致病性大腸桿菌全基因組測序及毒力基因分析
2021-06-30 00:59:18王嘉福冉雪琴黃世會
工業(yè)微生物 2021年3期
關鍵詞:小鼠
周 倩,王嘉福*,冉雪琴,牛 熙,黃世會
1.貴州大學生命科學學院,貴州貴陽 550025;2.貴州大學動物科學學院,貴州貴陽 550025
大腸桿菌存在于人和動物腸道中[1],某些大腸桿菌是導致仔豬腹瀉的主要病原菌之一[2],其導致的疾病發(fā)病率高、發(fā)病急、死亡率高、涉及范圍廣,給養(yǎng)豬業(yè)造成巨大經(jīng)濟損失,嚴重影響了養(yǎng)豬業(yè)的發(fā)展[3]。然而由該菌引發(fā)的疾病目前缺少有效的治療手段,臨床上主要以母體抗原、抗血清、抗生素等為主,容易產(chǎn)生耐藥性、藥物殘留等問題[4-5]。
大腸桿菌通過黏附素黏附于斷奶仔豬小腸上皮細胞,就定居在小腸上并開始大量繁殖并產(chǎn)生毒素,產(chǎn)生的毒素被吸收后,引起胃腸道血管、皮下組織和腦血管的損傷,從而使宿主發(fā)生一系列疾病[6]。致病性大腸桿菌侵襲宿主組織引起其發(fā)病的過程是由多種毒力因子相互協(xié)調(diào)而發(fā)揮作用,大腸桿菌的主要毒力因子包括毒素、黏附素、外膜蛋白、鐵轉運系統(tǒng)和吸收系統(tǒng)等[7]。為研究致病性大腸桿菌致病力差異的原因,采用二代高通量測序技術對7株不同致病力的大腸桿菌進行全基因組測序,分析毒力基因的差異,以期解析大腸桿菌的致病因子和致病機制。
1 材料與方法
1.1 材料
菌株P211、P555、P32、P444和P111由本實驗室從貴州某豬場腹瀉仔豬糞樣中分離,菌株S10670、E24190購自中國工業(yè)微生物菌種保藏管理中心。40只SPF級小鼠購自北京斯貝福生物技術有限公司。
1.2 菌株純化鑒定
使用EMB、MAC和LB固體培養(yǎng)基對7菌株進行純化,隨后對菌株進行革蘭氏染色鏡檢和生理生化鑒定,再用細菌16S rDNA基因通用引物27F和1 492R進行擴增(表1),擴增產(chǎn)物進行Sanger測序,用NCBI網(wǎng)站對測序產(chǎn)物進行Blast同源比對,對菌株進行分子系統(tǒng)學鑒定。

表1 細菌16S rDNA擴增引物
1.3 小鼠致病性試驗
挑取單個純化后的7株菌株于LB液體培養(yǎng)基中培養(yǎng)至濃度為1×108CFU/mL的菌懸液,將40只小鼠隨機分為8組,7個實驗組和一個對照組,每組5只小鼠,實驗組小鼠的給藥途徑、給藥方法和給藥劑量參照《藥理實驗方法學》的動物實驗技術進行[8],按照10 g/0.2 mL 的劑量腹腔注射菌懸液,對照組則按照10 g/0.2 mL 的劑量腹腔注射生理鹽水。正常飼喂,自由飲水,感染后觀察48 h,記錄小鼠臨床癥狀和死亡時間,死亡小鼠及時剖檢觀察內(nèi)臟組織病變并制作病理切片,觀察病理變化。
1.4 測序樣品
挑取單個純化鑒定后的菌株用LB液體培養(yǎng)基中培養(yǎng)于37 ℃、160 r/min培養(yǎng)過夜后收集菌體,使用細菌基因組DNA提取試劑盒提取基因組,經(jīng)酶標儀及1%的瓊脂糖凝膠電泳檢測其濃度、純度均達到基因組DNA 的測序要求后進行全基因組測序。測序工作由華大深圳股份有限公司完成,測序平臺為DNBSEQ。
1.5 基因文庫構建
將基因組DNA的大片段隨機打斷獲得200 bp~400 bp的DNA片段,然后修復雙鏈DNA末端,并在3'末端加上A堿基,配制接頭連接反應體系使接頭與DNA連接,擴增連接產(chǎn)物并純化回收,將PCR產(chǎn)物變?yōu)閱捂満螅渲骗h(huán)化反應體系,得到單鏈環(huán)形產(chǎn)物,消化掉未被環(huán)化的線性DNA分子后,即得到最終的文庫。
1.6 全基因組測序與基因組組裝
檢測合格文庫安排上機測序(DNBSEQ):單鏈環(huán)狀DNA分子通過滾環(huán)復制,形成一個包含300多個拷貝的DNA納米球(DNB)。將得到的DNBs采用高密度DNA納米芯片技術,加到芯片上的網(wǎng)狀小孔內(nèi)。通過聯(lián)合探針錨定聚合技術(cPAS)進行測序。得到測序原始數(shù)據(jù)后,對raw reads過濾質(zhì)控,去除低質(zhì)量reads以及adapter和duplication污染后,從而得到高質(zhì)量Clean Reads后,采用SOAPdenovo[9]組裝軟件進行序列組裝,通過不斷優(yōu)化參數(shù)Kmer值為15獲得最好的組裝結果。
1.7 基因預測及注釋
采用Glimmer 3.02[10]進行組裝結果的基因預測,分別采用RNAmmer-1.2[11]和tRNAscan-SE[12]軟件對基因組中包含的rRNA和tRNA進行預測。將預測基因的氨基酸序列與eggNOG(http://eggnog-mapper.embl.de/)[13]數(shù)據(jù)庫和VFDB(https://card.mcmaster.ca)[14]數(shù)據(jù)庫進行BLAST比對,將得到的基因進行功能注釋,獲得COG注釋信息和菌株毒力基因,并對COG注釋結果進行統(tǒng)計和分類,通過VFBD數(shù)據(jù)庫注釋到的毒力因子,分析不同致病力菌株的毒力基因差異。
2 結果
2.1 菌株的鑒定
2.1.1菌落形態(tài)特征
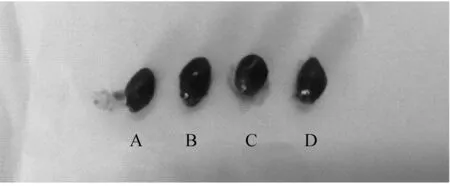
7株菌株在MAC瓊脂平板上形成圓形、表面光滑、濕潤的粉紅色菌落(圖1A);在EMB瓊脂培養(yǎng)基上形成表面光滑、凸起、帶黑色金屬光澤的菌落(圖1B);在LB固體培養(yǎng)基上形成圓形、表面光滑、半透明的乳白色菌落(圖1C);革蘭氏染色,鏡檢可見紅色短桿菌、中等大小、兩頭鈍圓、呈單個或多數(shù)散在排列(圖1D),鑒定結果均與大腸桿菌形態(tài)特征相符。
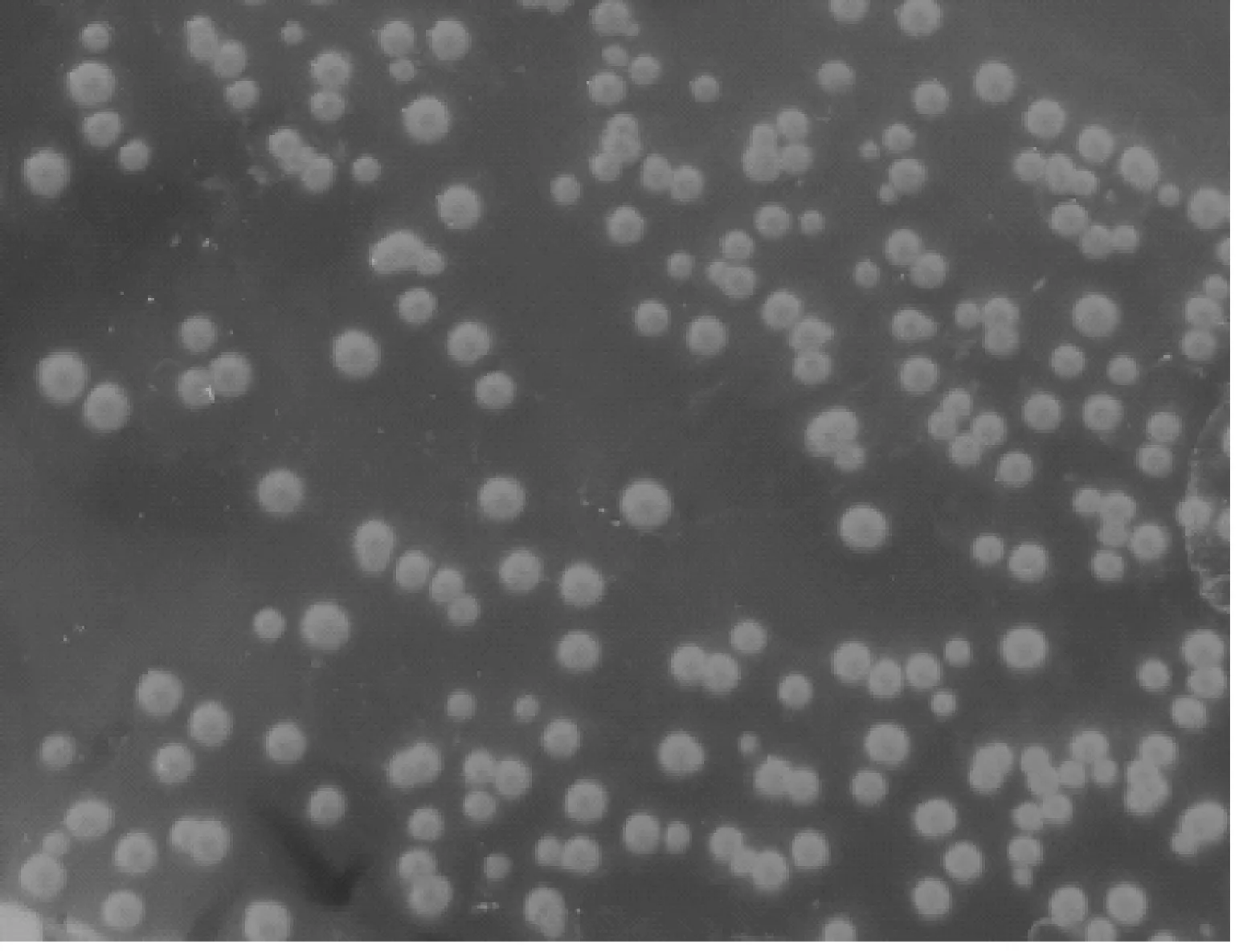
A
2.1.2生理生化鑒定
7株菌株均能發(fā)酵葡萄糖、甘露醇、乳糖、麥芽糖,尿素、蛋白胨水、乙酰甲基甲醇(V-P)、Koser肉湯、氧化酶、硫化氫試驗陰性,甲基紅試驗(MR)陽性,具有動力,結果顯示該7株菌與大腸桿菌具有相似的生理生化特性(表2)。
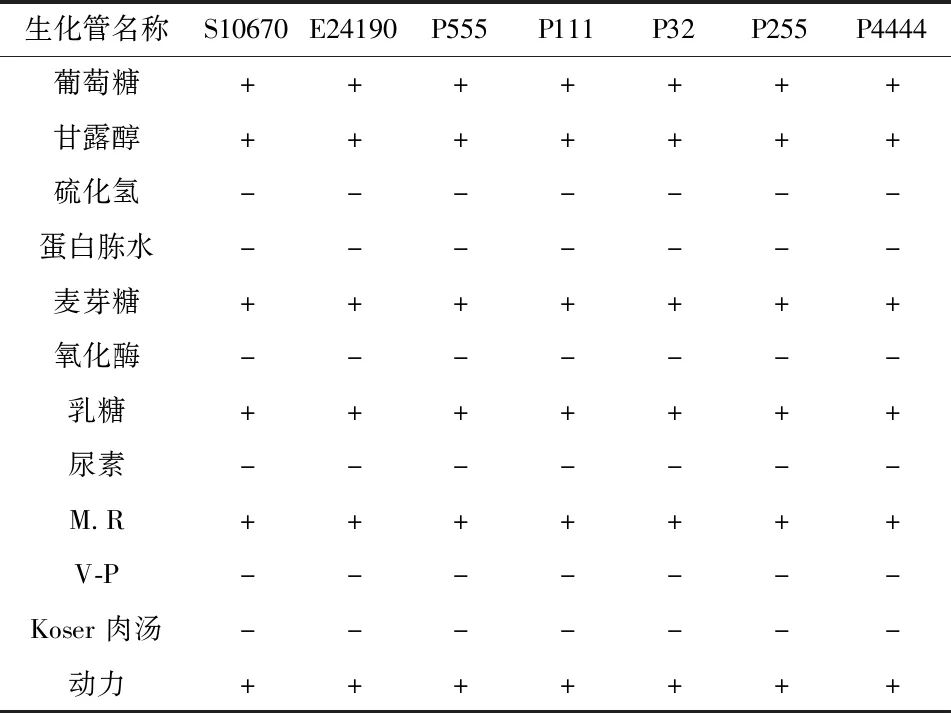
表2 7株菌株生理生化鑒定結果
2.1.3分子生物學鑒定
菌株16S rDNA基因經(jīng)PCR擴增獲得1 500 bp的片段,與預期相符(圖2),將16S r DNA基因序列與NCBI核酸數(shù)據(jù)庫比對,7株菌株與已知大腸桿菌的相似性最高,均在99.5%以上,將7株菌株鑒定為大腸桿菌。
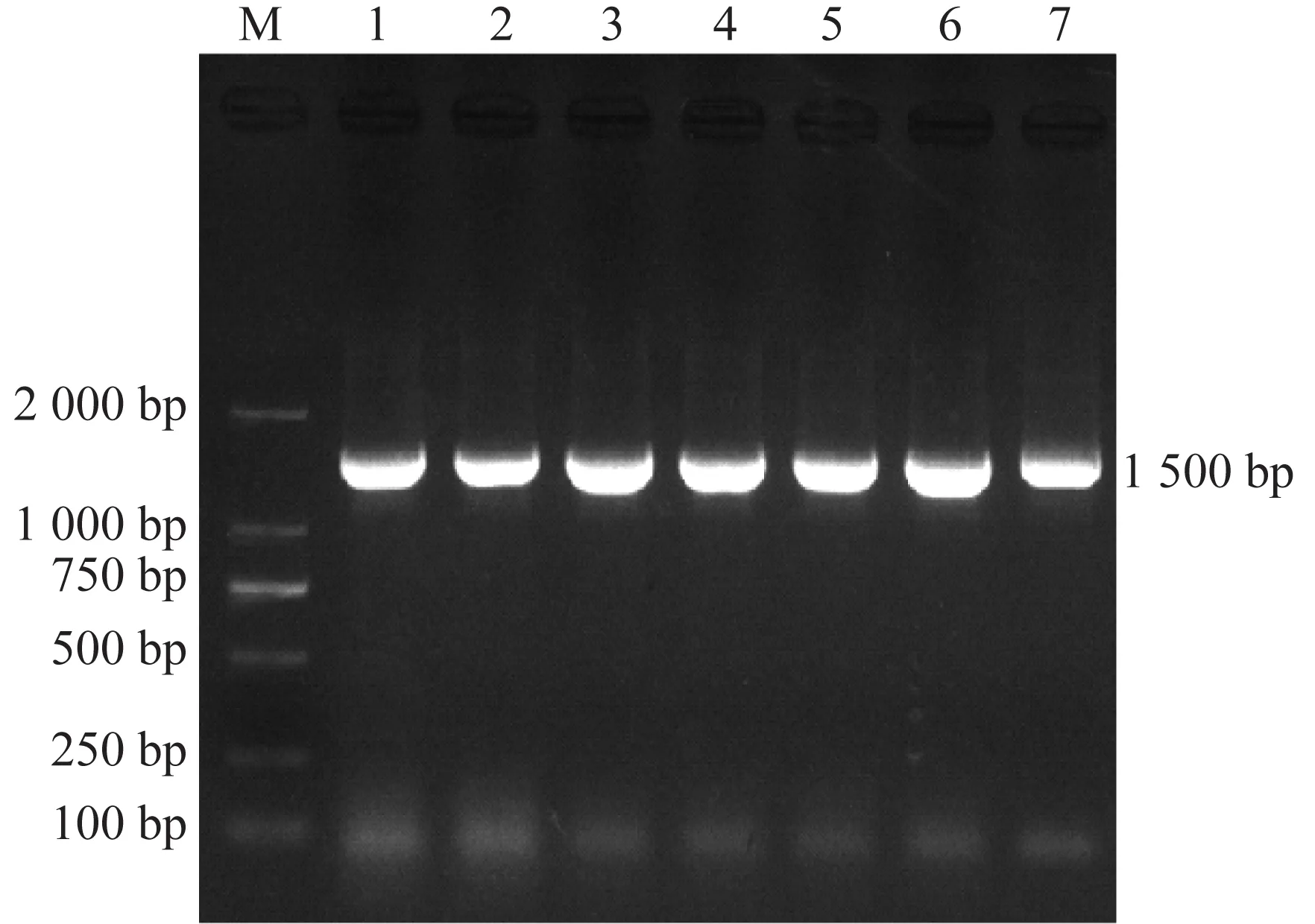
注:M:D 2 000 DNA marker ;1.S40670;2.E24190;3.P211;4.P555;5.P32;6.P444;7.P111圖2 16S rDNA基因擴增結果
2.2 小鼠致病性試驗
7株菌株用相同濃度(1×108CFU/mL)的菌懸液對小鼠進行感染試驗,結果顯示(表3),48 h后S10670和E24190實驗組小鼠死亡5只,病死率為100%,P211和P555實驗組小鼠死亡2只,病死率為40%,P32、P444和P111試驗組小鼠死亡1只,病死率為20%,對照組小鼠全部存活。因此,將菌株S10670和E24190歸為強毒株,P211和P555定為中等毒株,P32、P444和P32為弱毒株。感染后的小鼠出現(xiàn)背毛聳亂無光澤,進食少,活動緩慢,精神萎靡不活躍,倦怠、嗜睡,并且扎堆出現(xiàn),對外界刺激遲鈍,其中S10670和E24190組臨床現(xiàn)象最嚴重,重度腹瀉,接種菌株S10670的小鼠還有眼角包裹著眼屎,眼部出現(xiàn)水腫現(xiàn)象;P211和P555臨床現(xiàn)象次之,P32、P444和P111臨床現(xiàn)象最弱。
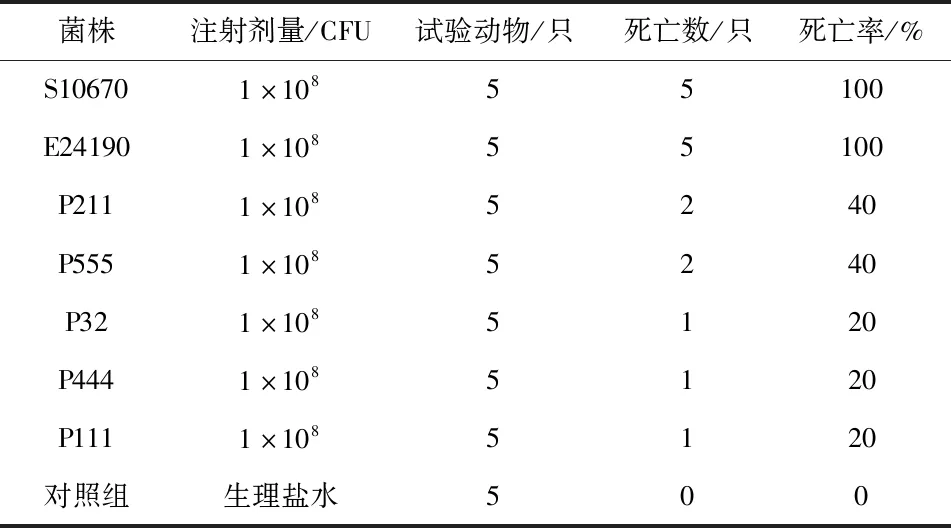
表3 小鼠致病性試驗結果
2.3 剖檢和組織病理學觀察
對死亡小鼠進行剖解后(圖3)觀察小組內(nèi)臟組織,與對照組相比實驗組小鼠內(nèi)臟組織出現(xiàn)不同程度的病變,表現(xiàn)為心臟、肝臟、脾臟、肺臟、腎臟腫大充血,顏色變深;腸道變薄,內(nèi)有大量內(nèi)容物,且味道惡臭。病理切片顯示(圖4),實驗組小鼠心臟組織心肌纖維紊亂、排列不整齊,部分出現(xiàn)斷裂崩解伴有出血;肝臟組織出現(xiàn)充血,細胞腫大,局部呈紅色塊狀,組織界限模糊;脾臟組織出現(xiàn)充血;肺臟組織部分輪廓不清晰,出血,肺泡腔內(nèi)有紅細胞,肺泡壁增厚;腎臟組織出現(xiàn)充血,腎小球結構不明顯;腸上皮細胞受損、腫大,腸黏膜壞死,腸絨毛脫落。其中組織病變和病理現(xiàn)象強毒株病變最嚴重,中等毒株次之,弱毒株最不明顯。
心
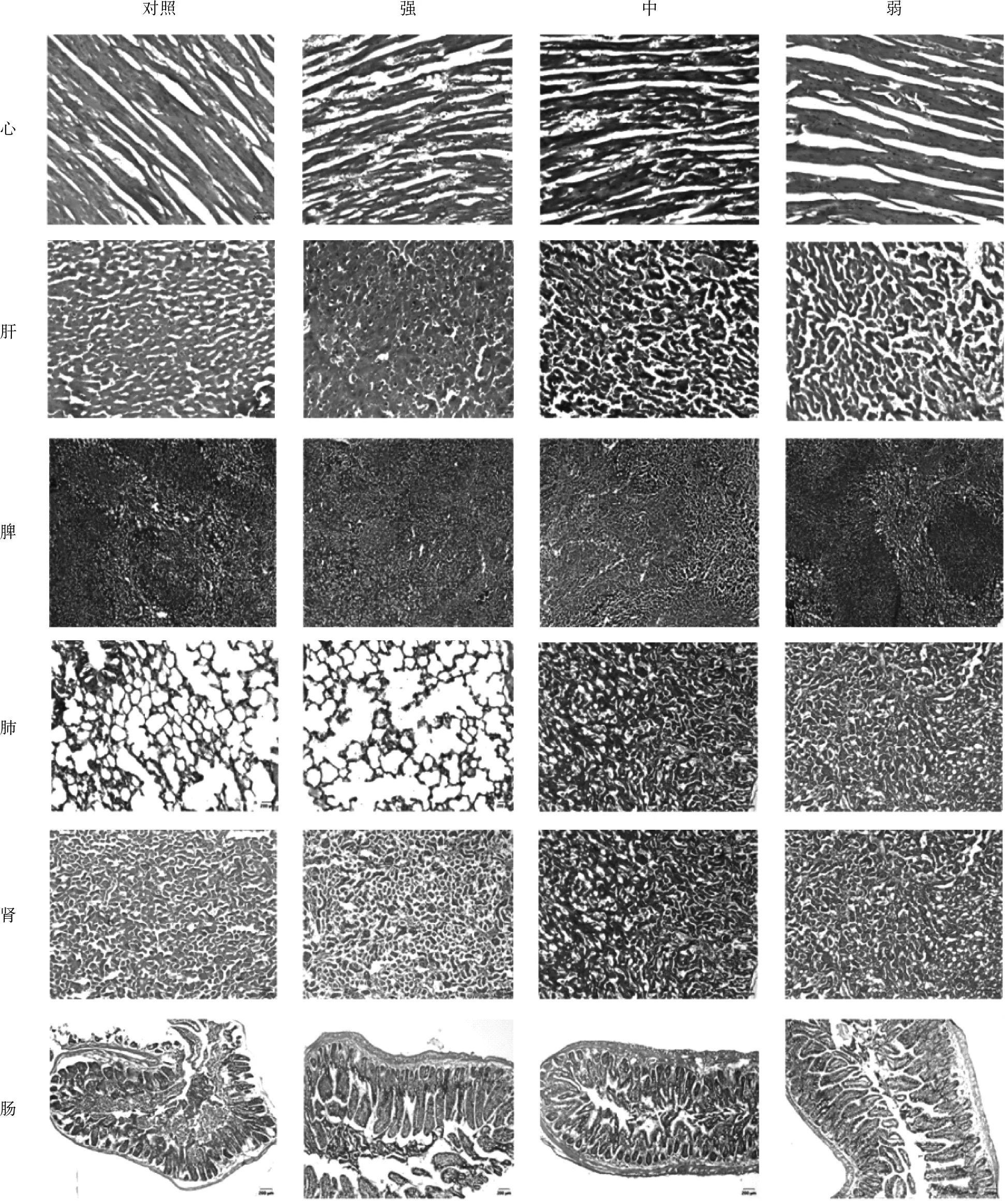
圖4 組織切片圖
2.4 基因組組裝
對測序數(shù)據(jù)進行質(zhì)量評估和組裝,由結果可得菌株S10670、E24190、P211、P555、P32、P444和P111組裝后得基因組大小依次為5.3 Mb、5.1 Mb、4.7 Mb、4.8 Mb、4.9 Mb、4.8 Mb和4.6 Mb,GC含量均在50%左右,基因組大小和GC含量均在大腸桿菌基因組大小和GC含量的正常范圍,其它詳細信息見表4,組裝結果可知菌株S10670的基因組最大。
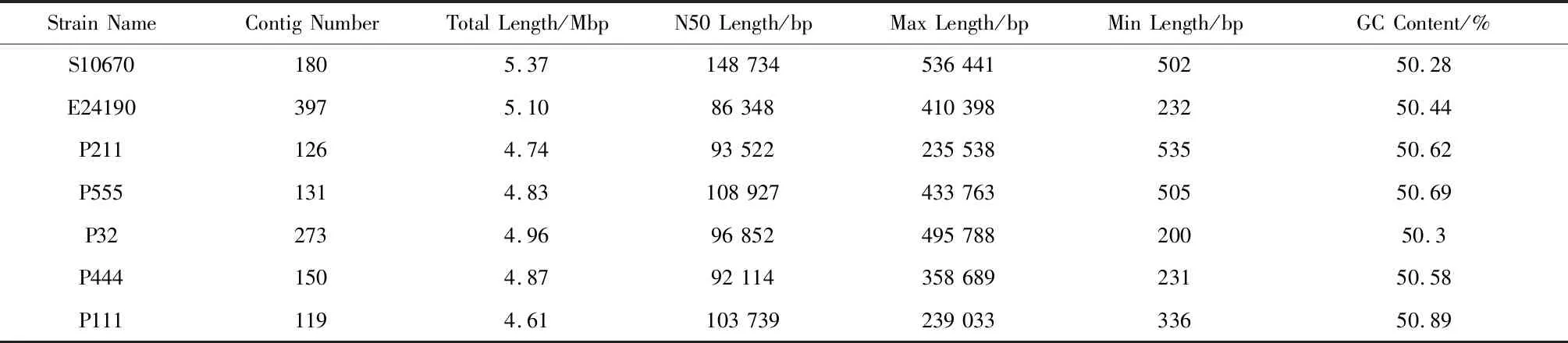
表4 基因組組裝結果
2.5 基因預測
對組裝好的序列進行基因組成分析,菌株S10670、E24190、P211、P555、P32、P444和P111采用Glimmer軟件預測基因數(shù)量分別為5 124、4 936、4 563、4 601、4 762、4 724和4 376,用rRNAmmer1.2軟件預測出的rRNA數(shù)量依次為9個、9個、5個、5個、3個、3個和7個,tRNAscan軟件預測出的tRNA數(shù)量依次為85個、74個、75個、66個、77個、70個和74個,基因、rRNA、tRNA數(shù)量和GC含量均符合大腸桿菌基因組的基本特征(表5)。
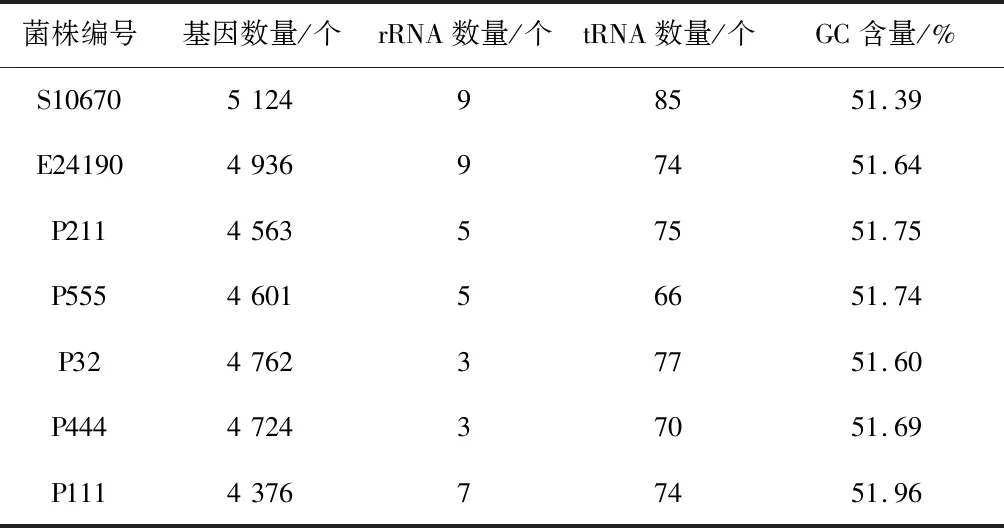
表5 基因預測結果統(tǒng)計
2.6 COG功能注釋
本研究7株菌株基因的COG分類得到22類,菌株S10670、E24190、P211、P555、P32、P444和P111注釋到的功能基因數(shù)量分別為3 482個、3 483個、3 274個、3 286個、3 342個、3 341個和3 183個。由圖5可知,注釋到的與代謝相關基因的數(shù)量最多。
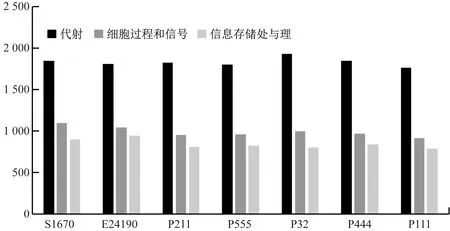
圖5 COG注釋分類統(tǒng)計
2.7 VFDB注釋
通過與毒力因子數(shù)據(jù)庫(VFDB)比對,菌株S10670、E24190、P211、P555、P32、P444和P111注釋到的毒力基因數(shù)量分別為126、61、52、55、47、44、38個,其中分泌系統(tǒng)和黏附與侵襲毒力基因最多。由表6可知,強毒株S10670的基因組中含有志賀毒素基因(Stx),溶血素基因(hlyA,hlyB,hlyC,hlyD,hlyE/clyA),黏附相關的毒力基因有外膜蛋白黏附素基因(paa)、緊密黏附素基因eae、緊密黏附素易位受體蛋白基因Tir、toxB,血紅素吸收相關基因(chuA,chuT,chuW,chuX,chuY),大量III型分泌系統(tǒng)相關基因如NLE系列基因(nleA,nleB1,nleB2-1,nleC,nleE-1,nleF,nleG-1,nleG2-3,nleG7,nleH1-1,nleH1-2)、Esp系列基因(espB,espF,espG,map,tir,espJ,espL1,espL2,espL4,espM1,espR1,espR3,espR4,espW,espX1~espX6,espY1~espY5)、Esc系列基因(escC,escD,escF,escJ,escL,escN,escO,escR,escS,escT,escU,escV)和VI型分泌系統(tǒng)相關基因(aec15~aec19,aec22~aec32);強毒株E24190基因組中含有腸毒素基因(LT)、溶血素基因(hlyA)和CFA/I菌毛相關基因(cfaA,cfaB,cfaC,cfaD/cfaE);中等毒株P211基因組中含有CFA/I菌毛相關基因(cfaA,cfaB,cfaC,cfaD/cfaE)、P菌毛相關基因(papA,papC,papD,papH)和鐵轉運系統(tǒng)吸收系統(tǒng)相關基因(sitA,sitB,sitC,sitD);中等毒株P555基因組中含有12個VI分泌系統(tǒng)相關基因(aec15~aec19,aec22~aec32);弱毒株P32、P444和111基因組中含有的毒力基因也存在于菌株S10670、E24190、P211和P555中,如黏附相關毒力因子大腸桿菌普通菌毛(ECP)、大腸桿菌層粘連蛋白結合蛋白菌毛(ELF)、出血性大腸桿菌菌毛(HCP)和I型菌毛(Fim),侵襲素相關毒力因子(ibeB,ibeC),鐵轉運系統(tǒng)相關毒力基因(entA,entB,entC,entE,entS,hitC)分泌系統(tǒng)相關毒力基因(espL1,espL4,espR1,espX1,espX4,espX5,aec15)和溶血素基因(hlyE/clyA)。
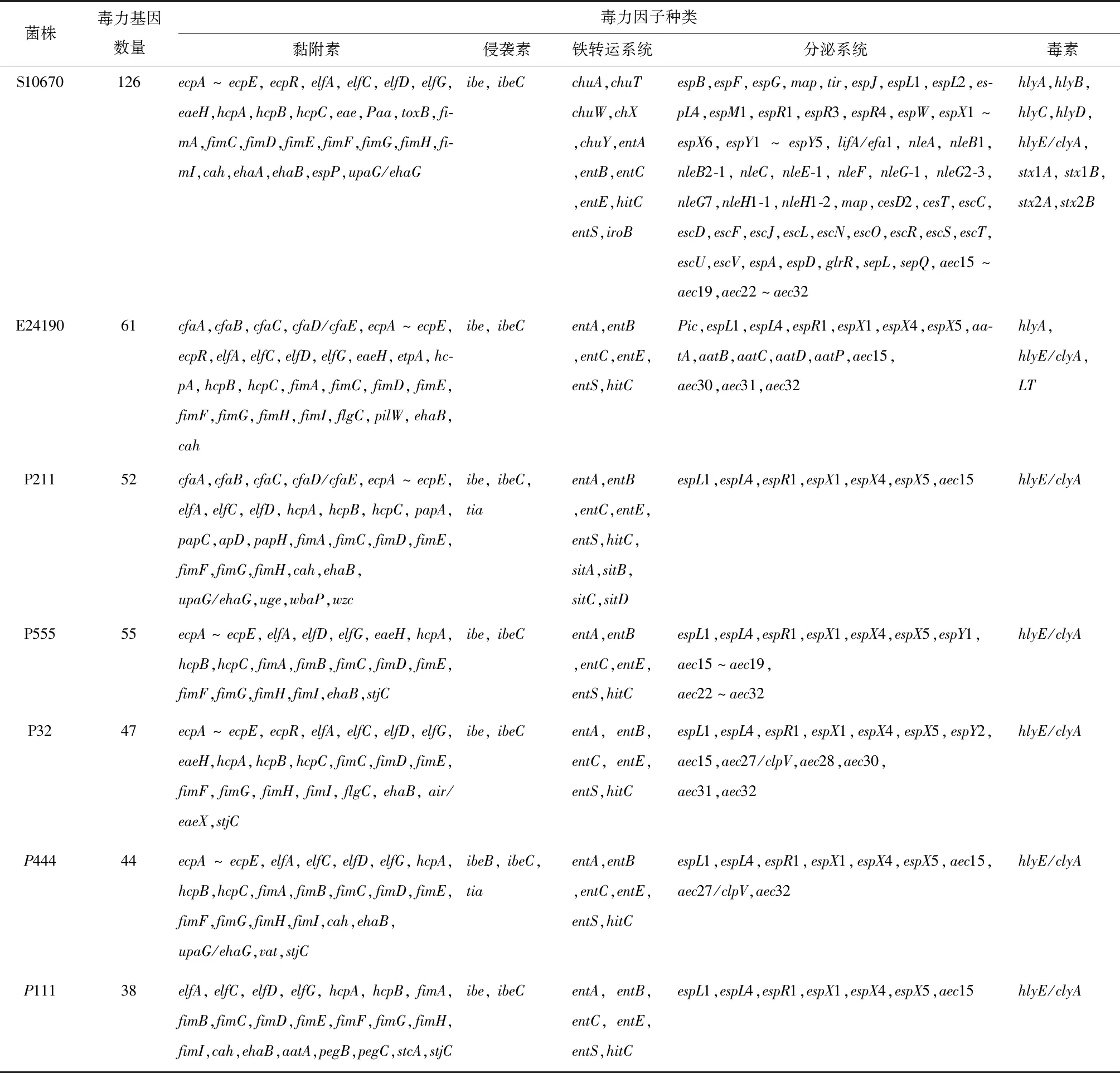
表6 毒力因子統(tǒng)計
3 討論與結論
大腸桿菌的致病性是由多種毒力因子互相協(xié)作產(chǎn)生。志賀樣毒素(Stx)是一種志賀毒素,具有細胞毒性、腸毒性、神經(jīng)毒性作用,是毒性最強的細菌毒素之一,包括Stx1和Stx2兩種亞型,Stx2較Stx1的毒力更強,Stx2的變異體Stx2e是導致仔豬發(fā)生水腫病的重要毒素[15]。緊密黏附素eae能使細菌緊密黏附到腸上皮細胞,eae與緊密黏附素易位受體蛋白Tir結合引起進一步的信號傳導,細胞內(nèi)鈣離子濃度驟增,腸內(nèi)液體分泌增加,引起特征性的粘附脫落病變(attaching and effacing,A/E)[16,17]。Chu系列基因編碼的蛋白與血紅素的吸收有關,血紅素是宿主體內(nèi)最豐富的鐵離子資源,主要由chuA基因從細胞外或者宿主的血紅素中攝取鐵和血紅蛋白中的鐵元素[18]。溶血素(Hemolysin)基因(hlyA,hlyB,hlyC,hlyD,hlyE/clyA)孔道形成蛋白家族的成員,溶血素可溶解哺乳動物紅細胞,在靶細胞膜上形成孔道繼而殺死靶細胞[19]。Ⅲ型分泌系統(tǒng)效應因子( Non-LEE encoded effactor ,Nle ) NleH和NleD具有抗凋亡活性,NleH能夠促進細胞的生存,抑制腸細胞的丟失,從而維持大腸桿菌的定值[20]。在感染期過程中,大腸桿菌表面分子能夠誘導外源性的細胞凋亡,Ⅲ型分泌系統(tǒng)效應因子如EspF,Map能夠誘導內(nèi)源性的凋亡途徑[21]。EspF能夠誘導線粒體的溶解﹑破壞細胞緊密聯(lián)接結構和促進抗凋亡蛋白的降解[22]。Map能夠破壞細胞之間的緊密聯(lián)結,誘導線粒體功能失活[23]。毒力因子Stx、eae、Tir、Chu、hlyB、hlyC、hlyD和III型分泌系統(tǒng)相關基因Nle、EspF 、Map在本研究中僅存在于強毒株S10670中,hlyA存在于強毒株S10670和E24190中。熱敏腸毒素(LT)為一種免疫蛋白,與霍亂腸毒素(CT)有著密切關系,組成一個毒素家族,LT是產(chǎn)腸毒素大腸桿菌的主要致病因子之一,可引起水樣腹瀉[24]。LT在本研究中僅存在于強毒株E24190中。
VI型分泌系統(tǒng)(T6SS)是一種新型的分泌系統(tǒng),普遍存在于革蘭氏陰性菌中,最早是在霍亂弧菌中發(fā)現(xiàn)的,T6SS參與細菌生物膜的形成,并介導細菌與宿主之間的粘附和毒性作用[25],本研究檢測到的VI分泌系統(tǒng)是ACE T6SS相關毒力因子。ACE T6SS僅存在于強毒株S10670和中等毒株P555中。CFA/I是一種優(yōu)勢血清型定居因子,整個CFA/I定居因子由4個基因 (cfaA,cfaB,cfaC,cfaD/E) 編碼[26,27],編碼的相應多肽稱為cfaA,cfaB,cfaC,cfaD/E,成熟定居因子由cfaB和cfaE組成,cfaB是主要結構亞單位,cfaE位于定居因子頂端,二者均能介導CFA/I對人小腸上皮細胞相應受體的結合,致病性大腸桿菌通過其菌體表面的宿主特異性定居因子CFA/I介導粘附于小腸黏膜上皮細胞[28],大量增殖,釋放腸毒素,引起仔豬腹瀉,CFA/I在本研究中存在于強毒株E24190和中等毒株P211中。
P型菌毛多分布于尿道致病性大腸桿菌(UPEC)的黏附素,能介導大腸桿菌粘附于上尿道,P型菌毛是UPEC導致腎盂腎炎的主要毒力因子,已知的編碼P菌毛操縱子的基因有papA、papB、papC、papD、papE、papF、papG、papH和papI,其中 PapA 蛋白是 P 菌毛的主要成分[29]。鐵/錳轉運蛋白(Iron/manganese transport)是鐵攝取轉運系統(tǒng)毒力因子,由sitA、sitB、sitC和sitD四個基因編碼組成,該系統(tǒng)中的sitB是ATP結合蛋白,所以該系統(tǒng)相當于是ATP離子泵,可以提供細菌轉運過程所需的能量,外膜轉運系統(tǒng)還可以將二價鐵載入細胞內(nèi),參與營養(yǎng)物質(zhì)的攝取、細菌毒素的分泌以及將抗生素泵出細胞使細菌具有抗藥性[30]。P菌毛和鐵/錳轉運蛋白本研究中僅存在于中等毒株P211中。
綜上所述,本研究通過細菌感染小鼠鑒別7株菌株的致病力,采用二代測序技術測定7株致病性大腸桿菌的基因組序列,經(jīng)組裝,得到致病力相關的毒力基因,研究結果為大腸桿菌分子致病機制奠定理論基礎。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
