
昆蟲取食和機械損傷對棉花和玉米脂氧合酶活性的誘導作用
2021-07-01 09:27:05呂敏衛甜劉懷阿蘇建坤
江蘇農業科學 2021年10期
呂敏 衛甜 劉懷阿 蘇建坤
摘要:脂氧合酶(LOX)是植物防御反應中的關鍵酶,脂氧合酶途徑的產物包括茉莉酸、綠葉揮發物等,在植物防御反應中起著重要作用。為了明確脂氧合酶在昆蟲取食和機械損傷誘導植物通信中的作用,以棉花、玉米為研究對象,分析不同處理(包括棉蚜取食和機械損傷誘導棉花及其相應揮發物誘導鄰近玉米、玉米螟取食和機械損傷誘導玉米及其相應揮發物誘導鄰近棉花)的誘導防御機制,測定不同方法處理24、48、72 h后棉花、玉米的脂氧合酶的活性。結果表明,昆蟲取食和機械損傷不僅會誘導棉花、玉米植株的LOX活性升高,還能誘導鄰近健康植株的LOX活性升高。研究結果表明,脂氧合酶在昆蟲取食和機械損傷誘導的棉花、玉米間通信中起作用。
關鍵詞:脂氧合酶;昆蟲取食;機械損傷;棉花;玉米;誘導防御機制
中圖分類號: S188+.3 ?文獻標志碼: A ?文章編號:1002-1302(2021)10-0086-05
植物在其生長發育過程中形成了多種誘導防御機制來抵御生物因子的入侵[1],昆蟲取食作為一種重要的生物脅迫而廣泛存在于自然界中[2]。面對昆蟲的取食危害,植物形成了直接誘導和間接誘導2種防御機制。直接誘導防御是指植物產生次生代謝物質直接殺傷昆蟲,間接誘導防御是指植物通過釋放一系列特定的揮發性化合物吸引昆蟲的捕食性、寄生性天敵,從而減少昆蟲的危害。脂氧合酶是昆蟲取食誘導的揮發性化合物形成過程中的關鍵酶。
脂氧合酶(lipoxygenase,簡稱LOX)在生物體內主要催化含有(Z,Z)-1,4戊二稀結構單元的不飽和脂肪酸的加氧反應,生成過氧化物及其一系列次生產物。脂氧合酶是植物代謝過程中的關鍵酶,它在調節植物的生長發育、成熟衰老、抗蟲、抗病及抗脅迫等過程中起著重要作用[3]。脂氧合酶通常處于靜止狀態,只有當生物體發育到一定階段或受到環境脅迫時才啟動[4-5]。脂氧合酶的誘導表達作用已經在植物受到機械損傷、昆蟲取食和病原菌侵染中得到證明[6-10]。楊毛臀螢葉甲取食能誘導榿木葉片中酚類物質的含量增加,使得在防御反應中起作用的多酚氧化酶、脂氧合酶和過氧化物酶的活性增加,蛋白酶抑制劑也增多[11-13]。大豆通過LOX途徑對昆蟲的取食作出反應,大豆夜蛾(Anticarsia gemmatalis)取食大豆植物后會誘導脂氧合酶活性增加、蛋白酶抑制劑增多,同時,茉莉酸會激活編碼蛋白酶抑制劑基因的轉錄[14]。
昆蟲取食還能誘導植物間的通信作用,即昆蟲取食不僅能誘導植物本身產生防御反應,還能誘導鄰近健康植株產生防御反應[15-16]。植物間是通過空氣進行交流的,暴露于二斑葉螨取食誘導利馬豆釋放的揮發物中的健康利馬豆和黃瓜對捕食性螨的吸引力都增強[17-19]。Karban等通過田間試驗發現,受傷艾草灌叢附近的煙草多酚氧化酶活性升高,受蝗蟲、煙草天蛾取食程度的影響較小[20]。
棉花、玉米是重要的經濟作物和糧食作物,研究它們之間的通信作用可為棉花、玉米的間作套種提供依據。本研究分析受到機械損傷和棉蚜取食棉花誘導棉花及其鄰近健康玉米,以及受到機械損傷和玉米螟取食玉米誘導玉米及其鄰近棉花的脂氧合酶活性,以期明確昆蟲取食和機械損傷誘導棉花和玉米間通信作用的機制,有利于找到害蟲綜合防治的新途徑。
1 材料與方法
1.1 試驗材料
棉花品種為中棉35,將種子催芽后種于沙土(已滅菌)中,等到子葉完全展開后,將幼苗移栽到盛有營養液[21]的塑料杯(容量為250 mL)中,在溫室中培養1~2周后供試。玉米品種是農大108,將玉米種子浸泡12 h后種于營養土中2周,再用于試驗。
1.2 供試昆蟲
供試昆蟲為棉蚜,為室內飼養的品系,飼養在日光溫室內(溫度為25~35 ℃,相對濕度為60%~80%,自然光周期);玉米螟卵購自中國農業科學院植物保護研究所,室內飼養至3齡。飼養溫度為 25~28 ℃,相對濕度為60%~80%,光—暗周期為 16 h—8 h。
1.3 試驗處理
試驗參照呂敏等的處理方法[15]進行處理。
1.4 酯氧合酶活性的測定方法
將棉花、玉米幼苗剪去根部后洗凈,在液氮中研磨,加入0.1 g/g聚乙烯吡咯烷酮(PVPP)和 3 mL/g 濃度為0.1 mol/L、pH值為7.0、含Triton 0.1%的磷酸緩沖液(棉苗)或0.1 mol/L pH值為6.5的磷酸緩沖液(玉米),離心(4 ℃、12 000 g)20 min,上清液用于酶活性的測定。棉苗、玉米的 3 mL 反應體系為2.875 mL反應緩沖液、25 μL底物、100 μL酶液。棉苗的反應緩沖液為pH值為3.5的甘氨酸-鹽酸緩沖液。
測定中用到的緩沖液:pH值為3.0~3.5的甘氨酸-鹽酸緩沖液,pH值為4.0~5.5的檸檬酸緩沖液,pH值為6.0~7.5的磷酸緩沖液,pH值為 8.0~9.0的硼酸緩沖液。底物的配制和活性測定參考Axelord等的方法[22]。
1.5 蛋白質含量的測定
參照Braford的考馬斯亮蘭G-250法測定蛋白質含量[23],以牛血清白蛋白作為標準蛋白制作標準曲線。
1.6 數據處理
各試驗數據用Excel進行作圖分析,并用SPSS軟件進行處理間的差異顯著性檢驗。
2 結果與分析
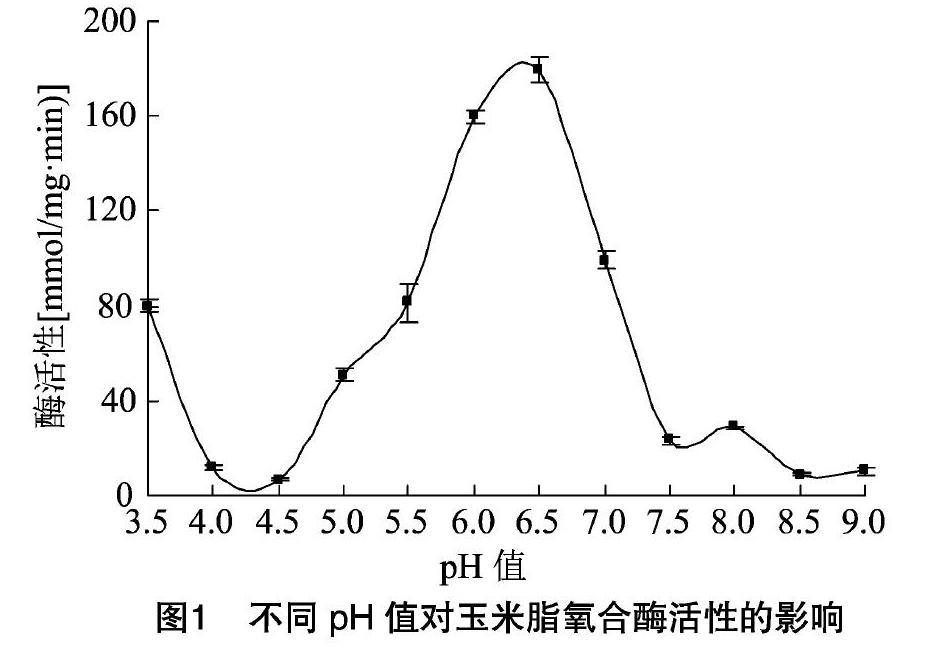
2.1 脂氧合酶的最適pH值
棉花脂氧合酶的最適pH值為3.5,本試驗測定了pH值為3.5~9.0的玉米脂氧合酶活性的變化。如圖1所示,玉米脂氧合酶的活性在pH值為6.5時最高。
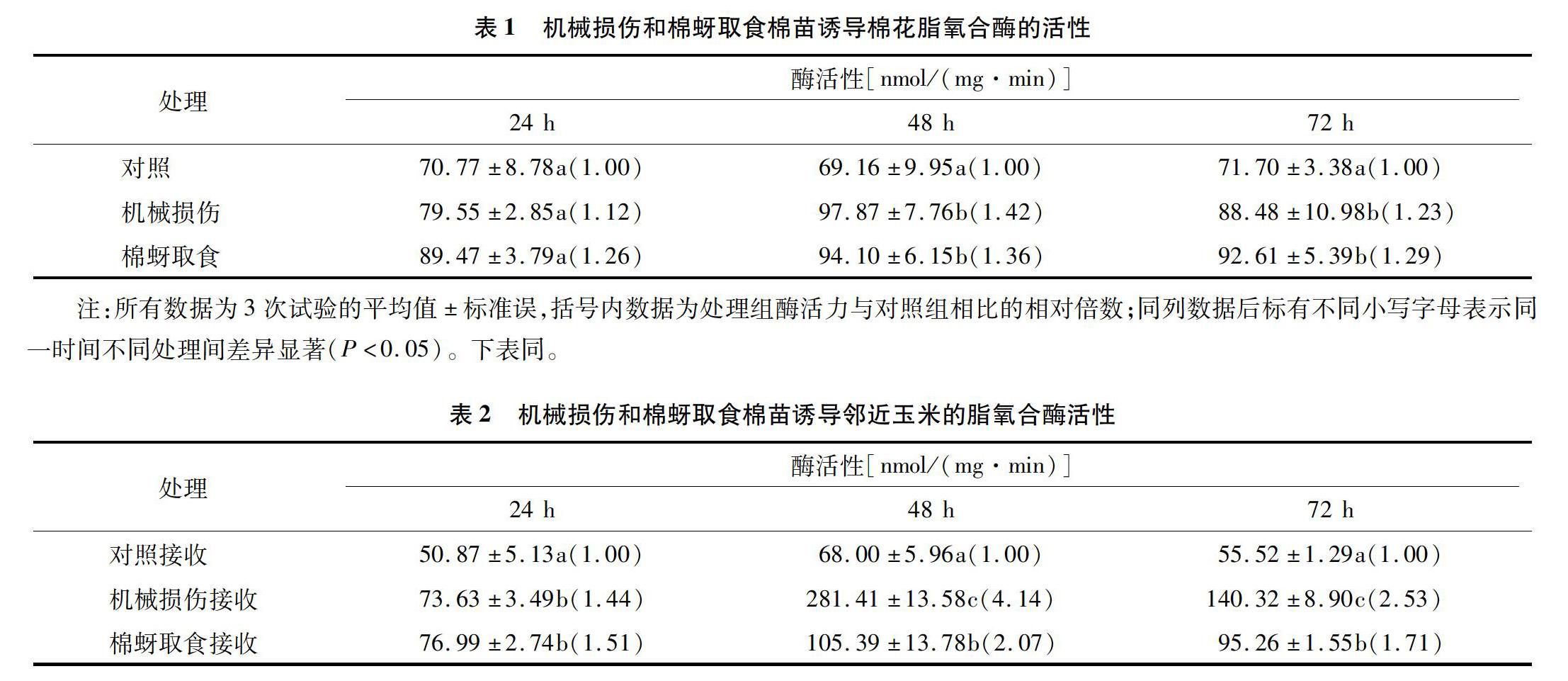
2.2 機械損傷和棉蚜取食誘導棉花及鄰近玉米脂氧合酶活性的變化
從表1、表2可以看出,機械損傷和棉蚜取食誘導棉花及其相應誘導揮發物接收玉米的脂氧合酶的活性與對照相比均有所增加。在處理后24、48、72 h,機械損傷誘導棉花脂氧合酶的活性分別為對照的1.12、1.42、1.23倍,棉蚜取食誘導棉花脂氧合酶的活性分別是對照的1.26、1.36、1.29倍,在處理后48 h達到最大值,為94.10 nmol/(mg·min)。在處理后24 h,機械損傷、棉蚜取食誘導揮發物接收玉米的脂氧合酶活性分別是對照的1.44、1.51倍,在處理后48 h,酶活性達到最大值,分別為281.41、105.39 nmol/(mg·min),分別是對照的4.14、2.07倍,隨后在處理后72 h下降,但是處理72 h后的LOX活性仍然與對照間差異顯著(P<0.05),分別是對照的2.53、1.71倍。
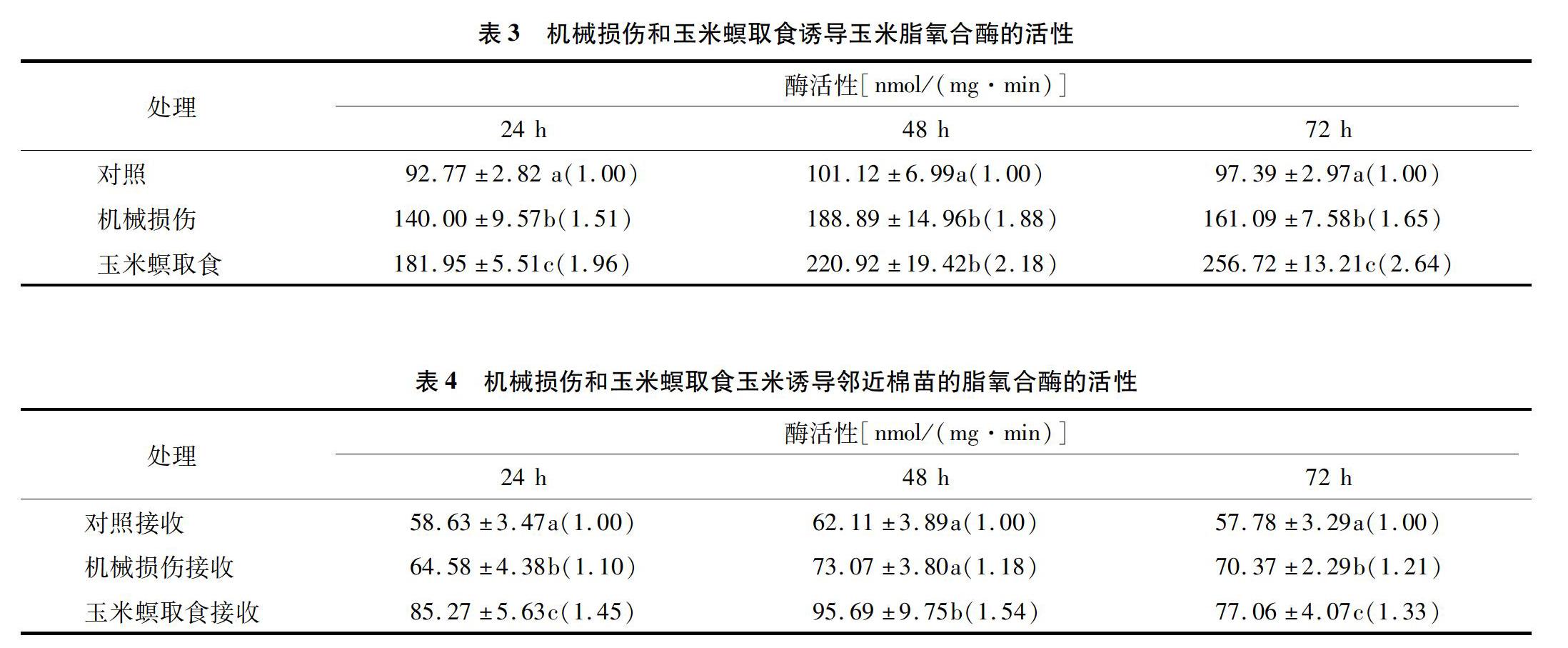
2.3 機械損傷和玉米螟取食誘導玉米鄰近棉苗脂氧合酶活性的變化
從表3、表4可以看出,玉米螟取食和機械損傷誘導玉米及其相應誘導揮發物接收棉苗的脂氧合酶的活性與對照相比均有所增加。機械損傷誘導玉米的脂氧合酶活性隨時間增加,到48 h達到最大值,隨后下降,而玉米螟取食誘導玉米脂氧合酶的活性最大值在72 h,為對照組的2.64倍。在24、48、72 h,機械損傷誘導揮發物接收棉花的脂氧合酶活性分別是對照組的1.10、1.18、1.21倍;玉米螟取食誘導揮發物接收棉苗脂氧合酶的活性分別為對照組的1.45、1.54、1.33倍。
3 討論與結論
脂氧合酶是脂氧合酶途徑中的關鍵酶。脂氧合酶途徑可以生成一系列綠葉揮發性化合物,受傷植物釋放的綠葉揮發物可直接殺傷植食性昆蟲[24-25]。此外,脂氧合酶途徑還參與信號分子的合成,如合成產物茉莉酸,在植物防御反應信號交流中發揮了重要的作用。昆蟲取食和機械損傷能誘導植物脂氧合酶活性增加。豆青葉甲和三刺苜蓿角蟬取食誘導了番茄、大豆和棉花的脂氧合酶活性增加[26]。Qin等研究發現,機械損傷和棉蚜取食能誘導棉苗體內苯丙氨酸解氨酶和脂氧合酶活性升高[27]。本研究發現,棉花被棉蚜取食或受到機械傷害后,自身的脂氧合酶活性增加,這與秦秋菊的研究結果[28]一致,說明脂氧合酶途徑參與了由機械損傷和昆蟲取食誘導的棉花防御反應。試驗結果還表明,機械損傷和玉米螟取食誘導玉米的脂氧合酶的活性顯著增加(P<0.05)。玉米螟取食誘導玉米的脂氧合酶的活性比機械損傷誘導的高,這是由于昆蟲取食和機械損傷對植物的誘導作用不同。因為許多昆蟲取食后還能在傷口部位引入特異性激發子,如甜菜夜蛾(Spodoptera exigua)口腔分泌物中的脂肪酸-氨基酸軛合物(FACs)[29]。另外,機械損傷與昆蟲取食損傷的不同還表現在時間上,玉米螟取食誘導玉米脂氧合酶的活性在72 h達到最大值,而機械損傷誘導玉米脂氧合酶的活性在48 h達到最大值。
越來越多的研究證明了植物間通信的存在。包括同種植物間的通信[17-18,27,30],異種植物間的通信作用也被很多試驗證明[19-20]。秦秋菊等研究發現,棉蚜取食和機械損傷還能引起鄰近的健康棉花脂氧合酶活性增加[28]。Lv等的研究結果證明了昆蟲取食和機械損傷能誘導棉花和玉米間通信作用的存在,且苯丙氨酸解氨酶在其中起作用[31]。本試驗結果表明,暴露于棉蚜取食和機械損傷的揮發物中健康玉米的脂氧合酶活性增加。另外,暴露于玉米螟取食和機械損傷揮發物中的棉苗的脂氧合酶活性也增加,表明脂氧合酶在昆蟲取食和機械損傷誘導的棉花和玉米間通信作用中起作用。
暴露于植物衍生的揮發性化合物能激發鄰近植物的防御反應[28,32-36],昆蟲取食誘導產生的植物揮發性化合物通過表皮吸收或激活生理反應影響鄰近植物[37]。昆蟲取食誘導的植物揮發性化合物因不同的取食方式有不同的途徑。蒺藜苜蓿被毛蟲和螨取食會產生不同種不同量的揮發性化合物,茉莉酸、水楊酸都有積累,但是這種積累因口器不同而不同。螨類取食誘導的水楊酸積累量高于毛蟲取食誘導的,同時這2種昆蟲取食的茉莉酸積累的時間不同[38]。Turlings等對比分析了甜菜夜蛾取食后釋放的揮發性化合物并與健康植株釋放的揮發性化合物發現,在玉米被害后0~1 h內,二者沒有區別,都是綠葉揮發物,5~6 h后,與對照的健康葉片相比,被害植株釋放大量萜烯等(herbivore-induced plant volatiles,簡稱HIVs)[39],這更加證明不同昆蟲取食誘導植物釋放的揮發性化合物量和時間上不同。本試驗得出,玉米螟取食誘導揮發物接收棉花以及棉蚜取食誘導揮發物接收玉米的LOX的活性都在48 h達到最大。這可能與揮發物釋放的時效性有關,須要進一步分析棉蚜取食誘導棉花及玉米螟取食誘導玉米產生的揮發性化合物的成分及其釋放時效。
參考文獻:
[1]Karban R,Baldwin I T. Induced response to herbivory[M]. Chicago:Chicago University Press,1997.
[2]張慶花,陳迎迎,張海龍,等. 1-戊烯-3-酮在植物防御反應中的作用[J]. 植物生理學報,2019,55 (3):225-231.
[3]吳桂玲,馮定坤. 植物脂氧合酶的研究進展[J]. 廣州化工,2019,47(17):37-39.
[4]Gardner W H. Biological roles and biochemistry of the lipoxygenase pathway[J]. HortScience,1995,30 (2):197-205.
[5]李翠霞,李志忠,張 繼. 外源誘導物對百里香植株再生過程中脂氧合酶活性的影響[J]. 草業科學,2012,29(9):1390-1395.
[6]Saravitz D M,Siedow J N. The differential expression of wound-inducible lipoxygenase genes in soybean leaves[J]. Plant Physiology,1996,110(1):287-299.
[7]Wang C,Zien C A,Afitlhile M,et al. Involvement of phospholipase D inwound-induced accumulation of jasmonic acid in Arabidopsis[J]. The Plant Cell,2000,12(11):2237-2246.
[8]Vieira A A,Oliveira M G A,José I C,et al. Biochemical evaluation of lipoxygenase pathway of soybean plant submitted to wounding[J]. Brazilian Journal of Plant Physiology,2001,13(1):5-12.
[9]da Silva Fortunato F,de Almeida Oliveira M G,Brumano M H N,et al. Lipoxygenase-induced defense of soybean varieties to the attack of the velvetbean caterpillar (Anticarsia gemmatalis Hübner)[J]. Journal of Pest Science,2007,80:241-247.
[10]Melan M A,Dong X,Endara M E,et al. An Arabidopsis thaliana lipoxygenase gene can be induced by pathogens,abscisic acid,and methyl jasmonate[J]. Plant Physiology,1993,101(2):441-450.
[11]Pea-Cortes H,Sanchez-Serrano J,Rocha-Sosa M,et al. Systemic induction of proteinase-inhibitor-II gene expression in potato plants by wounding[J]. Planta,1988,174(1):84-89.
[12]Farmer E E,Ryan C A. Octadecanoid precursors of jasmonic acid activate the synthesis of wound-inducible proteinase inhibitors[J]. Plant Cell,1992,4(2):129-134.
[13]Koiwa H,Bressan R A,Hasegawa P M. Regulation of protease inhibitors and plant defense[J]. Trends in Plant Science,1997,2(10):379-384.
[14]Fortunato F S,Oliveira M G A,Brumano M H N,et al. Effect of the Anticarsia gemmatalis injury on the lipoxygenases activity from soybean leaves[J]. Bioscience Journal,2004,20(2):37-46.
[15]呂 敏,孫婳婳,高希武. 昆蟲取食誘導棉花和玉米多酚氧化酶和過氧化物酶的活性研究[J]. 中國農學通報,2012,28(6):211-216.
[16]Rhoades D F. Responses of alder and willow to attack by tent caterpillars and webworms:evidence for pheromonal sensitivity of willows[M]//Hedin P A. Plant resistance to insects. Washington DC:American Chemical Society,1983:55-68.
[17]Bruin J,Dicke M,Sabelis M W. Plants are better protected against spider-mites after exposure to volatiles from infested conspecifics[J]. Experientia,1992,48:525-529.
[18]Dicke M. Evolution of induced indirect defence of plants[M]//Tollrian R,Harvell C D. The ecology and evolution of inducible defenses. Princeton,New Jersey,USA:Princeton University Press,1990:62-88.
[19]Oudejans A M C,Bruin J. Dose spider-mite damage induce information transfer between plants of different species ?[J]. Med Fac Landbouww Univ Gent,1995,59:733-739.
[20]Karban R,Baldwin I T,Baxter K J,et al. Communication between plants:induced resistance in wild tobacco plants following clipping of neighboring sagebrush[J]. Oecologia,2000,25:66-71.
[21]Liang R X,Li C J.Diferences in cluster-root formation and carboxylate exudation in Lupinusa albus L. and different nutrient deficiencies[J]. Plant Soil,2003,248:221-227.
[22]Axelrod B,Cheesbrough T M,Laakso S. Lipoxygenase from soybeans[J]. Methods in Enzymology,1981,71:441-451.
[23]Bradford M M A. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding[J]. Analytical Biochemistry,1976,25(1):248-256.
[24]Farmer E E. Surface-to-air signal[J]. Nature,2001,411(6839):854-856.
[25]Turlings T C J,Benrey B. Effects of plant metabolites on the behavior and development of parasitic wasps[J]. Ecoscience,1998,5:321-333.
[26]Felton G W,Summers C B,Mueller A J. Oxidative response in soybean foliage to herbivory by bean leaf beetle and three cornered alfalfa hopper[J]. Journal of Chemical Ecology,1994,20:639-650.
[27]Qin Q J,Shi X Y,Liang P,et al. Induction of phenylalanine ammonia-lyase and lipoxygenase by artificial damage and aphid infestation in cotton seedlings[J]. Progress in Natrual Science,2005,15(5):419-423.
[28]秦秋菊. 害蟲取食和機械損傷誘導的棉花防御反應[D]. 北京:中國農業大學,2005.
[29]Turlings T C J,Alborn H T,Loughrin J H,et al. Volicitin,an elicitor of maize volatiles in oral secretion of Spodoptera exigua:isolation and bioactivity[J]. Journal of Chemical Ecology,2000,26:189-202.
[30]Tscharntke T,Thiessen S,Dolch R,et al. Herbivory,induced resistance,and interplant signal transfer in Alnus glutinosa[J]. Biochemical Systematics and Ecology,2001,29:1025-1047.
[31]Lv M,Kong H L,Liu H A,et al. Induction of phenylalanine ammonia-lyase (PAL) in insect damaged and neighboring undamaged cotton and maize seedlings[J]. International Journal of Pest Management,2017,63(2):166-171.
[32]Arimura G,Ozawa R,Shimoda T,et al. Herbivory-induced volatiles elicit defence genes in lima bean leaves[J]. Nature,2000,406(6795):512-515.
[33]Arimura G,Ozawa R,Horiuchi J,et al. Plant-plant interactions mediated by volatiles emitted from plants infested by spider mites[J]. Biochemical Systematics and Ecology,2001,29(10):1049-1061.
[34]Arimura G,Tashiro K,Kuhara S,et al. Gene responses in bean leaves induced by herbivory and by herbivore-induced volatiles[J]. Biochemical and Biophysical Research Communications,2000,277(2):305-310.
[35]Alméras E,Stolz S,Vollenweider S,et al. Reactive electrophile species activate defense gene expression in Arabidopsis[J]. Plant Journal,2003,34(2):202-216.
[36]Weber H,Chételat A,Reymond P,et al. Selective and powerful stress gene expression in Arabidopsis in response to malondialdehyde[J]. Plant Journal,2004,37(6):877-888.
[37]Choh Y,Shimoda T,Ozawa R,et al. Exposure of lima bean leaves to volatiles from herbivore-induced conspecific plants results in emission of carnivore attractants:active or passive process?[J]. Journal of Chemical Ecology,2004,30(7):1305-1317.
[38]Leitner M,Boland W,Mithofer A. Direct and indirect defences induced by piercing-sucking and chewing herbivores in Medicago truncatula[J]. New Phytologist,2005,167(2):597-606.
[39]Turlings T C J,Tumlinson J H,Heath R R,et al. Isolation and identification of allelochemicals that attract the larval parasitoid Cotesi marginiventris (Gresson) to the microhabitat of one of its hosts[J]. Journal of Chemical Ecology,1991,17(11):2235-2251.
