中溫α-淀粉酶的分子進化及酶學性質
2021-06-30 14:57:02馬銀鳳申培立王彩喆牛丹丹田康明王正祥
食品與發酵工業 2021年11期
馬銀鳳,申培立,王彩喆,牛丹丹,田康明,王正祥*
1(天津科技大學 化工與材料學院,天津,300457) 2(天津科技大學 生物工程學院,天津,300457)
解淀粉芽胞桿菌α-淀粉酶(Bacillusamyloliquefaciensα-amylase,BAA)可以隨機水解直鏈淀粉或支鏈淀粉分子中的α-1,4糖苷鍵,生成麥芽糊精和少量葡萄糖,是一種內切型液化酶,最適作用溫度為55 ℃、最適作用pH為6~7,是一種典型的鈣離子依賴型淀粉液化酶[1-2]。BAA在淀粉液化、織物退漿和食品加工等工業中具有重要應用價值,是工業上產量最大、應用領域最廣的中溫型α-淀粉酶[3-4]。
BAA作為我國開發應用較早的工業酶制劑之一,早在19世紀60年代就開始了工業化生產,但發酵水平一直停滯在300~500 U/mL,且酶制劑產品質量不穩定、貨架期太短(3個月左右)。2009年前后,經過分子育種技術,在改善BAA酶制劑質量和貨架期的同時,BAA發酵生產水平提高至1 500 U/mL以上,由此確立了BAA生產的世界領先地位[5]。但是,針對BAA工業應用屬性的研究,一直以來未有很大的突破。現有針對BAA的分子進化研究,主要包括:LEE等[6]對BAA編碼基因進行隨機突變,發現233位的氨基酸殘基與BAA的酶學性能有關;CORNELIUS等[7]通過易錯PCR與DNA體外隨機重排技術(DNA shuffling)對BAA編碼基因進行改造,得到水解活力提高8倍的突變體BAA29,比酶活性由15 U/mg提高至140 U/mg;WU等[8]通過易錯PCR對BAA編碼基因進行隨機突變,得到Ca2+依賴性降低的突變體Q264S;林碧瑜[9]發現BAA第28位氨基酸由天冬氨酸突變為谷氨酸可以提高比酶活性,與野生型BAA相比突變體的比酶活性提高了約30%。
BAA工業應用屬性有待改善的主要方面:1)酶活性的鈣離子依賴型,即如何降低其在實際工業應用中氯化鈣或碳酸鈣等鈣鹽的補加;2)如何適度提高其最適作用溫度,有利于淀粉液化和加工工藝的實施。為此,本文在前期研究積累[9]的基礎上,進一步對其特定氨基酸殘基進行突變并解析酶學性質,研究結果對后續進一步提升BAA的應用具有顯著價值。
1 材料與方法
1.1 材料
1.1.1 菌種與質粒
含有BAA突變體2M6表達質粒的Bacillussubtilis2M6-A1為本實驗室前期構建與保藏;B.subtilis1A717為本實驗室保藏,用于質粒構建;載體pHY-WZX為本實驗室前期構建[10],用于BAA及其突變體的表達;BacilluslicheniformisZ902(Δamy)[11]為酶高效表達宿主細胞,由本實驗室前期通過分子改造B.licheniformisCBBD302[12]獲得。
1.1.2 酶與試劑
中溫α-淀粉酶工業產品,江蘇銳陽生物科技有限公司;Pyrobest DNA聚合酶,Takara公司;限制性內切酶XbaI、SmaI、T4DNA連接酶和DNA分子量標準,上海生工生物工程股份有限公司;瓊脂糖凝膠DNA回收試劑盒和小量DNA產物純化試劑盒,北京莊盟生物科技有限公司,其他試劑除特殊要求外均為國產分析純。
1.1.3 培養基
LB培養基(g/L):酵母浸出物5,蛋白胨10,氯化鈉10;固體培養基添加20 g/L瓊脂。必要時在培養基中添加終質量濃度為20 μg/mL的硫酸卡那霉素。
產酶鑒定培養基:在固體LB培養基的基礎上,加入1%可溶性淀粉[13]。
發酵培養基(g/L):CaCl20.3,(NH4)2SO43,棉籽粉20,α-乳糖20,KH2PO410,K2HPO420,pH 7.2[13]。
1.2 實驗方法
1.2.1 常規分子克隆操作
質粒DNA的提取、酶切、轉化子篩選等均按照實驗室常規方法進行[14];枯草桿菌與地衣芽胞桿菌的遺傳轉化按照文獻進行[15-16];DNA純化按照試劑盒說明書進行。
1.2.2 定點突變
以突變體2M6編碼基因為模板,按照文獻方法進行定點突變[5]。具體步驟為:以baa-28a(5′-CGATAAATGTTCCGCATCATTCTG-3′)和baa-28b(5′-CAGAATGATGCGGAACATTTATCG-3′)為突變引物,引入突變位點D28E,與兩端引物BAA-1(5′-TACTCTAGAGTAAATGGCACGCTGATGCAGTAT-3′)和BAA-2(5′-TTTCTGAACATAAATGGAGCAGG-3′)組合,通過PCR擴增獲得上下游兩段DNA片段;再以此兩段DNA片段為模板,以BAA-1和BAA-2為引物,將上下游兩段DNA片段融合為突變體E2M6全長基因。采用Sanger核苷酸序列測定法確認突變基因序列。
1.2.3 突變體的表達、制備與純化
將突變體全長編碼DNA片段純化后經XbaI酶切,并克隆入表達載體pHY-WZX的XbaI/SmaI位點,轉化入枯草桿菌1A717構建重組質粒pHY-E2M6,正確構建的質粒轉化入地衣芽胞桿菌Z902中,獲得產酶重組菌地衣芽胞桿菌Z-E2M6。
將Z-E2M6接種入盛有50 mL發酵培養基的250 mL搖瓶,37 ℃、220 r/min條件下培養120 h;發酵結束后離心收集上清液,即為突變體粗酶液。酶液經飽和度為30%~40%的硫酸銨沉淀,再經凝膠層析柱Superdex-G75進一步純化。純化蛋白經SDS-PAGE分析[17],蛋白質濃度用Bradford法測定[18]。
1.2.4 酶活測定
中溫α-淀粉酶酶活力按照GB 1886.174—2016《食品安全國家標準 食品添加劑 食品工業用酶制劑》進行測定[19]。酶活性定義為:1 mL液體酶在60 ℃、pH 6.0條件下,1 h液化1 g可溶性淀粉即為1個酶活力單位,以U/mL表示。
1.2.5 酶學性質分析
1.2.5.1 最適作用溫度及熱穩定性
將酶樣在不同溫度(30~90 ℃)下測定酶活力,最高酶活力值以100%計,計算其不同溫度下的相對酶活力,確定最適作用溫度。將酶樣用pH 6.0緩沖液進行適當稀釋,在不同溫度(50、55、60、65 ℃)下保溫1 h,間隔10 min取樣,測定殘余酶活力,未經熱處理的酶活力值以100%計,計算處理后酶樣的相對酶活力。
1.2.5.2 最適作用pH及pH穩定性
將酶樣用不同pH(4~8)緩沖液進行稀釋,并用對應pH緩沖液測定酶活力,最高酶活力值以100%計,計算其不同pH條件下的相對酶活力,確定最適作用pH。將酶樣與不同pH值緩沖液以1∶10(體積比)混合,37 ℃靜置1 h,測定酶樣的殘余酶活力值。未經處理的酶活力值以100%計,計算處理后酶樣的相對酶活力。
1.2.5.3 金屬離子及化學試劑對酶活的力影響
在反應體系中加入不同金屬離子和化學試劑,終濃度為1 mmol/L或5 mmol/L,在酶的最適反應條件下測定樣品酶活力,未經處理的酶活力值以100%計。
1.2.5.4 酶動力學參數的測定
將酶樣用最適作用pH緩沖液適當稀釋后,以不同質量濃度(0.4、1、2、4,10和20 mg/mL)可溶性淀粉溶液為底物,在最適反應條件下按照實驗室常規方法[9]進行測定,并以雙倒數法(Lineweaver-Burk法)作圖,計算酶的動力學參數。
2 實驗結果
2.1 突變體的構建與制備
以突變體2M6編碼基因為模板,特異性引物序列介導BAA編碼基因的定點突變,獲得突變體E2M6全長編碼基因。將此基因酶切后克隆入表達載體pHY-WZX中,獲得了重組表達載體pHY-E2M6。進一步將此表達質粒轉化入地衣芽胞桿菌Z902中,獲得產酶重組菌地衣芽胞桿菌Z-E2M6。
經搖瓶發酵制備酶液,并經硫酸銨沉淀和凝膠過濾色譜進行純化,獲得了純化的BAA突變體E2M6,經SDS-PAGE電泳鑒定,呈現單一純蛋白條帶(圖1),分子質量為58 kDa,與理論大小相符。
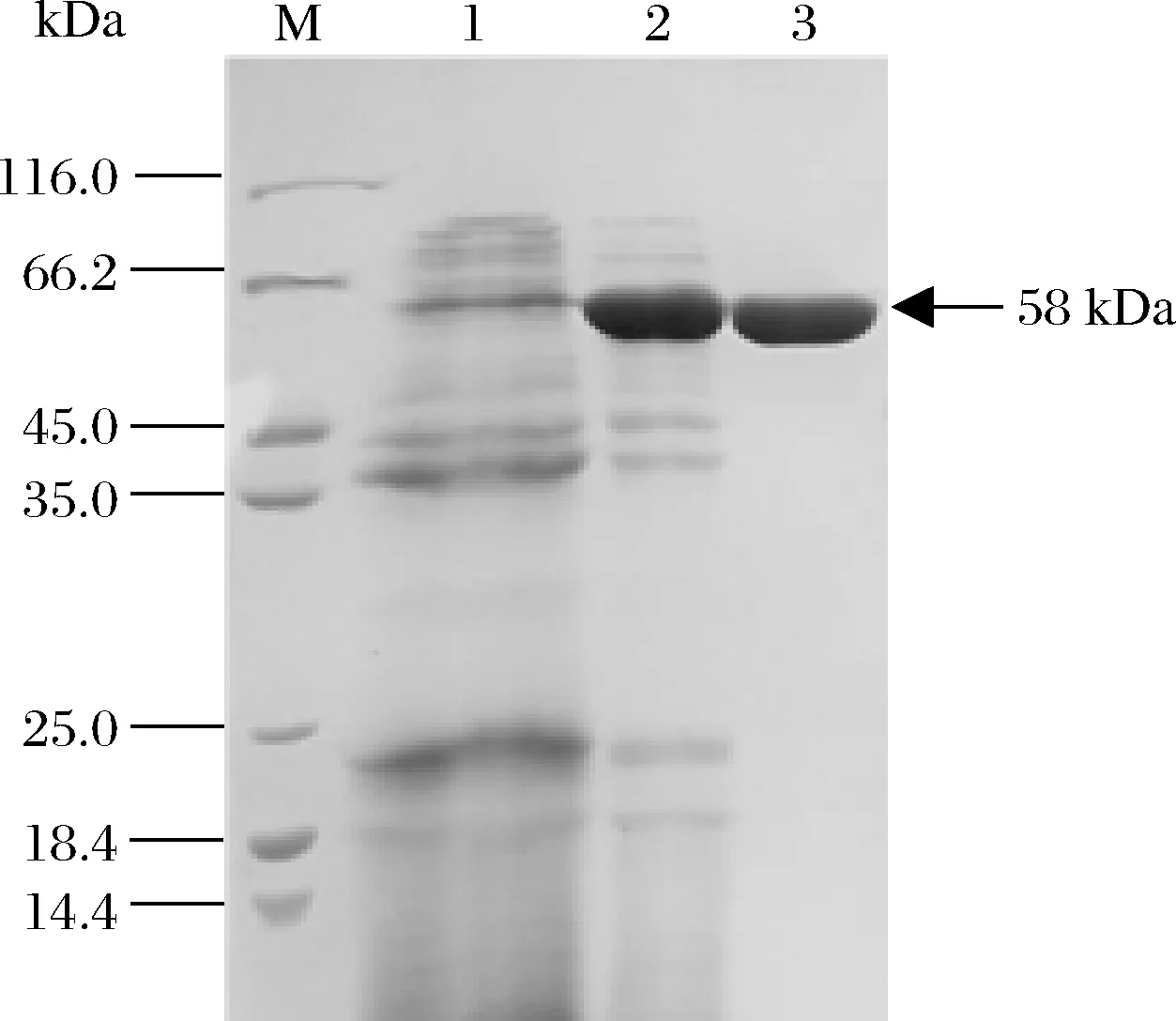
M-蛋白分子量標準;1-B.licheniformis Z902發酵酶液;2-E2M6粗酶液;3-純化后的E2M6圖1 純化突變體E2M6的SDS-PAGE圖譜Fig.1 The SDS-PAGE profile of mutant E2M6
2.2 突變體的酶學性質
2.2.1 最適作用溫度與最適作用pH
由圖2-a可知,突變體E2M6的最適作用溫度為60 ℃,60~70 ℃之間維持較高活性,突變體的最適酶活較野生型BAA提高了至少5 ℃;在30~60 ℃溫度范圍內,突變體E2M6與BAA的變化趨勢大致相同;當溫度升至80 ℃時,野生型BAA的酶活力明顯下降(小于40%),突變體E2M6仍保留70%以上的酶活力;當反應溫度升至90 ℃時,突變體E2M6仍保留20%以上的酶活力,而BAA僅殘余約10%的酶活力。可見,對BAA的突變拓寬了BAA的作用溫度范圍。

a-溫度;b-pH圖2 突變體E2M6的最適作用溫度與最適作用pHFig.2 Optimum temperature and pH optimal of mutant E2M6
在60 ℃條件下,測定不同pH(4.0~8.0)條件下的酶活力,結果如圖2-b所示。突變體E2M6與BAA的最適作用pH相同,都為pH 6.0,且兩者在不同pH條件下的酶活力大致相同,在pH 5.0~7.0范圍內皆具有較高活性。
2.2.2 熱穩定性與pH穩定性
BAA突變體的熱穩定性和pH穩定性如圖3所示。突變體在50 ℃下保溫1 h后,E2M6與BAA分別保留80%、60%以上的酶活力;55 ℃保溫1 h后E2M6與BAA分別保留60%、40%以上酶活力;60 ℃保溫1 h后E2M6與BAA分別保留50%、30%以上的酶活力;65 ℃保溫1 h后E2M6與BAA分別保留40%、15%以上的酶活力(圖3-a)。綜上,E2M6熱穩定性較BAA有所提升。
突變體E2M6與BAA的pH穩定性趨勢大致相同。當pH在6.0~9.0范圍內,pH的升高對BAA及E2M6酶活力影響較小,相對酶活力保持在80%以上(圖3-b)。
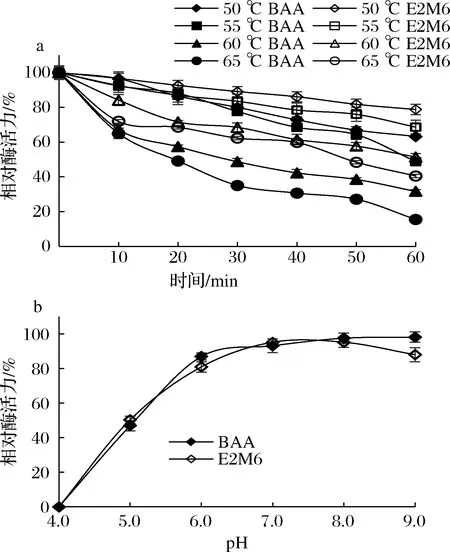
a-熱穩定性;b-pH穩定性圖3 突變體E2M6的熱穩定性與pH穩定性Fig.3 The thermostability and pH stability of the mutant E2M6
2.2.3 金屬離子和化學試劑對酶活性的影響
不同金屬離子及化學試劑對突變體酶活性的影響結果如表1。與BAA相比,突變體E2M6酶活力對Ca2+、Mg2+、Na+等金屬離子的依賴性降低;SDS對BAA及突變體E2M6酶活均有抑制作用,但與BAA相比E2M6對SDS更敏感。
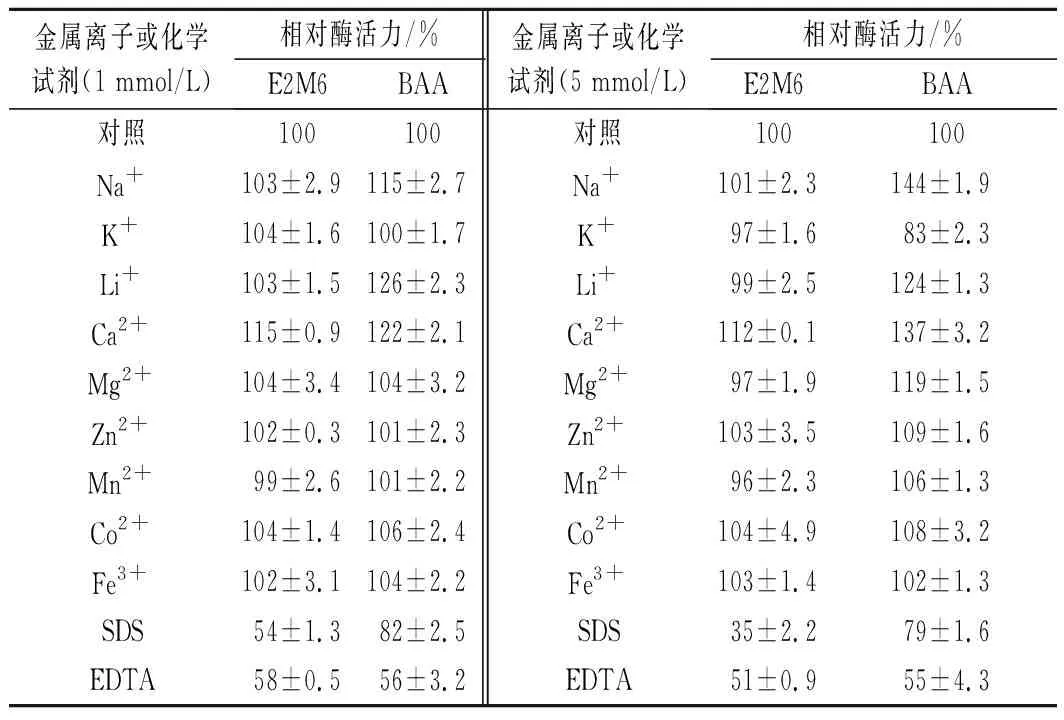
表1 不同金屬離子和化學試劑對突變體E2M6的影響Table 1 Effect of different metal ions and chemicals on E2M6
2.2.4 突變體E2M6動力學特征
測定突變體E2M6的比酶活力數據與動力學常數,結果匯總于表2。突變體E2M6比酶活力為185 U/mg,較BAA提高了約66%;突變體E2M6的底物親和力較BAA顯著提高,最大反應速度雖有所降低,但突變體E2M6的淀粉水解效率(κcat/Km)有所提升,較BAA提高了1.75倍。

表2 突變體E2M6的酶促學動力學特征Table 2 The kinetic parameters of the mutant E2M6
3 結論
通過研究,本文獲得了BAA工業應用性能顯著提升的突變體,其活力的鈣離子依賴性顯著降低,最適作用溫度有所提高,更適合淀粉加工工業應用需求。
