核盤菌對大豆致病力差異與抗氧化酶系統(tǒng)酶活性的關(guān)系
2021-04-29 21:46:52李易初石鳳梅馬立功劉佳孟慶林
安徽農(nóng)業(yè)科學(xué) 2021年7期
關(guān)鍵詞:大豆
李易初 石鳳梅 馬立功 劉佳 孟慶林

摘要 采集黑龍江省大豆菌核病樣分離得到15個菌株,選用大豆離體葉片接種法鑒定菌株致病力強(qiáng)弱,測定致病力差異菌株體內(nèi)POD、SOD、CAT活性。結(jié)果發(fā)現(xiàn),黑龍江地區(qū)大豆菌核病菌致病力分化明顯,菌株致病力分化與來源地?zé)o明顯相關(guān)性;致病力差異菌株體內(nèi)POD、SOD、CAT活性存在差異;60%供試菌株體內(nèi)SOD、POD和CAT活性與致病力呈正相關(guān),菌株致病力強(qiáng)則菌株體內(nèi)相應(yīng)的酶活性強(qiáng);菌株CAT活性與其致病力相關(guān)性不明顯,需進(jìn)一步試驗驗證。
關(guān)鍵詞 大豆;核盤菌;致病力;酶活性;POD;SOD;CAT
中圖分類號 S435.651文獻(xiàn)標(biāo)識碼 A文章編號 0517-6611(2021)07-142-04
Abstract Fifteen strains were isolated from the samples of Sclerotinia sclerotiorum on soybean in Heilongjiang Province, and the pathogenicity of the strains were determined by inoculating the leaves in vitro, and the enzymes activity of POD, SOD and CAT in the strains with different pathogenicity were determined. It showed that the pathogenicity of S. sclerotiorum on soybean in Heilongjiang was differentiated obviously, and there was no obvious correlation between the pathogenicity and the collection places. The enzyme activities of POD, SOD and CAT were different in 15 strains. The enzyme activities of POD,SOD and CAT in strains were correlated with pathogenicity of the strains, and the enzyme activities of POD,SOD and CAT in the 60% strains of the test were bigger which strains with stronger pathogenicity. There was no significant correlation between the activity of CAT in strains and its pathogenicity.
Key words Soybean;Sclerotinia sclerotiorum;Pathogenic;Enzyme activity;POD;SOD;CAT
作者簡介 李易初(1986—),女,遼寧本溪人,助理研究員,碩士,從事植物病害研究。*通信作者,研究員,碩士,從事植物病害研究。
菌核病[Sclerotinia sclerotiorum(Lib.)de Bary]是危害全球大豆產(chǎn)量及品質(zhì)的一種常見病害,我國以黑龍江省、內(nèi)蒙古等地危害較重。大豆作為墾區(qū)主栽農(nóng)作物,大豆菌核病每年都有發(fā)生,一般田塊發(fā)病率達(dá)10%,重病田可達(dá)50%,減產(chǎn)可達(dá)50%以上[1]。菌核病可侵染各生育期的大豆植株,初期為水浸狀病斑,逐漸引起葉片變黃萎縮,莖桿褪綠溢縮死亡,氣候濕潤葉部病斑背面可見大量白色菌絲,死亡植株可在大豆莖稈內(nèi)部找到大量黑色鼠糞狀菌核[2]。大豆菌核病致病菌為核盤菌[Sclerotinia sclerotiorum(Lib.)de Bary],屬于子囊菌門柔膜菌目核盤菌屬的一種植物病原真菌[3]。核盤菌生態(tài)分布廣泛,可以侵染包括油菜、大豆、向日葵等多種雙子葉農(nóng)作物在內(nèi)的400多種植物[4]。核盤菌在不良生存條件下形成菌核,以菌核為休眠體可保持萌發(fā)能力存活8年甚至更久[5]。單個菌核在適宜條件下能多點多次萌發(fā)形成子囊盤,每個健康的子囊盤可持續(xù)多天噴發(fā)子囊孢子,研究發(fā)現(xiàn)核盤菌多種菌態(tài)均可完成侵染,氣生菌絲和子囊孢子都可直接侵染寄主,實際生產(chǎn)中無完全免疫大豆品種材料,因此對大豆菌核病的防治比較困難[6]。
核盤菌的致病機(jī)制仍未明確,研究發(fā)現(xiàn)核盤菌菌株在不同寄主上均表現(xiàn)出致病力分化,國外曾報道過致病力不同的核盤菌生理生化特性存在極大的差異[7],果膠酶、纖維素酶活性與核盤菌菌株致病力存在相關(guān)性[8-10],菌株體內(nèi)抗氧化酶系統(tǒng)中SOD酶、POD酶及CAT酶活性與菌株致病力關(guān)系研究較少。筆者通過對病原菌致病力差異測定,確定致病力強(qiáng)弱菌株,并比較致病力差異菌株內(nèi)3種酶的活性,揭示菌株致病力與酶活性的關(guān)系,旨在研究核盤菌致病力與病原菌體內(nèi)氧化酶的相關(guān)性,為進(jìn)一步研究大豆菌核病菌致病機(jī)制、致病力主控酶所屬基因測定提供前期研究,為更加有效地預(yù)防控制菌核病提供理論依據(jù)。
1 材料與方法
1.1 病原菌的制備
收集到的菌核樣本根據(jù)采集地點編號,通風(fēng)陰涼處晾干,晾干后存放于紙袋,室溫保存?zhèn)溆谩嶒炇页R?guī)方法分離菌核樣品,在無菌室內(nèi)用75%無水乙醇浸泡菌核表面消毒30 s,無菌水沖洗3遍,菌核表面水分用濾紙吸干,鑷子夾取菌核放在培養(yǎng)皿中,手術(shù)刀切取菌核薄片(如菌核過硬不易切片,可先用無菌水浸泡數(shù)分鐘,再重復(fù)表面消毒等后續(xù)操作再切片),接種于PDA培養(yǎng)基25 ℃恒溫培養(yǎng),菌核片長出白色菌絲,挑取菌絲轉(zhuǎn)接在新的PDA平板上,菌株分離試驗中所用器具均需提前滅菌消毒,以此方法轉(zhuǎn)接 3 次得到的菌株,可認(rèn)為遺傳穩(wěn)定的純化菌株。按照上述方法分離采集的大豆菌核樣本,經(jīng)柯赫氏法則回接試驗再分離得到的菌株,轉(zhuǎn)接保存于試管斜面,放置在4 ℃低溫備用。試驗前,菌株需提前在PDA培養(yǎng)基上活化再使用。分離得到黑龍江地區(qū)大豆菌核病致病菌,選取15個菌株進(jìn)行試驗(表1)。
1.2 大豆菌核病菌致病力差異
核盤菌的致病性測定方法主要有菌絲體接種法[11]、子囊孢子法[12]和草酸鑒定法[13]。目前致病力測定最常用的方法是菌絲體接種法,將菌核分離培養(yǎng)得到的PDA生長的菌絲體,采用菌塊貼片的方法接種到對應(yīng)寄主植物的組織上,接種部位多以植株葉片和莖桿為主[14-15]。采用室內(nèi)離體葉片菌絲體接種法,通過接種相同時間內(nèi)不同菌株侵染得到的病斑直徑大小測定菌株的致病力差異。供試菌株先在PDA培養(yǎng)基上活化相同時間,取直徑0.5 cm菌絲塊轉(zhuǎn)接在新PDA平板培養(yǎng)72 h,沿菌落外緣打取直徑0.5 cm的菌塊。溫室提前種植大豆盆栽(墾豐41),摘取帶有葉柄的同時期同位置的健康大豆葉片,一般選擇三出復(fù)葉中間卵狀復(fù)葉,將菌塊著生菌絲一面避開葉脈貼在葉片表面,接種后用手輕微按壓,確保菌塊不掉落,每個菌株設(shè)3次重復(fù),設(shè)置空白PDA菌塊為對照試驗。接種葉片統(tǒng)一盛放在鋪有濕潤無菌紗布的托盤中,濕潤脫脂棉包裹葉柄保濕,保鮮膜覆蓋托盤表面,25 ℃培養(yǎng)72 h后調(diào)查發(fā)病情況,根據(jù)十字交叉法測定病斑直徑,記錄不同菌株侵染后病斑直徑大小。按照下列公式計算病斑直徑及平均值。
病斑直徑(cm)=病斑整體直徑(cm)-菌塊直徑(cm)
1.3 強(qiáng)弱致病力差異菌株體內(nèi)氧化酶活性的測定
選擇致病力差異較大的菌株進(jìn)行氧化酶活性測定。
方法參照《植物生理生化實驗原理和技術(shù)》[16],具體試驗步驟有改動。
菌絲的收集:將直徑0.5 cm 新鮮菌塊接種到鋪有滅菌玻璃紙的PDA培養(yǎng)基中心,25 ℃培養(yǎng)箱中培養(yǎng),每菌株設(shè)3次重復(fù),菌株生長時間與致病力分化試驗的培養(yǎng)時間一致。培養(yǎng)72 h后,無菌環(huán)境下刮取玻璃紙上生長的菌絲,收集存放在5 mL離心管中。離心管中加入1 mL無菌蒸餾水,放于室溫下靜置4 h,離心機(jī)10 000 r/s離心2 min,取上清液,即為粗酶液,如不立即試驗需放置在-80 ℃冰箱保存。
按照POD 測試盒(購自上海酶聯(lián)生物技術(shù)有限公司)說明書的方法,嚴(yán)格按照操作步驟進(jìn)行過氧化物酶(POD)活性測定。
按照SOD 測試盒(購自上海酶聯(lián)生物技術(shù)有限公司)說明書的方法,嚴(yán)格按照操作步驟進(jìn)行SOD酶活性測定。
按照CAT 測試盒(購自上海酶聯(lián)生物技術(shù)有限公司)說明書的方法,嚴(yán)格按照操作步驟進(jìn)行測定。
1.4 數(shù)據(jù)分析
試驗數(shù)據(jù)用Excel 2013及SPSS 22.0 軟件進(jìn)行處理并進(jìn)行單因素方差分析。
2 結(jié)果與分析
2.1 核盤菌致病力分化
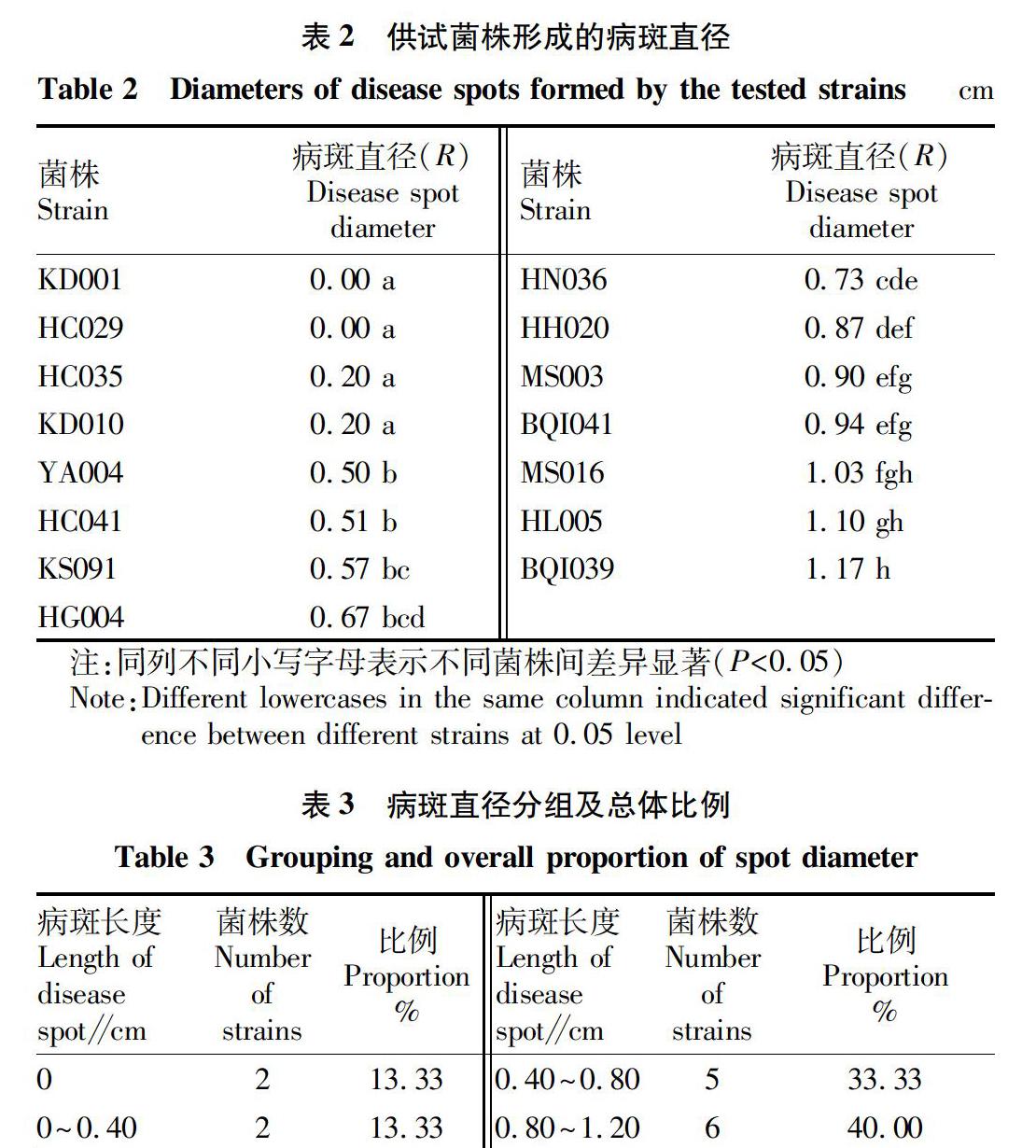
供試菌株離體葉片接菌試驗得到的病斑直徑均值見表2。由表2可知,病斑直徑最小值為0 cm,接種后葉片無病斑,菌塊著生的氣生菌絲變長,接觸葉片部位未發(fā)病;最大病斑直徑是1.17 cm,葉片表面病斑明顯,病部葉片顏色變黑、軟爛。按照病斑直徑從小到大順序?qū)┰嚲昱判颍闗D001、HC029、HC035、KD010和YA004形成病斑直徑≤0.50 cm,其中菌株KD001和HC029未形成病斑;菌株HC041、KS091、HG004、HN036、HH020、MS003和BQI041形成病斑直徑在0.50~1.00 cm,菌株MS016、HL005和BQI039形成病斑直徑較大均≥1.00 cm。根據(jù)多重比較結(jié)果可知,致病力最強(qiáng)BQI039和最弱菌株KD001和HC029之間致病力差異最顯著。供試菌株根據(jù)差異顯著性被分成8組,組間差異較大。由表3可知,致病性強(qiáng)菌株占供試菌株比例較大,侵染病斑直徑超過0.80 cm的菌株,占總供試菌株的比例最大(40.0%);病斑直徑超過0.40 cm的菌株有11株,占比為73.33%;而侵染病斑直徑在0~0.40 cm的致病力較弱菌株,僅有4株,占總菌株的26.66%。菌株形成病斑直徑差異反映出致病力強(qiáng)弱菌株間差異較大,也表明核盤菌菌株致病力存在分化現(xiàn)象。
對供試核盤菌菌株致病力測定數(shù)據(jù)的F檢驗結(jié)果表明,核盤菌菌株間致病力分化明顯,菌株間致病力存在極顯著差異,黑龍江地區(qū)引起大豆病害的核盤菌菌株存在致病力差異。結(jié)合表1菌核采集地點,相同來源地菌株HC029、HC035和HC041的病斑直徑分別是0、0.20、0.51 cm;致病力最強(qiáng)的菌株BQI039與相同采集地的菌株BQI041,病斑直徑差為0.23 cm;菌株KD001和HC029,菌株HC035和KD010的來源地均不同,但致病力表現(xiàn)相同。綜合上述情況,黑龍江省核盤菌菌株致病力與菌株來源地?zé)o明顯相關(guān)性。
2.2 強(qiáng)弱致病力差異菌株體內(nèi)氧化酶活性
2.2.1 CAT活性與致病力關(guān)系。
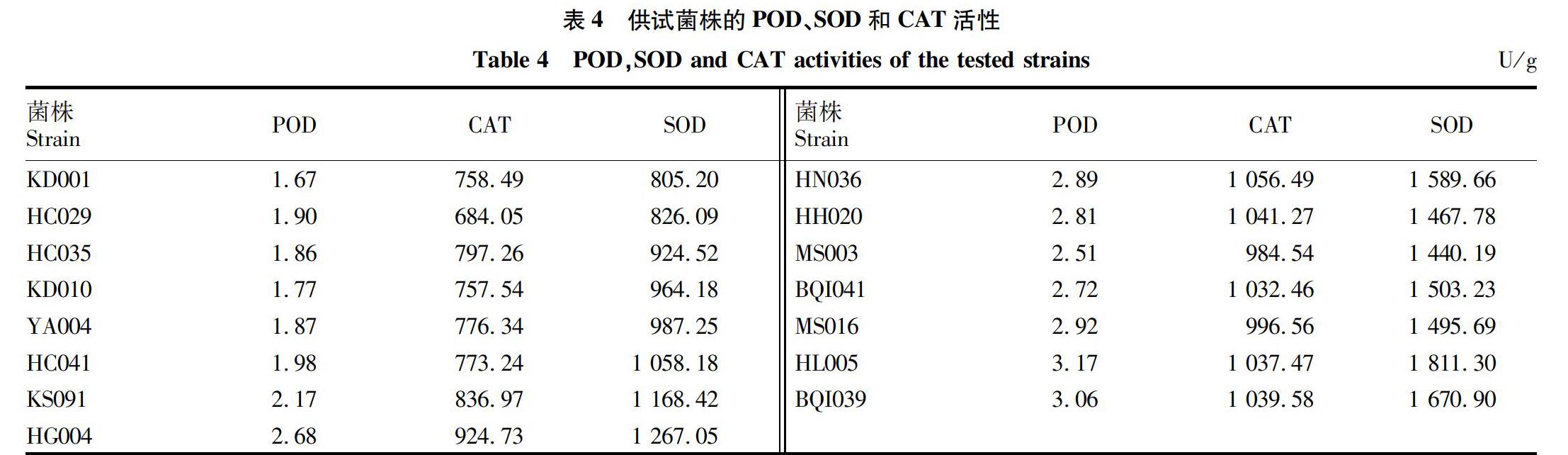
供試菌株中,CAT活性在684.05~1 056.49 U/g(表4),由菌株間CAT活性整體趨勢可知,菌株致病力強(qiáng),則菌株CAT活性強(qiáng)。其中,菌株KD001、HC035、HN036、HH020和MS016 CAT活性在總體起伏變化中與致病力強(qiáng)弱規(guī)律不相符,超過30%的供試菌株CAT活性與致病力無明顯相關(guān)性,66.7%供試菌株的致病力強(qiáng)酶活性表現(xiàn)較強(qiáng)。致病力表現(xiàn)一致的菌株KD001和HC029,CAT活性分別是758.49和684.05 U/g,相差74.44 U/g;菌株HC035和KD010的致病力表現(xiàn)也相同,菌株酶活之差是39.72 U/g,酶活強(qiáng)弱未能體現(xiàn)菌株致病力相同的特性。綜合上述情況,核盤菌菌株CAT活性總體趨勢與菌株致病力有一定的相關(guān)性,菌株CAT活性與致病性表現(xiàn)未完全一致,兩者相關(guān)性不顯著。
2.2.2 SOD活性與致病力的關(guān)系。
供試菌株SOD活性在805.20~1 811.30 U/g,酶活性最大值與最小值差超過1 000 U/g,菌株間酶活性差異較大。接種形成病斑直徑>0.5 cm的菌株,其SOD活性>1 000 U/g;病斑直徑>1.0 cm的菌株,其酶活性均>1 500 U/g。按照致病力從弱到強(qiáng)順序排列菌株,將菌株的酶活性制成柱形圖觀察(圖1),菌株致病力強(qiáng)則菌株體內(nèi)SOD活性越大。供試菌株中僅有菌株HN036和HL005較致病力相似菌株的酶活性高,未完全符合酶活性與致病力的關(guān)系,仍比致病力弱的菌株酶活大;菌株HH020酶活略高于致病力較強(qiáng)的菌株MS003;菌株KD001和HC029、菌株HC035和KD010致病力表現(xiàn)一致,菌株的SOD活性相近。供試12個菌株的SOD活性與菌株的致病力呈正相關(guān),達(dá)供試菌株的80%,致病力強(qiáng)的核盤菌菌株,自身的SOD酶活性較大。
2.2.3 POD活性與致病力的關(guān)系。
由圖2可知,供試菌株P(guān)OD活性較小,最大值未超過3.2 U/g。菌株接種形成病斑直徑≤0.5 cm,酶活性≤2.0 U/g;病斑直徑>0.5 cm,酶活性>2.0 U/g;病斑直徑>1.1 cm,酶活性>3.0 U/g。致病力最強(qiáng)菌株的POD活性是3.06 U/g,致病力最弱菌株酶活性為167 U/g,致病力強(qiáng)菌株的對應(yīng)酶活性較大,致病力弱菌株對應(yīng)酶活性較小,致病力差異菌株的POD活性也存在差異。菌株KD001和HC029致病力表現(xiàn)相同,菌株HC029酶活性高于KD001 0.23 U/g;菌株HC035與KD010致病力表現(xiàn)相同,菌株HC035酶活性較菌株KD010高0.09 U/g。菌株HC029、HC035、HG004、HN036、HH020和HL005酶活性增加幅度過大,略高于同致病力及致病力較強(qiáng)菌株,15株菌株中有60%的菌株完全符合菌株致病力強(qiáng)則菌株內(nèi)POD活性較大的規(guī)律。供試菌株P(guān)OD活性與致病力存在相關(guān)性,致病力強(qiáng)的菌株,酶活性亦較強(qiáng)。核盤菌菌株致病力強(qiáng),菌株體內(nèi)POD活性大,隨著致病力增長酶活性總體呈上升趨勢。
2.2.4 核盤菌體內(nèi)酶活性變化。
由圖3可知,CAT活性變化平緩,供試菌株的酶活性集中在600~1 100 U/g,酶活性最高值是菌株HH020的1 056.49 U/g,最低值是HC029為684.05 U/g,CAT活性最高值與致病力最強(qiáng)菌株不對應(yīng),活性最低菌株與致病力最弱菌株相對應(yīng),菌株間CAT活性變化不明顯。供試菌株間SOD活性變化較大,活性最低值來自致病力最弱的菌株KD010,為805.2 U/g;活性最高為1 811.3 U/g,來自菌株致病力第二強(qiáng)的菌株HL005,致病力最強(qiáng)菌株活性第二高;菌株間酶活性變化較大,變化幅度超過1 000 U/g,CAT及SOD 2組酶活性變化趨勢相關(guān)性不顯著。
POD活性在1.67~3.17 U/g,酶活性最小值與致病力最弱菌株相一致,酶活性最大值來自致病性第二強(qiáng)的菌株HL005,致病力最強(qiáng)菌株酶活性為3.06 U/g,且與最大酶活性相差較小。POD活性最值與SOD最值來源菌株一致,POD整體變化趨勢與SOD活性變化規(guī)律相似,核盤菌菌株體內(nèi)SOD及POD活性表現(xiàn)總體趨勢一致。
3 結(jié)論與討論
15個供試菌株中有13個菌株72 h可侵染大豆葉片使其發(fā)病,接種后病斑大小差異為菌株致病力分化表現(xiàn),核盤菌菌株間致病力存在明顯差異。黑龍江大豆菌核病菌株的致病力差異明顯,該地區(qū)核盤菌致病力存在分化現(xiàn)象,且核盤菌種群內(nèi)致病性分化明顯。導(dǎo)致大豆菌核病菌菌株致病力分化原因較多,除環(huán)境因素外,菌株致病力分化可能與菌株本身的特質(zhì)密切相關(guān)。多地點來源樣品比較發(fā)現(xiàn),同一來源地的菌株存在致病力差異;致病力表現(xiàn)相同菌株來自不同地點,核盤菌致病力差異與地域性不相關(guān)[17-18]。
供試核盤菌菌株內(nèi)抗氧化酶系統(tǒng)中CAT、SOD、POD活性與菌株的致病力有一定相關(guān)性,菌株體內(nèi)3組酶的變化趨勢與菌株致病力強(qiáng)弱表現(xiàn)存在相關(guān)性,超過60%供試菌株符合致病力越強(qiáng),酶活性越大。3組酶中,菌株間SOD活性差異最大,與菌株致病力相關(guān)性最大,80%供試菌株符合致病力越強(qiáng),菌株體內(nèi)SOD活性越強(qiáng);菌株內(nèi)POD與SOD活性變化總體趨勢一致,超過60%的菌株致病力強(qiáng)弱與酶活性強(qiáng)弱正相關(guān);菌株間CAT活性變化不大,且CAT與SOD活性變化趨勢相關(guān)性不顯著,菌株CAT活性與致病力表現(xiàn)未完全一致,仍有待進(jìn)一步驗證。
參考文獻(xiàn)
[1] 王云龍.淺析大豆菌核病發(fā)病原因及防治重點[J].農(nóng)業(yè)與技術(shù),2018,38(18):33.
[2] 李憲偉.大豆病蟲害防治技術(shù)淺析[J].農(nóng)民致富之友,2015(23):94.
[3] 羊國根,程家森.核盤菌致病機(jī)理研究進(jìn)展[J].生物技術(shù)通報,2018,34(4):9-15.
[4] BOLAND G J,HALL R.Index of plant hosts of Sclerotinia sclerotiorum[J].Can J Plant Pathol,1994,16(2):93-108.
[5] ADAMS? P B, AYERS W.Ecology of Sclerotinia species[J].Phytopathology,1979,69(8):896-899.
[6] 唐慶華,宋薇薇,朱輝,等.大豆菌核病及其病原核盤菌研究進(jìn)展[C]∥中國植物病理學(xué)會.中國植物病理學(xué)會2011年學(xué)術(shù)年會論文集.北京:中國農(nóng)業(yè)科學(xué)技術(shù)出版社,2011:58-64.
[7] 母玉翠.核盤菌(Sclerotinia sclerotiorum)致病性分化的生化機(jī)制研究[D].合肥:安徽農(nóng)業(yè)大學(xué),2013.
[8] 黃娟,李國慶.兩株毒力不同的核盤菌產(chǎn)草酸、果膠酶的比較[J].氨基酸和生物資源,2008,30(2):5-8.
[9] POUSSEREAU N,CRETON S,BILLONGRAND G,et al.Regulation of acp1,encoding a nonaspartyl acid protease expressed during pathogenesis of Sclerotinia sclerotiorum[J].Microbiology,2001,147(3):717-726.
[10] POUSSEREAU N,GENTE S,RASCLE C,et al.aspS encoding an unusual asparty1 protease from Sclerotinia sclerotiorum is expressed during phytopathogenesis[J].FEMS Microbiology Letters,2001,194(1):27-32.
[11] 劉勇,布朗·霍爾騰斯.油菜苗期室內(nèi)抗菌核病性研究——Ⅰ.離體葉片子囊孢子接種鑒定[J].西南農(nóng)業(yè)學(xué)報,1993,6(S1):74-78.
[12] 劉勇,布朗·特伯德.油菜菌核病田間抗性鑒定和篩選——I.火柴棍莖桿菌絲接種法[J].西南農(nóng)業(yè)學(xué)報,1993,6(S1):42-46.
[13] 劉勝毅,周必文,潘家榮.油菜對毒素草酸的吸收代謝與抗性機(jī)理[J].植物病理學(xué)報,1998,28(1):33-37.
[14] KULL L S,VUONG T D,POWERS K S,et al.Evaluation of resistance screening methods for Sclerotinia stem rot of soybean and dry bean[J].Plant disease,2003,87(12):1471-1476.
[15] 劉丹,李沛利,嚴(yán)吉明,等.核盤菌致病性測定方法研究[J].安徽農(nóng)業(yè)科學(xué),2011,39(30):18627-18629.
[16] 王學(xué)奎.植物生理生化實驗原理和技術(shù)[M].2版.北京:高等教育出版社,2006:134-136.
[17] 李建廠,李永紅,陳文杰,等.向日葵核盤菌菌株致病性研究及其溫度效應(yīng)[J].西北農(nóng)業(yè)學(xué)報,2003,12(1):114-117.
[18] 李沛利,葉華智.核盤菌致病性分化研究[J].植物保護(hù),2006,32(5):29-31.
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
