不同分子質量膠原抗凍肽的抗凍性能及機理研究
2021-07-02 13:30:04劉含曹慧徐斐袁敏葉泰于勁松黎燕陰鳳琴吳秀秀郝麗玲
食品與發酵工業 2021年11期
劉含,曹慧,徐斐,袁敏,葉泰,于勁松,黎燕,陰鳳琴,吳秀秀,郝麗玲
(上海理工大學 醫療器械與食品學院,上海,200093)
抗凍蛋白(antifreeze proteins,AFPs) 是生物為適應極端寒冷環境而產生的一類特異性糖肽或多肽[1],它能夠以非依數的形式降低溶液的冰點而不改變其熔點,從而導致溶液的熔點和冰點之間出現差值,這種差值稱為熱滯活性[1-2]。AFPs還可通過氫鍵吸附于冰晶表面,從而控制冰晶生長以及抑制冰晶之間發生重結晶。AFPs的熱滯活性及重結晶抑制活性使得其能減少或防止在凍融循環過程中冰晶對細胞和組織的傷害,并在結冰或亞結冰條件下保護生物體不受傷害[3],因而在食品、血液以及器官和組織的冷凍儲存中有廣泛的應用前景[4]。
迄今為止,人們在植物、海洋魚類、昆蟲及真菌中都發現了不同類型的AFPs[5-6],但成本高、產率低等不足仍是限制AFPs產業化制備的主要因素,因而尋找合適的AFPs原料并提取出高熱滯活性的AFPs是亟需解決的問題。動物皮是膠原蛋白的主要載體,從膠原蛋白的特異性氨基酸序列及空間結構分析,膠原蛋白可能含有高活性的膠原抗凍肽[7]。我國具有豐富的牦牛資源,牦牛皮中膠原蛋白的含量達到其干重的70%以上。但目前牦牛皮主要應用于皮革的制作,利用率較低。因而利用牦牛皮作為提取AFPs的原料不僅可以有效利用畜禽加工副產物實現價值增值的目的,還能滿足市場對高性價比AFPs的需求。
膠原抗凍肽的分子質量對其抗凍特性有著顯著影響。BAR DOLEV等[8]研究發現南極細菌Marinomonasprimoryensis能分泌具有附著性的AFP1,使細菌附著在冰頂部以獲得氧氣和營養物質。該AFP是1種由 5 個不同區域組成的長蛋白質,相對分子質量 1.5 MDa,其中第4個相對分子質量為 34 kDa的特定區域能與冰晶發生特異性的結合。DAMODARAN等[9]利用蛋白酶水解魚皮明膠,證實了分子質量在1 000~2 500 Da的明膠多肽具有抑制冰晶生長的作用。這些實驗表明了不同分子質量的抗凍肽抑制冰晶生長的能力并不相同,但目前對不同分子質量的膠原抗凍肽抑制冰晶生長作用機理的研究卻很少。因此本實驗采用熱水抽提耦合酶水解法,制備了膠原蛋白酶解產物,并應用超濾膜技術得到了<1 k、1~3 k、3~5 k及>5 kDa這4種不同分子質量的膠原抗凍肽。在此基礎上,對不同分子質量范圍膠原抗凍肽的熱滯活性、重結晶抑制能力及修飾冰晶生長的特性進行了研究,同時總結了膠原抗凍肽與冰晶相互作用的機理。
1 材料與方法
1.1 實驗材料
新鮮牦牛皮,青海省屠宰場。將牦牛皮去毛、洗凈、絞碎,并采用質量分數為5%的 Na2CO3溶液脫脂18 h,進一步采用質量分數為5%的NaCl溶液去除鹽溶性非膠原成分,蒸餾水漂洗后貯藏于-20 ℃備用;牛血清白蛋白(bovine serum albumin,BSA),上海伯奧生物科技有限公司;木瓜蛋白酶(200 NFU/mg),南寧龐博公司;其余試劑均為分析純,國藥集團化學試劑有限公司。
1.2 實驗儀器
Pyris Diamond差示掃描量熱儀,美國Perkin Elmer 公司;RO-UF-4010(MSM-1812)型膜分離裝置超濾納濾反滲透,1 kDa、3 kDa、5 kDa等0.2 m2卷式膜組件,上海摩速科學器材有限公司;BP211D型天平,德國賽多利斯公司 (精確到0.01 mg) ; J-20XP型冷凍離心機,上海安亭科學儀器廠;BX51顯微鏡,日本奧林巴斯公司;FDCS冷熱臺,英國LinKam公司;Waters2414示差折光檢測器,美國沃特斯公司;Kje ltec 8400型全自動凱氏定氮儀, 丹麥福斯集團公司;LD85B3型真空冷凍干燥機,美國MillRock科技公司;TLID-2005袖珍型低溫恒溫槽,金壇市華峰儀器有限公司;Agilent1100型氨基酸自動分析儀,美國Agilent公司;79-1恒溫磁力攪拌器,寧波天恒儀器廠。
1.3 實驗方法
1.3.1 牦牛皮常規成分的測定
牦牛皮中的水分含量采用常壓干燥法測定(GB 5009.3—2016);粗蛋白含量采用微量凱氏定氮法測定(GB 5009.5—2016);粗脂肪含量采用索氏抽提法測定(GB 5009.6—2016)。
1.3.2 牦牛皮中膠原蛋白的提取
稱取20 g牦牛皮置于燒杯中,以1∶30的料液比(g∶mL)添加蒸餾水,在微沸狀態下加熱5 h,待溶液冷卻至室溫后置于酶反應器內。參考WANG等[10]的方法,并通過預實驗得到膠原蛋白的最佳酶解條件為:木瓜蛋白酶添加量2 000 U/g,溫度55 ℃,酶解時間48 h。酶解完成后5 000 r/min離心30 min,取上清液測定羥脯氨酸(Hyp)含量。
1.3.3 膠原蛋白含量測定
膠原蛋白含量采用WOESSNER[11]的方法進行測定。稱取4 g牦牛皮,加入30 mL 3 mol/L HCl 溶液于105 ℃下水解 16 h,水解完畢后過濾,獲得上清液。取4 mL稀釋好的上清液于比色管中,加入2 mL 14.1 g/mL氯胺T溶液,充分混勻后置于室溫下放置20 min以氧化Hyp;再加入2 mL顯色劑(10 g對二氨基苯甲醛,35 mL高氯酸,65 mL異丙醇,臨用前配制),并于60 ℃水浴加熱20 min;反應完畢后,在波長 560 nm 處測定溶液的吸光值,并建立Hyp標準曲線;根據標準曲線的公式和樣品吸光值計算膠原蛋白中Hyp 的含量,如公式(1)所示,膠原蛋白含量按公式(2)計算:
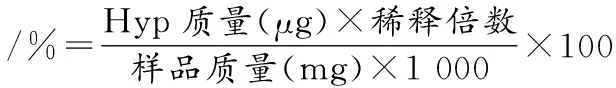
(1)
膠原蛋白含量/%=Hyp含量×7.46
(2)
式中:7.46為換算系數。
1.3.4 氨基酸組成
分別稱取10 mg不同分子質量的膠原抗凍肽于水解管中,加入2 mL 6 mol/L HCl溶液,再加入3~4滴苯酚;將水解管放入冰鹽水中預冷3~5 min,抽真空并封口,然后在110 ℃下水解 22 h;將水解后的樣品液轉移后過濾后轉移至量瓶內,用水定容至刻度,振蕩混勻。取1 mL濾液用氮吹儀60 ℃吹干,殘留物用檸檬酸鈉緩沖液復溶后取樣上機測定。色譜條件:C18柱(4.0 mm×125 mm);柱溫40 ℃;樣品流速1.0 mL/min;檢測波長440 nm(Hyp、Pro),570 nm(除Hyp、Pro以外的氨基酸);流動相A為20 mmol醋酸鈉溶液,B為V(20 mmol醋酸鈉溶液)∶V(甲醇)∶V(乙腈)=1∶2∶2。
1.3.5 膠原酶解產物分子質量分布的測定
采用水相凝膠滲透色譜法對酶解液中的多肽進行相對分子質量分布的測定。色譜條件:Waters2414示差折光檢測器;Waters凝膠色譜柱;流動相為0.1 mol/L的NaNO3溶液;流速0.5 mL/min;柱溫40 ℃。酶解液凍干粉配成5 mg/mL的溶液,進樣體積50 μL。
1.3.6 膠原蛋白酶解液的膜分離處理及得率計算
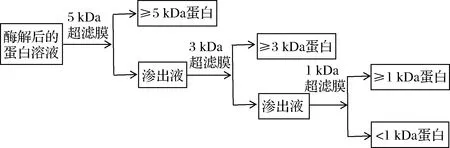
圖1 不同分子質量膠原抗凍肽制備工藝流程圖Fig.1 Preparation process of collagen antifreeze peptides with different molecular weights
膜分離設備的參數:壓力0.2 MPa,溫度25 ℃。根據1.3.3中的方法測定不同組分超濾液中膠原蛋白的含量,并根據公式(3)計算其得率:
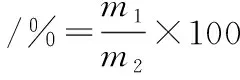
(3)
式中:m1,膜分離后一定分子質量范圍內的膠原抗凍肽總質量,g;m2,牦牛皮的總質量,g。
1.3.7 不同分子質量膠原抗凍肽的熱滯活性測定
稱取約10 mg樣品密封于鋁皿內,并放置于差示掃描量熱議內。當儀器充滿液氮并穩定后,以10 ℃/min的速率將樣品降溫至-20 ℃并保持5 min;然后以5 ℃/min的速率升溫至樣品呈固液共存狀態,即到達其保留溫度(holding temperature,Th),保持5 min;再將溫度以1 ℃/min的速率從Th降至-20 ℃。重復上述升降溫程序,分別記錄樣品的起始結晶溫度(T0)以及結晶熱(ΔHr)[9],并分別按照公式(4)計算熱滯活性值。以無抗凍活性的BSA作為對照。
熱滯活性=Th-T0
(4)
1.3.8 不同分子質量膠原抗凍肽的重結晶抑制效應
以質量分數40%的蔗糖溶液為溶劑,配制質量濃度100 mg/mL的抗凍肽溶液,利用低溫顯微鏡觀察其重結晶抑制能力。首先以10 ℃/min的速率將樣品溫度從25 ℃降至1 ℃,保持1 min以防止表面起霧;然后以90 ℃/min的速率降溫至-50 ℃以生成微小的冰晶,保持1 min;再以10 ℃/min的速率升溫至-6 ℃,保持40 min[12];圖像收集和分析由軟件IMAGE-PRO PLUS完成,圖像放大倍數為500倍。
1.3.9 不同分子質量膠原抗凍肽對冰晶生長形態的修飾
以40%的蔗糖溶液為溶劑,將制得的抗凍肽配制成100 mg/mL的溶液,并利用低溫顯微鏡觀察其對冰晶生長形態的影響。首先以10 ℃/min的速率將溫度從25 ℃降至-30 ℃,生成微小的冰晶,保溫1 min;然后以4 ℃/min的速率升溫,直至出現單冰晶,再以0.1 ℃/min的速率降溫,使冰晶生長[12]。圖像放大倍數為500倍,圖像收集和分析由軟件IMAGE-PRO PLUS完成,通過計算冰晶面積,評估添加不同分子質量抗凍肽的冰晶面積與降溫時間的關系。
2)在設計注水系統單站停運運行方案時,不僅要從下游注水井的壓力和水量需求分析,同時還要考慮上游污水系統調運的水量和水質是否能夠滿足注入系統要求。
2 結果與討論
2.1 牦牛皮的主要成分
牦牛皮的主要成分含量如表1所示。牦牛皮中水分含量最高,達到64.87%。除水分外,牦牛皮中蛋白質的含量達到28.17%,占干重質量的80.19%。牦牛皮中的蛋白主要為膠原蛋白,占總蛋白含量的91.91%。而豬皮、鵝皮和魚皮等動物皮中不僅總蛋白含量低于牦牛皮,膠原蛋白含量也僅占總蛋白含量的65%~85%[13-15],由此可見牦牛皮中膠原蛋白的含量十分豐富。

表1 牦牛皮的主要成分Table 1 Main ingredients of yak hide
2.2 不同分子質量膠原抗凍肽的得率及分子質量分布
經過膜分離后,不同分子質量膠原肽的得率如表2所示。可見,熱水耦合酶水解法對膠原肽的得率可達77%。其中,對小分子質量(<1 kDa)膠原肽的得率最高,為61.6%,其余依次為3~5 kDa、>5 kDa和1~3 kDa的膠原肽。不同分子質量膠原肽的分子質量分布如圖2所示,可見其主要為<1 kDa的膠原肽。
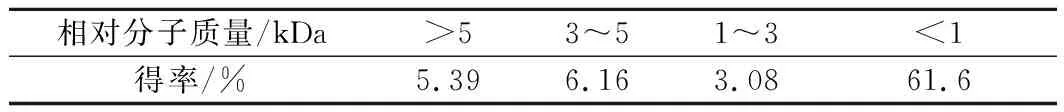
表2 不同分子質量膠原抗凍肽的得率Table 2 Contents of different peptides of collagen antifreeze peptide
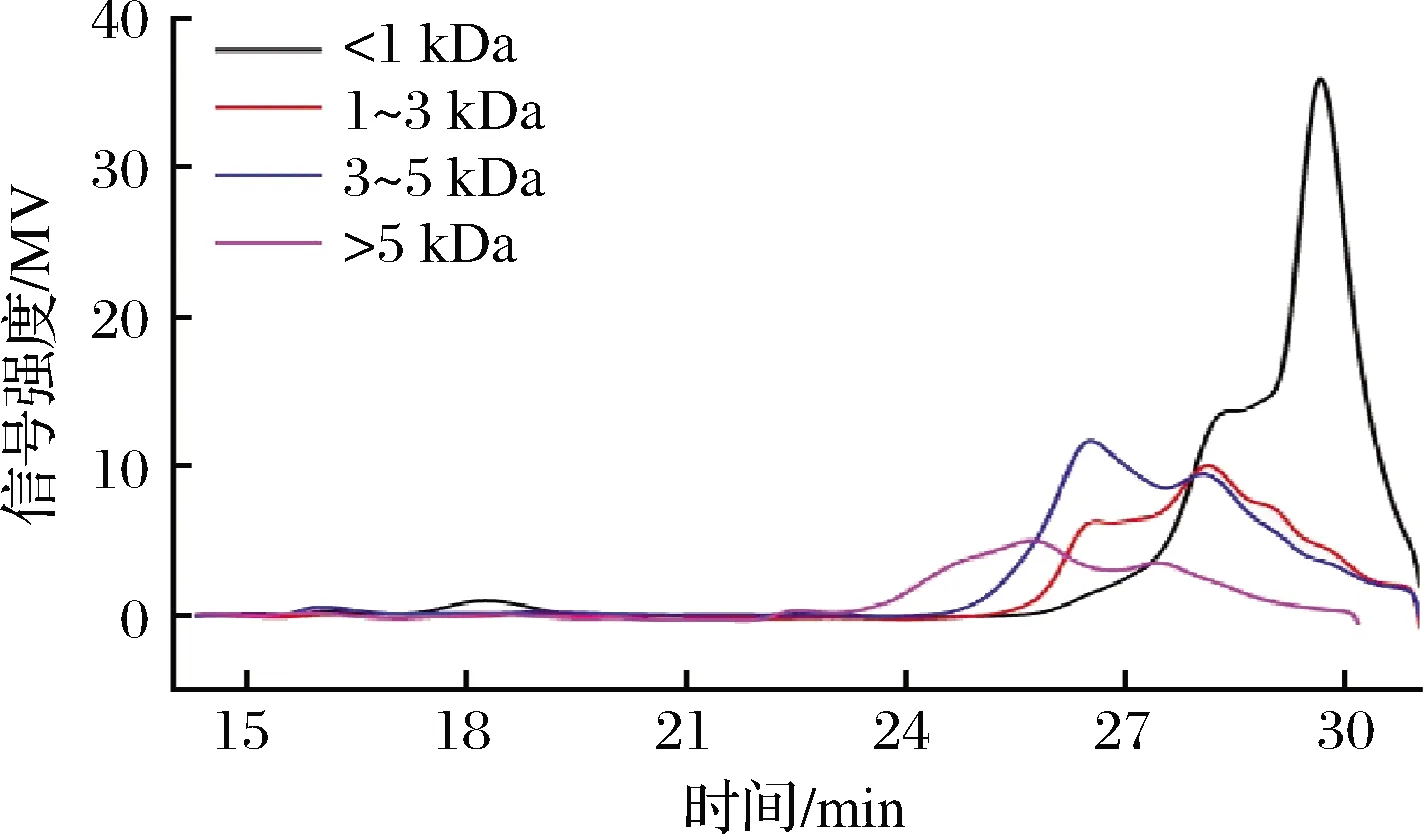
圖2 不同分子質量膠原抗凍肽的分子質量分布圖Fig.2 Molecular weight distribution of collagen antifreeze peptides with different molecular weights
2.3 不同分子質量膠原抗凍肽的氨基酸組成
采用氨基酸自動分析儀分析了不同分子質量膠原肽的氨基酸組成,結果如表3所示。在不同分子質量膠原肽的氨基酸組成中,甘氨酸(Gly)、脯氨酸(Pro)和Hyp含量較高,其中,Gly幾乎占了1/3,Pro含量約占9.6%~10.6%,Hyp含量約占3.4%~5.5%,為典型的膠原蛋白的氨基酸組成模式。膠原蛋白的氨基酸序列通常被描述為-(Gly-X-Y)n-,其中X通常為Pro或Hyp[9]。這一重復序列與GRAHAM等[16]從雪蚤中提取的AFP的重復序列-Gly-X-X-非常相似。
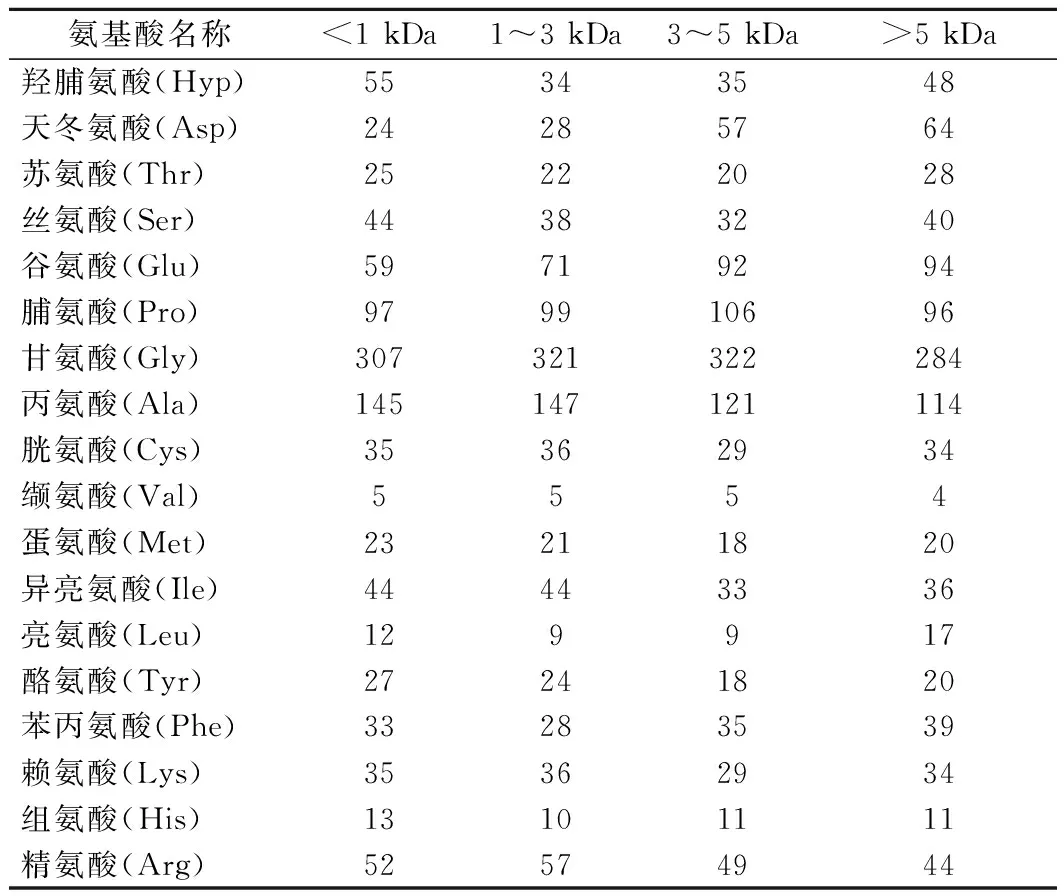
表3 不同分子質量膠原抗凍肽的氨基酸組成 單位:個/1 000個殘基
在4種不同分子質量的膠原肽中,<1 kDa和>5 kDa膠原肽的Hyp含量顯著高于其他分子質量的膠原多肽,分別達到5.5%和4.8%。已有研究發現,Hyp具有親水性,可通過羥基與冰晶棱面結合,從而抑制冰晶的生長,因而其對冰晶的重結晶抑制效應有顯著影響[17]。4種不同分子質量膠原肽中高含量的脯氨酸殘基的烷基側鏈可提供非極性環境從而起到穩定氫鍵的作用[7]。
2.4 不同分子質量膠原抗凍肽的熱滯活性
以無抗凍活性的BSA為對照,采用差示掃描量熱法測定了不同分子質量膠原抗凍肽的熱滯活性,結果如圖3所示。
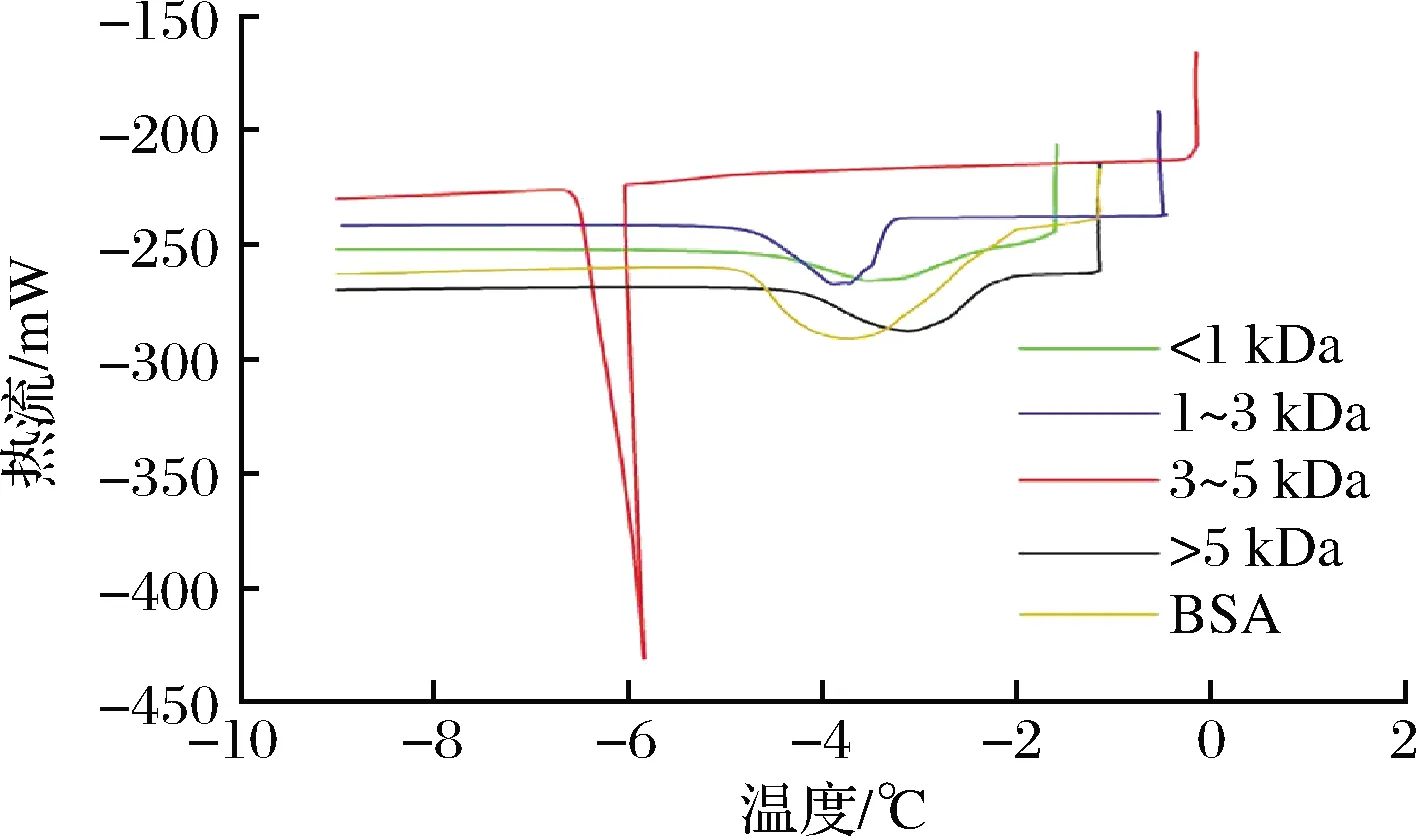
圖3 不同分子質量膠原抗凍肽的熱滯活性Fig.3 Thermal hysteresis activity of collagen antifreeze peptides with different molecular weights
相比較于無抗凍活性的BSA,1~3 kDa和3~5 kDa膠原抗凍肽的結晶峰出現明顯延遲,熱滯活性值分別達到2.66和5.77 ℃。<1 kDa和>5 kDa的膠原抗凍肽表現出輕微的熱滯活性,分別為0.05 和0.92 ℃。熱滯活性較高的1~3 kDa和3~5 kDa膠原抗凍肽的氨基酸組成中Hyp具有較低的比例,這與之前文獻報道的結果一致[18]。AFPs可分為“中等活躍”AFPs和“超級活躍”AFPs,“中等活躍”的AFPs可以使冰點降低不超過1 ℃,與冰晶的相互作用僅限于特定的非基底面;而“超級活躍”的AFPs可以結合多個冰晶面,甚至可使冰點下降3~6 ℃[19]。所以<1 kDa、>5 kDa的膠原抗凍肽可被歸類為中等活躍AFPs,1~3 kDa、3~5 kDa的膠原抗凍肽可被歸類為超級活躍AFPs。
2.5 不同分子質量膠原抗凍肽的重結晶抑制作用
以40%的蔗糖溶液為空白對照,利用低溫顯微鏡觀察了不同分子質量抗凍肽的重結晶抑制作用,冰晶在-50 ℃和-6 ℃保持40 min的狀態如圖4所示。

a~e-40%蔗糖;f~j-<1 kDa的多肽;k~o-1~3 kDa的多肽;p~t-3~5 kDa的多肽;u~y->5 kDa的多肽圖4 不同分子質量膠原抗凍肽的重結晶抑制活性Fig.4 Recrystallization inhibition activity of collagen antifreeze peptides with different molecular weights
在樣品溶液從室溫降溫至-50 ℃的過程中,樣品由液體狀態轉變成由多個單冰晶緊密結合組成的固體狀態,此時顯微形貌變暗(圖4-a)。隨著樣品溫度從-50 ℃緩慢增加至-6 ℃,部分冰晶融化形成彌散狀態(圖4-b)。繼續在-6 ℃保持40 min后,冰晶發生了重結晶現象,主要有融化與消失收縮、保持相對圓形均勻生長和多個小冰晶合并成較大的冰粒[20-22]等3種不同的形式。膠原抗凍肽的添加顯著抑制了冰晶的重結晶,不同樣品抑制重結晶的程度也出現了顯著的差異。在40%的蔗糖溶液中形成的典型圓形冰晶,隨著在-6 ℃保溫時間的延長,仍呈圓形均勻生長,但冰晶顆粒的平均尺寸顯著高于添加膠原抗凍肽的冰晶;添加<1 kDa和>5 kDa 膠原抗凍肽的樣品在-6 ℃保溫40 min后,其冰晶數量及尺寸沒有發生顯著變化,冰晶形狀多呈有棱角狀和拉長型;而添加1~3 kDa和3~5 kDa膠原抗凍肽樣品中的冰晶在-6 ℃保溫40 min后,部分冰晶融化和消失導致冰晶數量減少,冰晶形狀包括圓形、有棱角型、拉長型,其中圓形占有較大比例。可見,<1 kDa和>5 kDa的膠原抗凍肽表現出了更好的重結晶抑制活性,這可能與<1 kDa和>5 kDa的膠原抗凍肽具有較高含量的Hyp有關。1~3 kDa和3~5 kDa的膠原抗凍肽雖具有較高的熱滯活性,但抑制重結晶活性的效果并不佳,表明熱滯活性與重結晶抑制活性并不呈正相關,這與YU等[23]的研究結果一致。
膠原抗凍肽結合于冰晶表面的特定區域進而抑制了這些方向晶體的生長,這可以作為解釋冰晶形態發生變化的原因。膠原抗凍肽抑制冰晶重結晶的機理則可以通過“石頭壓海綿”模型來進行解釋。如圖5所示,白色圓圈代表AFPs,大面積的黑色部分代表冰晶,黑色小點代表游離的水。在冰晶重結晶過程中,AFPs附著在冰晶表面,好像“石頭”壓在“海綿”上,從而阻礙了冰晶平滑曲面的生長,游離的水只能凍結/附著在AFPs附著點之間的冰晶上,冰晶在AFPs附著點之間的位置凸起生長,使冰晶表面彎曲造成局部表面的曲率變大,進而通過Gibbs-Thompson效應或Kelvin效應降低冰晶的局部冰點,晶體停止生長[24-25]。
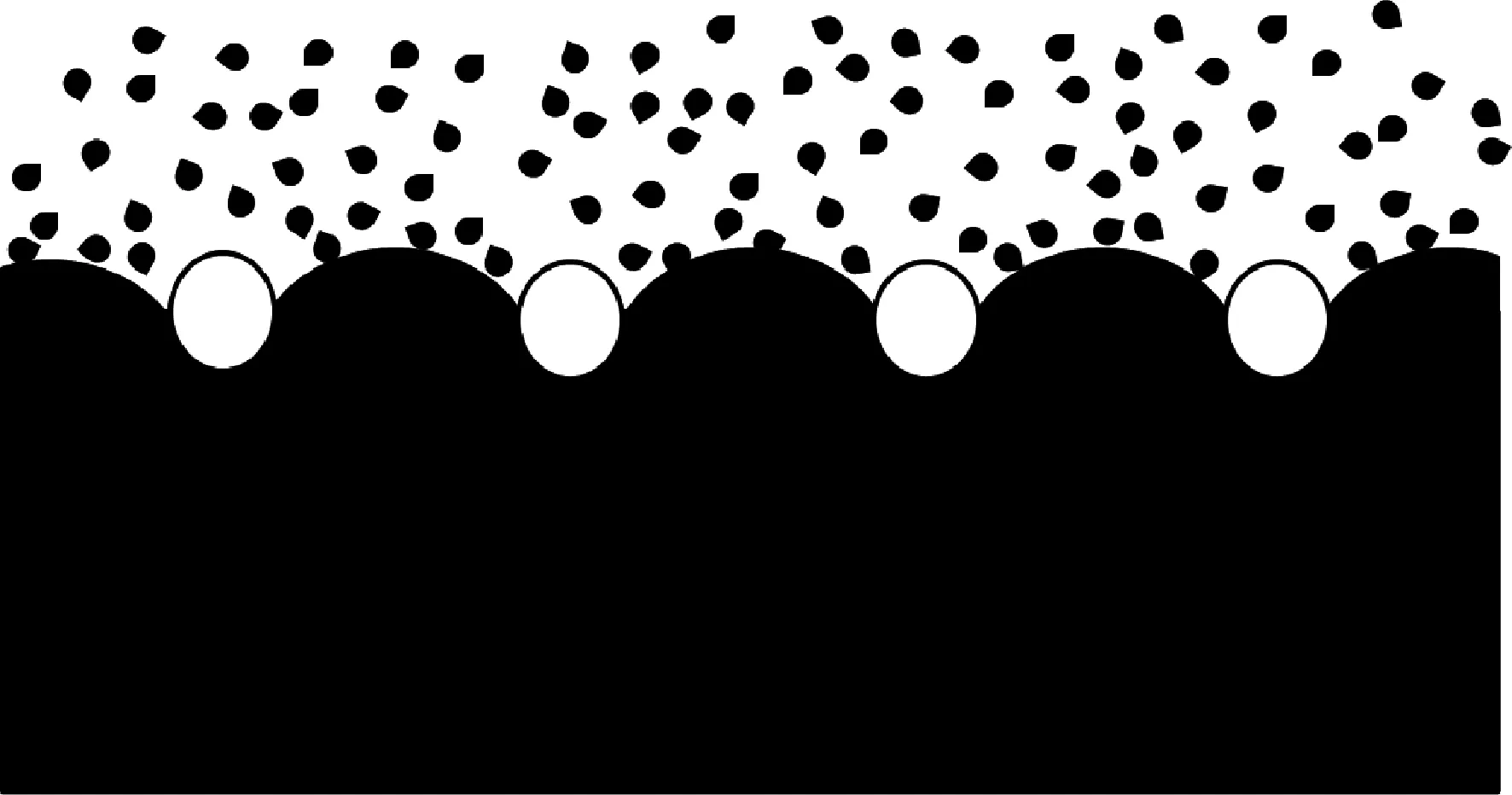
圖5 AFPs抑制重結晶機理圖Fig.5 Mechanism of AFPs inhibiting recrystallization
2.6 不同分子質量膠原抗凍肽對單冰晶生長形態的修飾效應
以40%的蔗糖溶液為空白對照,利用低溫顯微鏡觀察了不同分子質量膠原抗凍肽修飾單冰晶生長的作用,結果如圖6所示。
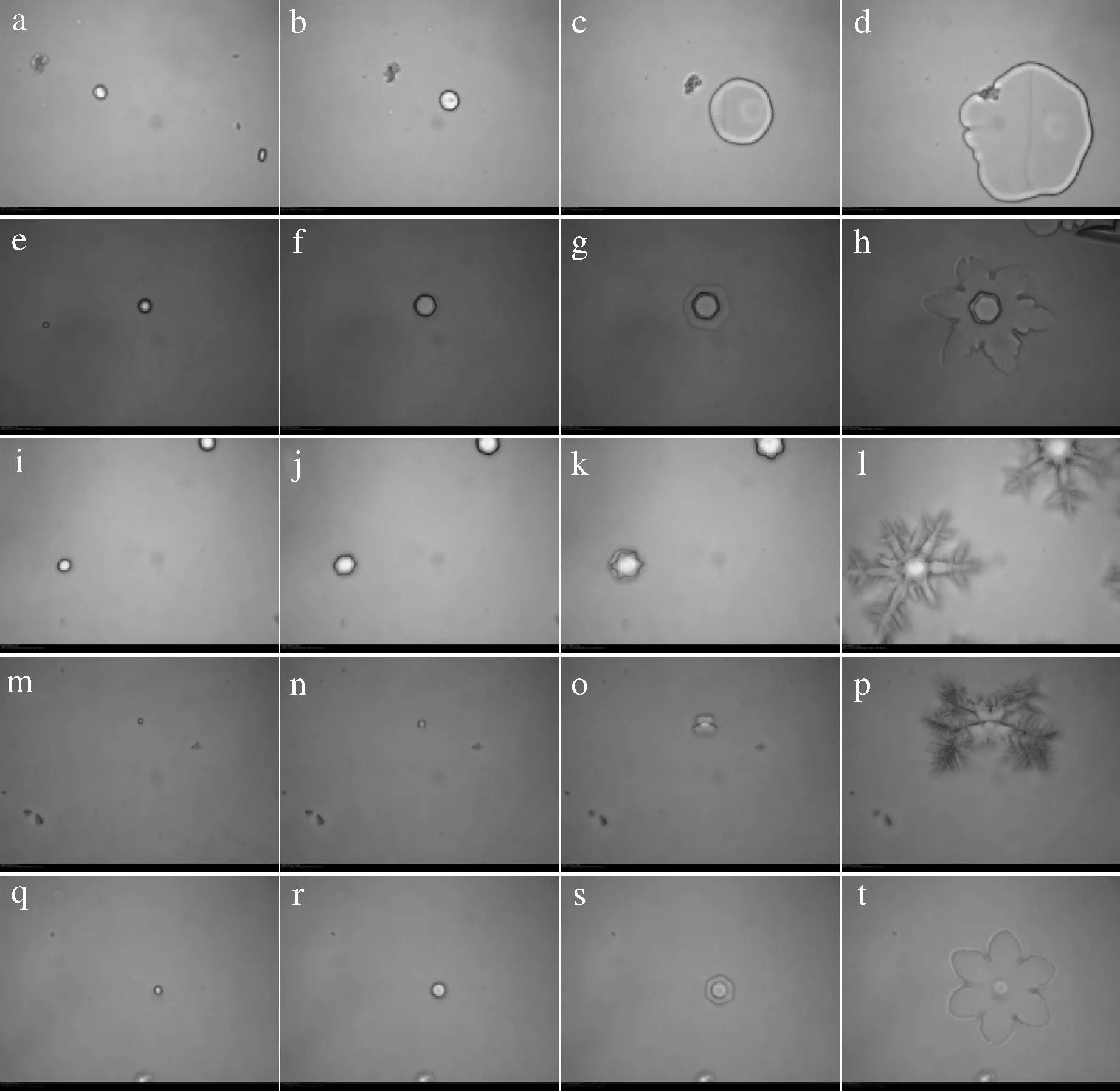
a~d-40%蔗糖;e~h-<1 kDa的多肽;i~l-1~3 kDa的多肽;m~p-3~5 kDa的多肽;q~t->5 kDa的多肽圖6 不同分子質量膠原抗凍肽對單冰晶生長形態的修飾作用Fig.6 Effects of collagen antifreeze peptides with different molecular weights on growth morphology of single ice crystals
由圖6可見,隨著樣品溫度的緩慢降低,40%蔗糖溶液中的冰晶始終保持圓形的形態,冰晶面積迅速生長擴大(圖6-a~圖6-d);而添加了膠原抗凍肽的樣品中(圖6-e~圖6-t),冰晶最初以六角型生長,在經過冰點時,冰晶在6個方向上迅速生長“爆裂”,其中<1 kDa和>5 kDa膠原抗凍肽導致冰晶呈六角星型擴張,而1~3 kDa和3~5 kDa膠原抗凍肽導致冰晶呈樹突狀擴張[26]。添加不同分子質量膠原抗凍肽的冰晶在經過冰點溫度時出現的形狀差異是其熱滯活性的顯著差別導致的。<1 kDa和>5 kDa的膠原抗凍肽為中等活躍的AFPs,會導致冰晶沿平行于c軸的方向爆發,而1~3 kDa和3~5 kDa的膠原抗凍肽為高度活躍的AFPs,會導致冰晶垂直于c軸爆發。在昆蟲[12]和細菌[27]的AFPs中可以觀察到類似的冰晶生長模式。
采用IMAGE-PRO PLUS軟件計算了不同分子質量膠原抗凍肽溶液中單個冰晶在緩慢降溫時的生長面積值(μm2)與時間(s)的關系,它們的斜率表示冰晶的生長速率。由圖7可見,40%蔗糖、<1 kDa、1~3 kDa、3~5 kDa和>5 kDa膠原抗凍肽溶液中的冰晶面積隨著時間生長的斜率值分別為395.36、185.82、266.42、149.50和180.60 μm2/s。很顯然,3~5 kDa膠原抗凍肽溶液中冰晶生長的斜率值最低,表明添加3~5 kDa的膠原抗凍肽能夠顯著延緩冰晶的生長。這可能是由于3~5 kDa膠原抗凍肽具有最高的熱滯活性,其與冰晶的多個平面,包括基底面都具有較強的親和力,從而能夠更均勻地與整個冰面結合,進而抑制冰晶的生長。
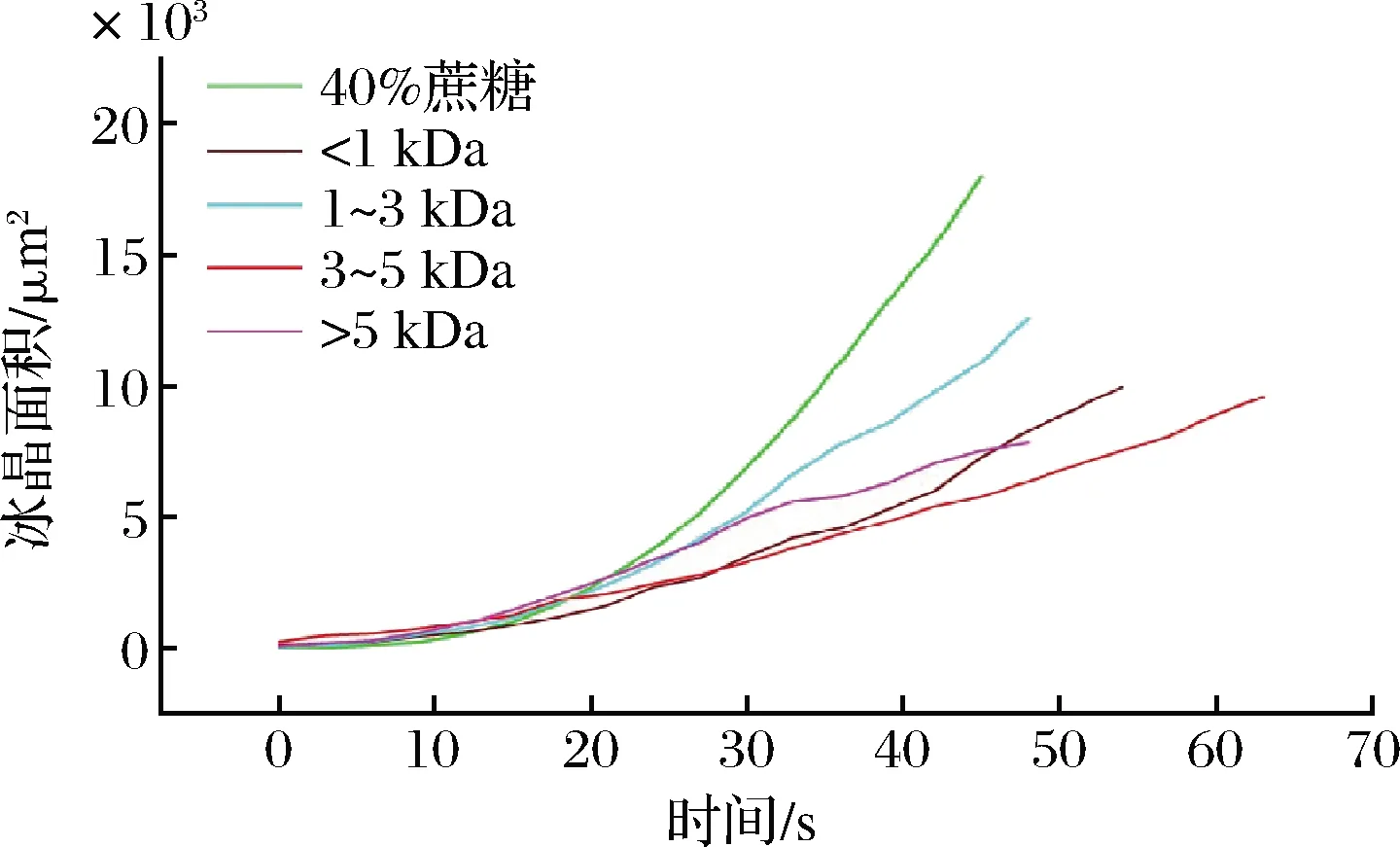
圖7 不同分子質量膠原抗凍肽對單個冰晶生長速率的影響Fig.7 Effect of antifreeze peptides with different molecular weights on growth rate of individual ice crystals
膠原抗凍肽的-Gly-X-Y-重復序列中存在分子質量較小的氨基酸殘基,由于沒有構象/空間約束,這些氨基酸殘基中的羰基氧原子可與冰核中的氧-氧鍵在同一個肽平面上排列。冰核中2個氧原子之間距離為4.52 ?,與三肽重復序列中氧原子距離4.538 ?十分接近,因此肽鏈中形成的氧平面可通過氫鍵相互作用與冰晶表面形成互補的結構,使膠原抗凍肽吸附在冰晶表面[28-29]。冰面在膠原抗凍肽吸附點之間的位置生長,造成表面曲率增加進而引起平衡蒸氣壓的移動,導致冰晶局部凍結溫度非平衡性下降。當表面達到一個特征曲率時冰晶停止生長,此時冰與水之間的蒸汽壓處于平衡狀態,不發生額外的結晶。繼續降低溫度使特征曲率增加,當溫度到達滯回冰點時,曲率達到了最大凸度,進一步冷卻導致冰晶體的“爆發”,即以一種看上去爆炸性的方式生長。
3 結論
本文主要研究了<1 kDa、1~3 kDa、3~5 kDa和>5 kDa等4種不同分子質量膠原抗凍肽的熱滯活性、重結晶抑制能力及修飾單冰晶生長的能力。結果表明,不同分子質量膠原抗凍肽均具有調節冰晶生長和熱滯的能力。其中3~5 kDa的膠原抗凍肽熱滯活性最高,表明其可以顯著降低結冰點,能夠在低溫度下保護樣品的生理功能。<1 kDa和>5 kDa的膠原抗凍肽重結晶抑制活性最好,表明其可保護樣品組分不受大冰晶的侵害。4種不同分子質量的膠原抗凍肽均具有修飾冰晶生長形態的效應,其中3~5 kDa的膠原抗凍肽具有最強地抑制單冰晶生長的能力。綜上可見,膠原抗凍肽的分子質量分布對其冰結構活性至關重要,因而在實際應用過程中,可以通過調整酶解參數控制膠原抗凍肽的分子質量,使之滿足于不同的需求。
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
產品可靠性報告(2017年7期)2017-09-05 09:49:12
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
