盤縣火腿深度腐敗的微生物及揮發(fā)性風(fēng)味化合物表征
2021-05-19 07:05:40母應(yīng)春
食品科學(xué) 2021年8期
母 雨,蘇 偉,*,母應(yīng)春
(1.貴州大學(xué)釀酒與食品工程學(xué)院,貴州 貴陽 550025;2.貴州大學(xué) 貴州省農(nóng)畜產(chǎn)品貯藏與加工重點(diǎn)實(shí)驗(yàn)室,貴州 貴陽 550025)
干腌火腿是一種傳統(tǒng)發(fā)酵肉制品,有數(shù)千年的消費(fèi)歷史。已經(jīng)證明干腌火腿富含多種營養(yǎng)物質(zhì)并具有潛在健康益處[1-2]。盤縣火腿是我國著名干腌火腿之一,于2012年獲得“國家地理標(biāo)志”稱號(hào)[3]。它的加工工藝包括鮮腿老化、腌制、堆碼翻壓、靜置和成熟,周期長達(dá)12~24 個(gè)月,長時(shí)間的成熟和獨(dú)特的地理環(huán)境賦予其鮮明的品質(zhì)特征。然而,盤縣火腿的生產(chǎn)采用自然發(fā)酵,且不使用任何防腐劑,其穩(wěn)定性完全依賴于鹽的滲透和成熟過程的脫水。因此,一些腐敗微生物很可能在加工早期生長繁殖,并最終導(dǎo)致產(chǎn)品的腐敗變質(zhì)。
干腌火腿的變質(zhì)是一個(gè)長期性、世界性的難題。據(jù)報(bào)道,西班牙每年有大約3 000萬 只火腿腐敗變質(zhì),造成的經(jīng)濟(jì)損失高達(dá)數(shù)千萬美元[4]。干腌火腿常見的腐敗類型有“深度腐敗”和“靜脈缺陷”。其中,“深度腐敗”是最重要的一種變質(zhì)類型,其特征是糊狀質(zhì)地和令人討厭的臭味,這種臭味常見于鄰近骨骼結(jié)構(gòu)的大塊肌肉中[5]。研究認(rèn)為這種類型的腐敗是由微生物菌群,特別是細(xì)菌造成的,且已基于培養(yǎng)法分離出相應(yīng)菌株[6]。“靜脈缺陷”屬于表層腐敗,腐敗區(qū)域通常具有較高的鹽含量,已從此類腐敗中培養(yǎng)分離出一些嗜鹽菌,如肉桿菌(Carnobacterium)、棒狀桿菌(Corynebacterium)、弧菌(Vibrio)和海生乳桿菌(Marinilactibacillus)[7-8]。然而,由于可存活但不可培養(yǎng)狀態(tài),培養(yǎng)法僅能分離出完整微生物種群的一部分[9]。相比之下,高通量測序可以對(duì)復(fù)雜的微生物群進(jìn)行更深入、更精確的評(píng)估,已用于揭示干腌火腿的群落組成[10]。此外,由于“深度腐敗”的主要缺陷在于異味,采用固相微萃取結(jié)合氣相色譜-質(zhì)譜(solid phase microextraction-gas chromatography-mass spectrometry,SPME-GC-MS)法檢測腐敗火腿的標(biāo)志性風(fēng)味。
本研究旨在表征與盤縣火腿“深度腐敗”有關(guān)的微生物和揮發(fā)性風(fēng)味,其豐度和含量可用作確定盤縣火腿變質(zhì)的指標(biāo),有助于發(fā)現(xiàn)和控制早期的腐敗。
1 材料與方法
1.1 材料與試劑
盤縣火腿樣品(正常和腐敗火腿各3 只)由貴州省盤州市火腿加工廠提供,所有樣品均取自相同工藝、環(huán)境下生產(chǎn)的同一批次火腿。為盡可能減少影響因素,正常火腿取樣部位與腐敗火腿一致。取樣完成后立即裝入無菌袋,并置于低溫泡沫盒中12 h內(nèi)送回實(shí)驗(yàn)室,貯存于-80 ℃直至分析。
Power Soil?DNA提取試劑盒 美國MP Bio公司;瓊脂糖 西班牙Biowest公司;DNA聚合酶 大連寶生物工程有限公司;正反引物 深圳市英俊生物技術(shù)有限公司;DNA純化磁珠 南京諾唯贊生物科技有限公司;MinElute?聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)純化試劑盒 德國Qiagen公司。
1.2 儀器與設(shè)備
Veriti? 9902 PCR儀 美國ABI公司;Nanodrop 2000超微量分光光度計(jì)、Trace1300-TSQ8000 GC-MS聯(lián)用儀 美國Thermo Fisher Scientific公司;Illumina MiSeq測序儀 美國Illumina公司。
1.3 方法
1.3.1 理化指標(biāo)的測定
水分含量測定:利用直接干燥法;鹽含量測定:采用GB 5009.44—2016《食品中氯化物的測定》方法;水分活度(aw)測定:水分活度儀;pH值測定:pH計(jì)。所有實(shí)驗(yàn)重復(fù)3 次。
1.3.2 高通量測序
1.3.2.1 總DNA提取與PCR擴(kuò)增
每個(gè)樣品在無菌條件下取10 g并充分研磨,然后根據(jù)說明書使用Power Soil DNA試劑盒提取樣品總DNA。DNA的質(zhì)量和數(shù)量以O(shè)D260nm/OD280nm和OD260nm/OD230nm的比值進(jìn)行評(píng)估。然后將DNA保存在-80 ℃,直到進(jìn)一步處理。利用引物(338F:5′-ACTCCTACGGGAGGCAGCA-3′和806R:5′-GGACTACHVGGGTWTCTAAT-3′)結(jié)合適配器序列和條形碼序列擴(kuò)增細(xì)菌16S rRNA的V3-V4區(qū)。
PCR擴(kuò)增體系的總量為50 μL,包括10 μL緩沖液、0.2 μL Q5 High-Fidelity DNA聚合酶、10 μL高GC增強(qiáng)劑、1 μL核苷酸、正反引物各10 μmol/L和60 ng的基因組DNA。擴(kuò)增程序?yàn)椋?5 ℃預(yù)變性5 min;95 ℃變性1 min,50 ℃退火1 min,72 ℃延伸1 min,共15 個(gè)循環(huán);72 ℃擴(kuò)展7 min。利用VAHTSTM DNA磁珠純化第1輪PCR擴(kuò)增產(chǎn)物。第2輪PCR擴(kuò)增體系為40 μL,包括20 μL 2×Phusion HF MM、8 μL ddH2O、正反引物各10 μmol/L以及第1輪擴(kuò)增的PCR產(chǎn)物10 μL。擴(kuò)增程序?yàn)椋?5 ℃預(yù)變性30 s;98 ℃變性10 s,65 ℃退火30 s,72 ℃延伸30 s,10 個(gè)循環(huán);72 ℃擴(kuò)展5 min。利用PCR純化試劑盒對(duì)瓊脂糖凝膠回收的PCR產(chǎn)物進(jìn)行純化,然后用Nanodrop 2000超微量分光光度計(jì)定量并按每個(gè)樣本的測序要求進(jìn)行混合,最后使用Illumina HiSeq 2500平臺(tái)對(duì)純化的混合樣本進(jìn)行細(xì)菌rRNA基因的高通量測序分析。
1.3.2.2 測序數(shù)據(jù)處理
根據(jù)雙端測序之間的overlap關(guān)系,利用FLASH v1.2.7軟件,通過overlap對(duì)每個(gè)樣品的序列進(jìn)行拼接,得到的拼接序列即原始序列數(shù)據(jù);然后利用Trimmomatic v0.33軟件對(duì)拼接后的原始數(shù)據(jù)進(jìn)行過濾,得到高質(zhì)量的序列數(shù)據(jù);最后,使用UCHIME v4.2軟件,鑒定并去除嵌合體序列,得到有效數(shù)據(jù)。利用QIIME v1.8.0對(duì)有效數(shù)據(jù)在97%的相似度水平下進(jìn)行聚類、獲得操作分類單元(operational taxonomic units,OTU),并基于Silva分類學(xué)數(shù)據(jù)庫對(duì)OTU進(jìn)行分類學(xué)注釋,設(shè)置比對(duì)閾值為70%。
1.3.3 揮發(fā)性風(fēng)味化合物分析
1.3.3.1 風(fēng)味化合物的提取與檢測
參照Maru?i?等[11]的方法提取和檢測揮發(fā)性風(fēng)味化合物,并略作修改。準(zhǔn)確稱取5.00 g切碎的火腿樣品置于25 mL飽和食鹽水中,均質(zhì)2 min以制備火腿勻漿。取10 mL勻漿置于20 mL頂空瓶中并用PTFE隔膜蓋緊,然后使用已老化的50/30 μm DVB/CAR/PDMS萃取頭在40 ℃振搖提取180 min。萃取完成后,立即將SPME纖維插入進(jìn)樣口,在230 ℃解吸5 min。
色譜條件:毛細(xì)管柱為DB-5MS(30 m×0.25 mm,0.25 μm),載氣為氦氣,流速為1.0 mL/min,不分流模式。柱升溫程序:50 ℃保持5 min,然后以5 ℃/min的速率升至200 ℃,最后以20 ℃/min的速率升至250 ℃并保持10 min。
質(zhì)譜條件:電子電離源,電子能量為70 eV,傳輸線和離子源溫度為280 ℃和230 ℃,掃描范圍m/z50~450,掃描速率為1 scan/s。
1.3.3.2 風(fēng)味化合物的定性與定量
定性:實(shí)驗(yàn)結(jié)果與NIST數(shù)據(jù)庫進(jìn)行比對(duì),僅保留正反匹配度均大于800的化合物。在相同的色譜條件下運(yùn)行C6~C30正構(gòu)烷烴混合標(biāo)品,以計(jì)算化合物的保留指數(shù)(retention index,RI)。計(jì)算公式如下:
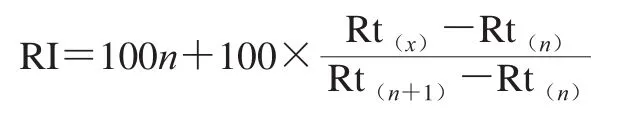
式中:Rt(x)、Rt(n)及Rt(n+1)分別為待測揮發(fā)性成分、含n個(gè)碳原子正構(gòu)烷烴及(n+1)個(gè)碳原子正構(gòu)烷烴的保留時(shí)間。
定量:峰面積歸一化法。
1.4 數(shù)據(jù)分析
SPSS Statistics 20.0用于數(shù)據(jù)的統(tǒng)計(jì)分析,數(shù)據(jù)均以表示,利用單因素方差分析確定顯著性(P<0.05)。基于獲得的OTU表格,利用QIIME計(jì)算樣品的α多樣性,包括豐富度指數(shù)(ACE和Chao1)和多樣性指數(shù)(Shannon和Simpson)。揮發(fā)性風(fēng)味化合物的LEFSe(LDA Effective Size)分析在Galaxy網(wǎng)站上進(jìn)行(http://huttenhower.sph.harvard.edu/galaxy/)。環(huán)境因子與細(xì)菌屬之間的冗余分析(redundancy analysis,RDA)基于R v3.6.1軟件的vegan包執(zhí)行。基于Pearson相關(guān)系數(shù)(r>0.7,P<0.05)建立微生物屬與揮發(fā)性風(fēng)味物質(zhì)間的相互關(guān)系,并利用Cytoscape v3.2.1軟件進(jìn)行網(wǎng)絡(luò)可視化。
2 結(jié)果與分析
2.1 物理化學(xué)特性

表1 正常與腐敗盤縣火腿理化特性Table 1 Physicochemical characteristics of normal and spoiled Panxian hams
如表1所示,腐敗火腿的pH值、aw、水分含量顯著高于正常火腿,而NaCl含量顯著低于正常火腿(P<0.05)。Blanco等[4]在腐敗西班牙火腿中發(fā)現(xiàn)了類似的情況。人工上鹽易導(dǎo)致鹽分布和滲透不均勻,因此低NaCl含量可能與腌制工藝有關(guān)。較高的pH值是產(chǎn)品變質(zhì)的指標(biāo)之一,腐敗火腿中pH值的上升可能與其較低的鹽含量有關(guān)。據(jù)報(bào)道,鹽質(zhì)量分?jǐn)?shù)5%~6%即可抑制肌肉組織蛋白酶活力,因此,較低濃度的NaCl導(dǎo)致過度的蛋白水解,從而產(chǎn)生糊狀和升高pH值[12]。另外,pH值越高持水能力越強(qiáng)[13],這使得腐敗火腿的水分含量高于正常火腿。aw與NaCl含量密切相關(guān),腐敗火腿中較低的NaCl含量導(dǎo)致較高的aw,Losantos等[14]報(bào)道了aw的下降對(duì)腐敗干腌火腿中腸桿菌科微生物的抑制作用。
2.2 細(xì)菌多樣性分析
利用Illumina MiSeq平臺(tái)從6 個(gè)樣品中獲得282 148 個(gè)高質(zhì)量16S rRNA序列,平均每個(gè)樣品產(chǎn)生47 024.67 個(gè)序列。如表2所示,正常和腐敗火腿在OTU數(shù)目和豐富度指數(shù)上不存在顯著差異(P>0.05),但在多樣性指數(shù)上存在顯著差異(P<0.05)。表明腐敗盤縣火腿中存在的細(xì)菌種類可能低于正常火腿。另外,正常和腐敗火腿覆蓋率均為0.999,表明該抽樣方案已經(jīng)基本覆蓋了樣本的細(xì)菌多樣性,數(shù)據(jù)可用于后續(xù)分析。

表2 正常與腐敗盤縣火腿的α多樣性分析Table 2 α-Diversity analysis of bacterial communities in normal and spoiled Panxian hams
為進(jìn)一步了解正常和腐敗火腿的細(xì)菌群落結(jié)構(gòu),在門和屬水平上對(duì)測序數(shù)據(jù)進(jìn)行分類。在門水平上(圖1A),鑒定出變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)、放線菌門(Actinobacteria)和擬桿菌門(Bacteroidetes)。其中,厚壁菌門是正常火腿中的優(yōu)勢(shì)菌門,而變形菌門主導(dǎo)腐敗火腿。在屬水平上(圖1B),僅顯示豐度前10的優(yōu)勢(shì)微生物,分別為葡萄球菌屬(Staphylococcus)、norank_Enterobacteriaceae、unclassified_Enterobacteriaceae、鹽單胞菌屬(Halomonas)、沙雷氏菌屬(Serratia)、段桿菌屬(Brevibacterium)、色鹽桿菌屬(Chromohalobacter)、涅斯捷連科氏菌屬(Nesterenkonia)、Solitalea和鹽水球菌屬(Salinicoccus)。

圖1 正常和腐敗盤縣火腿在門水平(A)和屬水平(B)上的細(xì)菌群落組成Fig. 1 Bacterial community structures of normal and spoiled Panxian hams at phylum (A) and genus (B) levels
與大多數(shù)干腌火腿一樣[15-16],葡萄球菌屬是正常火腿中最豐富的。由于較強(qiáng)的外源酶活力,包括蛋白酶、脂肪酶、過氧化氫酶和硝酸鹽還原酶[17],凝固酶陰性葡萄球菌被認(rèn)為是肌肉酶之后導(dǎo)致干腌火腿風(fēng)味、顏色和質(zhì)地發(fā)展的重要因素[18]。鹽單胞菌屬是正常火腿中的次要優(yōu)勢(shì)菌,與之前的報(bào)道一致[19]。在腐敗火腿中,腸桿菌科微生物(norank_Enterobacteriaceae、unclassified_Enterobacteriaceae和沙雷氏菌屬)占樣品總豐度的80%以上。腸桿菌科被認(rèn)為是各類腐敗干腌火腿中的優(yōu)勢(shì)菌群,特別是沙雷氏菌屬和變形桿菌屬(Proteus)[20-21]。Losantos等[14]從腐敗的塞拉諾和伊比利亞火腿中分離出液化沙雷氏菌(S. liquefaciens)和奇異變形桿菌(P. mirabilis),并認(rèn)為它們的蛋白水解能力可能是臭味形成的原因。類似地,觀察到腸桿菌科微生物與肌鈣蛋白和原肌球蛋白的強(qiáng)烈負(fù)相關(guān)[6]。
2.3 揮發(fā)性風(fēng)味化合物分析
表3顯示正常和腐敗盤縣火腿的揮發(fā)性風(fēng)味組成。共檢出60 種揮發(fā)性風(fēng)味物質(zhì),正常火腿39 種,腐敗火腿34 種。其中27 種為正常火腿獨(dú)有,21 種為腐敗火腿獨(dú)有。按種類將60 種風(fēng)味物質(zhì)分為醛類(23 種)、酮類(13 種)、醇類(6 種)、烴類(6 種)、含苯化合物(6 種)、酸類(2 種)、含硫化合物(2 種)及含氮化合物(2 種)。

表3 正常與腐敗盤縣火腿的揮發(fā)性風(fēng)味物質(zhì)種類和相對(duì)含量Table 3 Types and relative contents of volatile flavor compounds in normal and spoiled Panxian hams

續(xù)表3
為進(jìn)一步篩選正常和腐敗火腿之間的顯著鑒別因子,進(jìn)行LEFSe分析,閾值設(shè)置為LDA>4,結(jié)果如圖2所示。共鑒定出20 種具有顯著差別的鑒別因子,其中正常火腿12 種,腐敗火腿8 種。除酸類和含氮化合物外,其余6大類物質(zhì)在正常和腐敗火腿之間存在顯著差異。正常火腿含有豐富的醛類和醇類,包括壬醛、己醛、十六醛、辛醛、2-十一烯醛、十五醛、(E)-2-癸烯醛、庚醛和1-辛烯-3-醇;而高含量的烴類、酮類、含苯和含硫化合物是腐敗火腿的特征,特別是苯酚、苯乙醛、二甲基二硫醚、1-十五烯和3-甲基丁酸。

圖2 正常與腐敗火腿中的特征揮發(fā)性風(fēng)味化合物Fig. 2 Characteristic volatile flavor compounds in normal and spoiled Panxian hams
醛類物質(zhì)可以由脂質(zhì)氧化和氨基酸的Strecker反應(yīng)產(chǎn)生,對(duì)干腌火腿的整體風(fēng)味有重要貢獻(xiàn)。在正常火腿中,檢測到23 種醛,占風(fēng)味物質(zhì)總面積84.37%,而在腐敗火腿中僅檢出5 種,占總面積的6.05%。正常盤縣火腿中含量較大的醛為壬醛、己醛和十六醛。其中,壬醛和己醛是許多干腌火腿中最豐富的醛類物質(zhì),其低氣味閾值有助于增加甜味和青草香氣[22];而具有肉香味的十六醛是巴馬火腿[23]和伊斯特拉火腿[24]的典型特征。相反,腐敗火腿中的酮類物質(zhì)在種類和含量(11 種,14.75%)上均高于正常火腿(6 種,1.27%)。García等[25]報(bào)道了類似的結(jié)果,認(rèn)為伊比利亞火腿中酮類物質(zhì)的增加與腐敗微生物的活動(dòng)有關(guān)。另外,酮也可以通過游離脂肪酸的化學(xué)自動(dòng)氧化形成[26]。
醇類物質(zhì)的形成與脂質(zhì)氧化、氨基酸代謝、甲基酮還原和微生物繁殖密切相關(guān)。與之前的報(bào)道不同[27],醇類物質(zhì)在正常火腿中的含量高于腐敗火腿。醇類物質(zhì)的嗅聞閾值通常比醛酮類物質(zhì)低,但由花生四烯酸氧化形成的1-辛烯-3-醇經(jīng)常從干腌火腿中檢出,且具有較低的閾值,被認(rèn)為是盤縣火腿[19]、金華火腿[28]及如皋火腿[29]的特征風(fēng)味。在本研究中,1-辛烯-3-醇是正常火腿中最豐富的醇,也是正常與腐敗火腿之間的鑒別因子。
烴類物質(zhì)在種類和含量上的差異也有助于區(qū)分正常和腐敗盤縣火腿。正常和腐敗盤縣火腿中的烴類物質(zhì)分別占樣品風(fēng)味總面積的0.54%和13.51%。Martín等[27]在未變質(zhì)(3.98%)和變質(zhì)伊比利亞火腿(12%)中發(fā)現(xiàn)了類似的結(jié)果。通常,烴類化合物因其較高的閾值而對(duì)火腿風(fēng)味貢獻(xiàn)較小,但一些烯烴類物質(zhì)可能會(huì)影響整體風(fēng)味。1-十五烯是腐敗盤縣火腿中的特征化合物,占風(fēng)味總面積11.05%。據(jù)報(bào)道,它也是異味薏仁米中最主要的揮發(fā)性成分[30]。
含苯化合物是腐敗盤縣火腿中最主要的風(fēng)味成分,51.66%的峰面積由這類化合物代表。其中,含量較高的苯酚和苯乙醛是腐敗火腿的標(biāo)志性風(fēng)味。苯酚是一種具有特殊臭味的化合物,適量的苯酚似乎有助于發(fā)酵肉制品的風(fēng)味[31],但高含量的苯酚會(huì)影響金華火腿的風(fēng)味[32]。同時(shí),苯酚具有毒性,已被世界衛(wèi)生組織列入致癌物質(zhì)清單。苯乙醛是一種具有花香的氨基酸衍生物,經(jīng)常發(fā)現(xiàn)于國內(nèi)外干腌火腿中[33-34]。然而,高含量的苯乙醛已被認(rèn)為是伊比利亞火腿腐敗的特征之一[35]。
與之前研究結(jié)果一致[19],在正常盤縣火腿中未檢出含硫化合物。兩種含硫化合物(二甲基二硫醚和二甲基四硫化物)僅存在于腐敗火腿中,占風(fēng)味總面積的6.52%。硫化物代表燒焦和臭雞蛋的不愉快氣味,通常來自于微生物對(duì)含硫氨基酸的降解[36]。如圖2B所示,二甲基二硫醚被鑒定為腐敗盤縣火腿的特征風(fēng)味化合物。盡管二甲基二硫醚已被描述為金華和如皋火腿的主體風(fēng)味成分[28-29],但這類化合物在變質(zhì)伊比利亞火腿中的含量遠(yuǎn)高于未變質(zhì)火腿[25,35]。
3-甲基丁酸也是腐敗盤縣火腿的特征風(fēng)味。在伊比利亞火腿中,3-甲基丁酸的氣味被描述為像腳的、酸的和變質(zhì)火腿的味道,且僅在腐敗火腿中檢測到[35]。腐敗火腿中高含量的3-甲基丁酸可以通過以下兩方面解釋:1)3-甲基丁醛氧化形成3-甲基丁酸;2)腸桿菌科的高蛋白酶活性促進(jìn)游離氨基酸的釋放,氨基酸通過微生物發(fā)酵生成3-甲基丁酸。對(duì)于含氮化合物,四甲基吡嗪僅存在于正常火腿中,而N,N-二甲基-4-吡啶僅在腐敗火腿中檢測到。雖然有研究提出應(yīng)該將一些吡嗪類化合物視為檢測伊比利亞火腿初期腐敗的指標(biāo),但不包括四甲基吡嗪[27]。
2.4 環(huán)境因子對(duì)微生物群落的影響
物理化學(xué)性質(zhì)的改變顯著影響微生物群落的結(jié)構(gòu)和組成。合適的溫度、水分和鹽含量不僅有助于火腿獨(dú)特風(fēng)味的形成,還可以抑制有害微生物的生長繁殖[37]。因此,應(yīng)用RDA揭示優(yōu)勢(shì)菌屬與環(huán)境因子間的關(guān)系。銳角表示菌屬與環(huán)境因子之間呈正相關(guān),而鈍角表示呈負(fù)相關(guān),結(jié)果如圖3所示。水分含量、aw和pH值與未分類腸桿菌科微生物呈顯著正相關(guān),而與沙雷氏菌屬(Serratia)的正相關(guān)較弱。同時(shí),NaCl濃度顯示出對(duì)沙雷氏菌屬的強(qiáng)烈抑制,這種顯著負(fù)相關(guān)已在腐敗伊比利亞火腿中觀察到[6]。相反,NaCl濃度與其余優(yōu)勢(shì)菌屬呈正相關(guān),特別是色鹽桿菌(Chromohalobacter)和鹽單胞菌(Halomonas)。這兩個(gè)屬的大多數(shù)成員具有嗜鹽性,經(jīng)常從高鹽發(fā)酵食品中分離出來[38-39],具有蛋白酶和脂肪酶活性[40]。據(jù)報(bào)道,色鹽桿菌是韓國醬油品質(zhì)形成的關(guān)鍵[41],而鹽單胞菌的成員具有降解生物胺的能力[42]。正常火腿中最豐富的葡萄球菌與NaCl濃度的正相關(guān)較弱,但與水分含量、aw和pH值呈強(qiáng)烈負(fù)相關(guān)。類似地,最近一項(xiàng)研究揭示了肉類發(fā)酵過程中pH值對(duì)葡萄球菌群落結(jié)構(gòu)的顯著影響[43]。

圖3 優(yōu)勢(shì)細(xì)菌屬與環(huán)境因子之間的RDAFig. 3 RDA of the relationship between dominant bacterial genera and environmental factors
2.5 優(yōu)勢(shì)菌屬與揮發(fā)性風(fēng)味化合物之間的網(wǎng)絡(luò)關(guān)系
干腌火腿加工過程中伴隨著一系列的生化和酶促反應(yīng),包括蛋白質(zhì)水解、脂質(zhì)氧化、糖原分解和美拉德反應(yīng)等,這些反應(yīng)對(duì)火腿風(fēng)味的形成至關(guān)重要。眾所周知,肌肉內(nèi)源酶,特別是組織蛋白酶主導(dǎo)這些反應(yīng),而微生物的酶促作用也有助于風(fēng)味化合物的產(chǎn)生[44]。據(jù)報(bào)道,微生物或未知來源的揮發(fā)性風(fēng)味物質(zhì)占伊比利亞火腿總揮發(fā)性化合物的5.7%[29]和6.9%[45]。另外,郇延軍等[46]認(rèn)為微生物是影響不同等級(jí)金華火腿風(fēng)味的關(guān)鍵因素。因此,基于Pearson相關(guān)系數(shù)構(gòu)建優(yōu)勢(shì)菌屬與揮發(fā)性風(fēng)味物質(zhì)之間的相關(guān)性網(wǎng)絡(luò)。如圖4所示,代表正常火腿的7 種細(xì)菌屬和主導(dǎo)腐敗火腿的3 種細(xì)菌屬構(gòu)成兩個(gè)單獨(dú)的網(wǎng)絡(luò),分別促進(jìn)32 種和21 種風(fēng)味化合物的產(chǎn)生。與圖2結(jié)果一致,正常火腿以醛類和醇類物質(zhì)為主,而腐敗火腿以酮類、烴類、含苯和含硫化合物為代表。然而,在圖4A中觀察到苯甲醛和3 種酮的存在。雖然腐敗火腿中更高含量的苯甲醛已被報(bào)道[27],但這種具有苦杏仁和橡子氣味的化合物在干腌火腿中經(jīng)常發(fā)現(xiàn),并且被認(rèn)為是伊比利亞火腿的氣味活性成分[47]。如圖4B所示,2 種未知的腸桿菌科微生物與腐敗火腿的5 種特征風(fēng)味(苯酚、苯乙醛、1-十五烯、3-甲基丁酸和二甲基二硫醚)密切相關(guān),而沙雷氏菌屬只與苯酚、苯乙醛和3-甲基丁酸的產(chǎn)生有關(guān)。同時(shí),沙雷氏菌屬僅占腐敗火腿總豐度的8.85%(圖1B)。因此,盡管許多研究認(rèn)為沙雷氏菌屬是造成干腌火腿腐敗的主要腸桿菌科微生物[6,21],但它可能并不是導(dǎo)致盤縣火腿腐敗的關(guān)鍵因素。造成這種差異的原因之一可能是此前的研究均采用培養(yǎng)法,大部分微生物無法培養(yǎng)的局限性導(dǎo)致研究者得出值得商榷的結(jié)論。有必要應(yīng)用準(zhǔn)確性更高、測序深度更深的宏基因組進(jìn)行下一步研究。

圖4 優(yōu)勢(shì)細(xì)菌屬與揮發(fā)性風(fēng)味化合物的相關(guān)性網(wǎng)絡(luò)Fig. 4 Correlation network between dominant bacterial genera and volatile flavor compounds
3 結(jié) 論
本研究揭示了正常與腐敗盤縣火腿在理化特性、微生物多樣性和揮發(fā)性風(fēng)味物質(zhì)之間的差異,確定了與腐敗火腿高度相關(guān)的幾種特征,即高比例的腸桿菌科和高含量的苯酚、苯乙醛、1-十五烯、3-甲基丁酸及二甲基二硫醚,這對(duì)預(yù)防和減少盤縣火腿的腐敗具有重要意義。然而,目前基于16S rRNA的高通量測序未能準(zhǔn)確鑒定出主導(dǎo)盤縣火腿腐敗的微生物。因此,后續(xù)研究中,應(yīng)該使用基于第3代測序的宏基因組進(jìn)一步在屬水平和/或物種水平上表征這些微生物,并利用多組學(xué)技術(shù)(如蛋白組學(xué)、轉(zhuǎn)錄組學(xué)和代謝組學(xué))驗(yàn)證它們與揮發(fā)性風(fēng)味物質(zhì)之間的關(guān)系。
