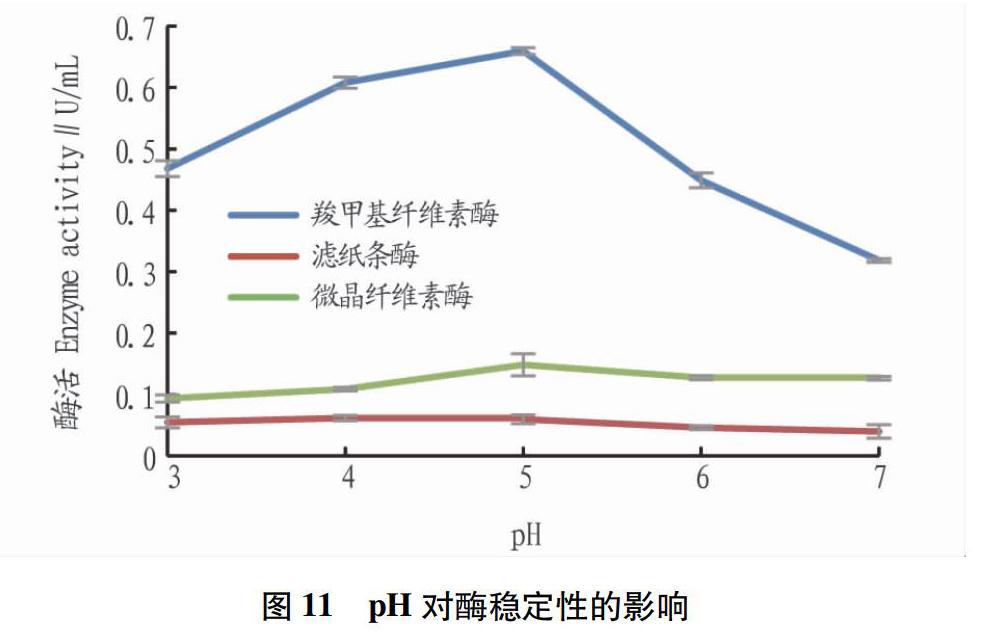
從圖10可見,保存在較低溫度下的酶穩定性較好,在40 ℃時酶活最穩定,羧甲基纖維素酶、濾紙條酶、微晶纖維素酶的酶活分別為1.123、0.117、0.373 U/mL,相對酶活力分別為93.89%、96.38%和95.64%。羧甲基纖維素酶穩定性隨溫度升高變化最大,超過50 ℃后酶活驟降。濾紙條酶活較低,但穩定性好。從圖11可見,羧甲基纖維素酶、濾紙條酶、微晶纖維素酶的酶活pH為5時酶活最穩定,分別為0.660、0.060、0.148 U/mL,相對酶活力分別為87.4%、86.4%、96.7%。羧甲基纖維素酶對pH較為敏感,濾紙條酶活和微晶纖維素酶活均較低,但穩定性好,在不同pH緩沖液下保存酶活變化較小,pH為3~7時均能保持60%以上活性。
3 小結
該研究以白蟻菌圃為菌源,相比主流研究將菌源聚集在土壤、白蟻腸道、腐爛木材等更具有創新性。試驗從菌圃中提取篩選出1株高產纖維素酶真菌,為后續白蟻菌圃混合菌群分離純化提供參考。通過分子學初步鑒定其高產纖維素酶菌株為栓孔菌屬,栓孔菌屬隸屬于擔子菌門、傘菌亞門、傘菌綱、多孔菌目、多孔菌科,屬于白腐菌的一大類,擔子門白腐真菌近年來作為產漆酶的重要優質菌源而被廣泛研究[17-20],從而為生物降解提供了重要途徑與方法。后續可對所產纖維素酶的酶活及穩定性進行初步探索,以期為其開發利用提供理論依據。
參考文獻
[1]
謝蓉蓉,孫建中,耿阿蕾,等.自然生物系統在生物質高效轉化利用中的科學價值與應用前景[J].生物產業技術,2015(2):32-40.
[2]李棟,田偉金,黎明,等.談談白蟻與人類的密切關系[J].昆蟲知識,2004,41(5):487-494.
[3]龍雁華.高等培菌白蟻高效利用木質纖維素的機理研究[D].合肥:安徽農業大學,2009.
[4]WARNECKE F,LUGINBHL P,IVANOVA N,et al.Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite[J].Nature,2007,450(7169):560-565.
[5]NIMCHUA T,THONGARAM T,UENGWETWANIT T,et al.Metagenomic analysis of novel lignocellulose-degrading enzymes from higher termite guts inhabiting microbes[J].J Microbiol Biotechnol,2012,22(4):462-469.
[6]LIU N,YAN X,ZHANG M L,et al.Microbiome of fungus-growing termites:A new reservoir for lignocellulase genes[J].Applied and environmental microbiology,2011,77(1):48-56.
[7]LIU N,ZHANG L,ZHOU H K,et al.Metagenomic insights into metabolic capacities of the gut microbiota in a fungus-cultivating termite (Odontotermes yunnanensis)[J].PLoS One,2013,8(7):1-10.
[8]POULSEN M,HU H F,LI C,et al.Complementary symbiont contributions to plant decomposition in a fungus-farming termite[J].Proceedings of the national academy of sciences,2014,111(40):14500-14505.
[9]戴芳瀾.真菌的形態和分類[M].北京:科學出版社,1987.
[10]魏景超.真菌鑒定手冊[M].上海:上海科學技術出版社,1979.
