林隙對桂北山區常綠闊葉林物種更新的影響
2021-05-19 12:00:08鄧善寶
湖北農業科學 2021年9期
關鍵詞:物種
鄧善寶
(廣西壯族自治區國有維都林場,廣西 來賓 546100)
林隙(Gap)是林冠層樹木死亡之后形成的林分空隙,空隙的產生為新物種的產生提供了空間。這種物質能量環境便于植物在林窗內正常更新并形成更替層[1-3]。林隙的形狀[4,5]、林隙的大小[6-8]、林隙的年齡[9]、林隙的形成木及邊緣木[10]構成了林隙的基本特征,林隙對森林樹種空間分布結構[11-14]和森林群里的更新[15-18]具有重要的影響。為此,本研究以2018 年冰凍災害后廣西三江縣獨峒造林點常綠闊葉林為研究對象,通過野外調查林隙的具體特征及幼苗樣方調查分析,揭示林隙對常綠闊葉林物種更新的影響,以期為三江縣災后生態系統的修復提供科學根據。
1 材料與方法
1.1 試驗地概況
三江侗族自治縣獨峒鎮位于廣西北部(東經108°53′—109°52′,北緯 25°22′—26°2′),是湘、桂、黔三省(區)交界地。屬中亞熱帶、南嶺濕潤氣候區,1 月最冷,平均為 5.1 ℃;7 月最熱,平均為 27.4 ℃。年平均雨量1 493 mm,夏季為降雨高峰季節,占全年的42%~48%;春季為降雨次高峰期,占全年的30%~35%。年均日照數1 333.3 h,年均無霜期321 d,年均降雪5 d,年平均結冰7.2 d。三江縣獨峒鎮屬九萬大山穹褶帶和龍脈褶斷帶之間,海拔850 m 以上為黃壤地帶性土壤,土層肥厚,有機質含量高。 海拔500~800 m,生長有栲類、櫟類、酸棗、擬赤揚、楠木、楓香、光皮樺等;海拔800 m 以上山地,多為水源林和灌木叢為主,原生植被為闊葉林,主要樹種有山毛櫸、青崗櫟、麻櫟、荷木、楓木、山槐等,山頂為苔蘚短曲林帶,并伴生五節芒、鐵芒箕、纖毛鴨咀草、野古草、四金脈等禾草群叢。
1.2 試驗方法
2019 年采用全站儀將3.2 hm2樣地劃分出80 個20 m×20 m 的樣方,再把每個樣方劃分成4 個10 m×10 m 和 16 個5 m×5 m 的小樣方。野外調查以 5 m×5 m 小樣方為測樹單元進行群落調查,標記DBH≥1.0 cm 的木本植物,記錄物種名、胸徑、樹高、枝下高、冠幅等數據。每個20 m×20 m 樣方中靠近原點的一個5 m×5 m 小樣方作為灌草層的調查樣方,記錄DBH<1.0 cm 灌木、藤本和草本植物的物種名、個體數、高度、蓋度等數據,喬木樹種幼苗分為小于10 cm、10~20 cm、20~30 cm、30~50 cm 和大于 50 cm 5 個高度級記錄個體數。在樣地內尋找每一個林隙,并根據樣地內每棵樹木的坐標和樹冠投影圖,采用坐標方格紙繪制出擴展林隙的多邊形平面圖,依據多邊平面圖和樹冠投影圖精確計算冠空隙(canopy gap,CG)和擴展林隙(expanded gap,EG)的面積,擴展林隙大小由邊界木基部組成的多邊形平面圖求算,冠空隙大小由林隙邊界木樹冠投影圖的空隙求算。計算每個林隙木本植物各層次的物種的重要值和每個01 號小樣方灌木、草本、藤本和幼苗的重要值,再以此計算物種更新的重要值位序。采用下列指數分析林隙和非林隙林分的物種多樣性:
物種重要值IV:
IV=(相對多度+相對頻度+相對顯著度)/3(DBH≥1.0 cm 的木本植物)
IV=(相對多度+相對頻度+相對蓋度)/3(灌木、草本和藤本)
Pielou 均勻度指數(J):J=H′/lnS式中:H′為Shannon-Wiener 指數;S為物種數。
1.3 統計分析方法
采用Excel 2007 對數據進行整理和計算。采用SPSS 19.0 進行方差分析、相關分析等統計分析。
2 結果與分析
2.1 林隙大小
圖1a 為樣地森林林隙大小的分布情況,常綠闊葉林中擴展林隙(EG)最小面積為67 m2,最大面積1 149 m2,平均值為 258.2 m2,EG 基本上在 70~600 m2,而其中大多數EG 的面積都在80~300 m2,占總數的65.9%。從冠空隙(CG)來看,CG 最小面積為28 m2,最大面積 928 m2,平均值為 199.3 m2,大多數CG 面積為 50~200 m2,占總數的 62.4%。圖 1b 為樣地森林林隙大小的分布情況,大于300 m2的EG 數量比例只有27.1%,但其面積比例達54.1%。擴展林隙(EG)和冠空隙(CG)在平均面積組成上有一定差異,是邊緣木的林冠所導致,可見邊緣木的樹冠伸出面積占有一定比例,對林窗的內部微環境起著一定的作用。
2.2 林隙對主要喬木樹種更新的影響
對每個物種都給出其在林隙內的重要值位序(Og)和非林隙林分中的重要值位序(Ong)。把只在林隙內出現,而在非林隙內不出現或很少出現的物種,稱為林隙更新物種(gap species);把只在非林隙林分中出現,而在林隙中沒有或很少出現的物種稱為非林隙更新物種(non gap species);將在林隙內的重要值位序先于非林隙林分中位序(Og-Ong<0)的物種稱為對林隙產生正更新反應的物種;將Og-Ong>0 的物種稱為對林隙產生負更新反應的物種;將|Og-Ong|<5 的物種,規定為對林隙更新反應不顯著物種;將 5≤ |Og-Ong|≤10 的物種,規定為對林隙更新反應中等的物種;將|Og-Ong|>10 的物種,規定為對林隙更新反應顯著的物種。結合表1 及表2 可知,薯豆杜英在林隙中的重要值為1.88,重要值位序為12,在非林隙中重要值為1.12,重要值位序為26,更新策略表現為顯著正更新。異葉榕在林隙中的重要值為1.66,重要值位序為16,在非林隙中重要值為1.09,重要值位序為28,更新策略表現為顯著正更新。南嶺槭在林隙中的重要值為1.51,重要值位序為20,在非林隙中重要值為0.37,重要值位序為55,更新策略表現為顯著正更新。天目紫莖在林隙中的重要值為1.01,重要值位序為29,在非林隙中重要值為2.06,重要值位序為11,更新策略表現為顯著負更新。深山含笑在林隙中的重要值為0.65,重要值位序為38,在非林隙中重要值為1.20,重要值位序為25,更新策略表現為顯著負更新。糙皮樺在林隙中的重要值為0.56,重要值位序為43,在非林隙中重要值為2.48,重要值位序為10,更新策略表現為顯著負更新。可以看出網脈瓊楠、百色猴歡喜、羅浮栲為中等正更新作用,薯豆杜英、異葉榕、南嶺槭為顯著正更新作用,香楠、桂南木蓮、毛折柄茶為中等負更新作用。說明林隙的干擾對于樹種更新的影響較大,且有些樹種之間表現了明顯的更新差異,林隙的出現使樹種的更新機制出現差異。
2.3 林隙對主要灌木物種更新的影響
由表3 可知,在林隙與非林隙中的匙萼柏拉木、草珊瑚、高粱泡、栓皮木姜子的密度與其他樹種相比偏大,匙萼柏拉木、栓皮木姜子、東方古柯、羅浮槭、毛狗骨柴、序葉苧麻、方枝假衛矛、黃杞、異葉榕、野黃桂、網脈瓊楠、天目紫莖在林隙內外的分布密度差異較大,序葉苧麻的幼苗只出現在林隙中,而在非林隙中未出現。林隙內外的陽光、濕度、土壤理化性質、溫度有所差別,較為喜陽的物種更新更快,使得林隙內物種多樣性增加。林隙內外灌木植物幼苗、幼樹在組成上有差異的主要物種為南嶺槭、小柱懸鉤子、百色猴歡喜、細枝柃、常山、滇粵山胡椒、方枝假衛矛、黃杞、異葉榕、野黃桂、網脈瓊楠、天目紫莖。
2.4 林隙對主要草本植物更新的影響
由表4 可知,林隙的出現主要是影響了蕨類植物的更新,其中陳氏耳蕨、赤車、白子菜有著顯著的正更新作用,對于波紋蕗蕨、黃金鳳、鈍齒樓梯草有著顯著的負更新作用,對于倒掛鐵角蕨、瓶蕨、中華復葉耳蕨有著較為顯著的負更新作用。
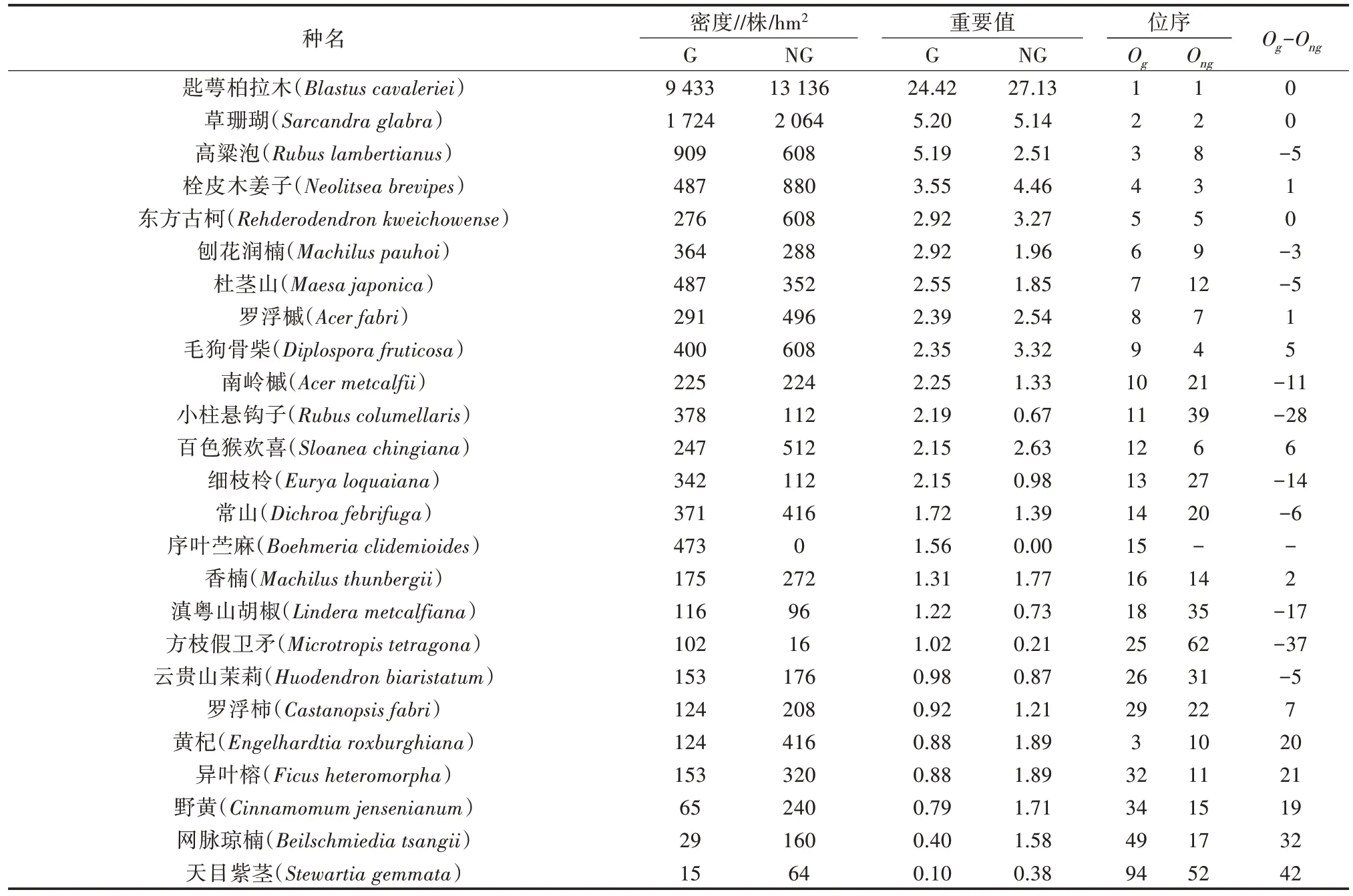
表3 林隙與非林隙林分中主要灌木植物的數量特征

表4 林隙與非林隙林分中主要草本植物的數量特征
2.5 林隙對藤本植物的更新影響
由表5 可知,林隙對于苦郎藤和細圓藤起著顯著的正更新作用,對于獨山瓜馥木則是顯著的負更新作用,對于羊角藤、雞失藤是中等的負更新作用。林隙的出現對于藤本的更新與抑制作用很明顯,林隙的抑制作用稍強于非林隙。
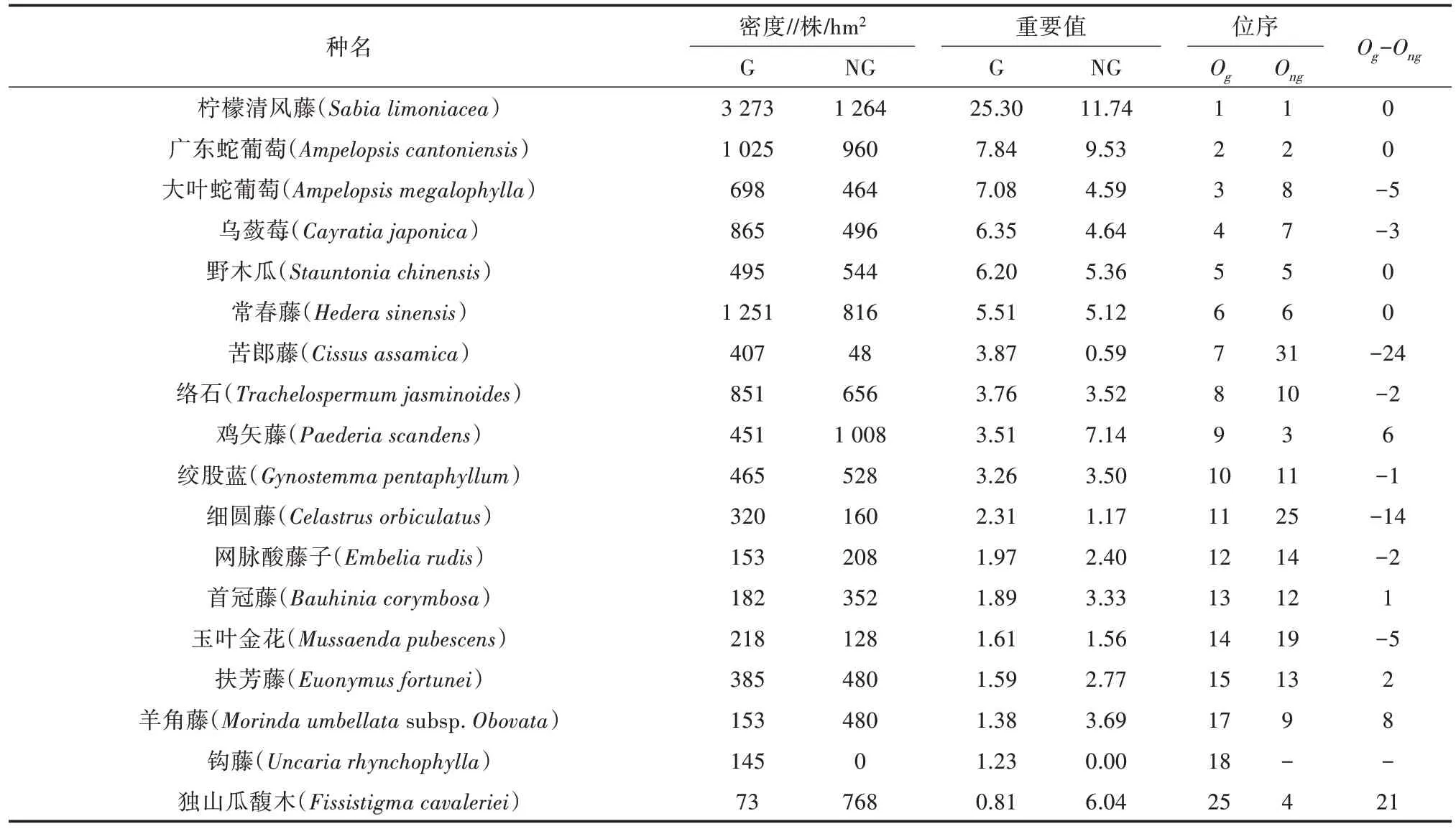
表5 林隙與非林隙林分中主要藤本植物的數量特征
3 小結與討論
林隙的形成促使林分中微環境發生改變,從而影響了物種的演替與更新,促進了適應林隙環境的物種更新生長,生態系統的動態穩定和物種多樣性得以維持。林隙干擾特征及物種更新機制的深入研究為生物多樣性的維持、森林經營管理等提供了可靠的依據。
通過林隙基本特征的調查,擴展林隙的面積變化在70~600 m2,平均面積為258.2 m2,冠空林隙的面積在50~200 m2,平均面積為199.3 m2;調查到85個林隙,林隙密度為每公頃 26.6 個,CG 總面積16 940 m2,占樣地面積52.9%;EG 總面積22 947.5 m2,占樣地面積71.7%。
通過對比林隙中主要喬木樹種和灌木層的重要值與重要值位序,喬木樹種中網脈瓊楠、百色猴歡喜、羅浮栲對于林隙是中等的正更新反應,薯豆杜英、異葉榕、南嶺槭對林隙是強烈的正更新反應;對于負更新反應方面,香楠、桂南木蓮、毛折柄茶對于林隙的反應是中等的負更新,天目紫莖、深山含笑、糙皮樺對于林隙是強烈的負更新反應,因此選取上述12 種喬木樹種為研究對象。同時灌木樹種更新反應中正更新反應程度從小到大為高粱泡、杜莖山、南嶺槭、細枝柃、滇粵山胡椒、小柱懸鉤子、方枝假衛矛;負更新反應程度從小到大為毛狗骨柴、百色猴歡喜、羅浮柿、野黃桂、黃杞、異葉榕、網脈瓊楠、天目紫莖。選取上述灌木中的喬木樹種為研究對象,對比林隙更新中主要喬木樹種與灌木中的喬木樹種,從上述可知,林分中林隙的干擾使得林隙內外中的喬木樹種產生更新反應,南嶺槭在林隙內正更新反應最為明顯,優勢度最大且將會成為林隙中的優勢樹種;薯豆杜英、異葉榕、百色猴歡喜、羅浮柿、網脈瓊楠的數量將會維持不變,且在其后續發展中優勢度會有所降低;天目紫莖、深山含笑、糙皮樺、毛折柄茶、桂南木蓮、香楠在林隙中的數量將會逐漸減少,優勢度也會逐漸降低。南嶺槭重要值位序差為-35,正更新反應極顯著,在灌木調查中其更新反應為顯著正更新,可知南嶺槭對霜凍災害的反應最為明顯且優勢度將逐漸增大。薯豆杜英、異葉榕的重要值位序差為-14 和-12,均表現為顯著的正更新,百色猴歡喜、羅浮栲、網脈瓊楠的重要值位序差為-6、-9、-5,均表現為中等正更新,它們相應的灌木更新表現為負更新或零更新,后續干擾中優勢度逐漸降低。天目紫莖、深山含笑、糙皮樺、毛折柄茶、桂南木蓮、香楠的更新策略均為負更新,相應的灌木更新表現為負更新或零更新,在它們的后續發展中優勢度逐漸降低且逐漸被其他樹種所取代。藤本、草本植物在林隙內外更新反應也較為顯著。
還通過對比草本植物、藤本植物林隙內外的密度、重要值、重要值位序,可以看到植物對林隙的更新反應差異明顯,其原因可能是,林隙內的微環境在陽光、濕度、溫度、土壤的理化性質等自然條件有別于林隙外的環境,因此,喜陽物種表現出對林隙有正更新反應,而喜陰物種表現出對林隙有負更新反應。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
