太子參NF-Y基因家族鑒定及表達分析
2021-07-21 01:31:10王崇敏劉曉清
西南大學學報(自然科學版) 2021年8期
李 軍,周 濤,王崇敏,劉曉清
貴州中醫藥大學 中藥民族藥資源研究院,貴陽 550025
核因子Y(nuclear factor Y,NF-Y),又稱CBF或亞鐵血紅素激活蛋白HAP,主要由NF-YA,NF-YB及NF-YC 3個亞家族組成[1].NF-Y蛋白家族的不同成員可通過不同途徑進入細胞核,其中NF-YA是由Importin β介導輸入核中,而含有HFM亞基的NF-YB和NF-YC作為異二聚體復合物則由Importin 13介導進入細胞核[2].3個亞家族可以單獨調控基因表達,也可形成NF-YB/NF-YC CONSTANS或NF-Y復合體,進一步結合CCAAT元件參與基因表達的調控[3].
在哺乳動物及酵母體內,NF-Y的3個亞基均由單基因編碼[4],但植物中每個亞基均由基因家族編碼[5],提供了更多的 NF-YA/NF-YB/NF-YC 三聚體組合,增加了核因子Y功能的復雜性,參與了在種子萌發、胚胎發育、開花過程以及植物對氮、磷、脫落酸(ABA)以及鹽分脅迫等的響應過程[6].研究表明大豆GmNF-YCa在蔗糖和甘露醇誘導后上調表達,過表達可顯著改善甘露醇和蔗糖脅迫下擬南芥種子的發芽率、根長和側根伸長明顯等[7].谷子SiNF-YA6主要在根部表達,同時受低氮、干旱、高鹽和 ABA 等脅迫的誘導表達,該基因的過表達顯著提高了擬南芥對低氮脅迫的耐受性[8].由此可見,NF-Y蛋白家族的不同成員不僅參與了植物多種生長發育進程,且在植物抵御逆境脅迫等方面具有重要作用.
太子參為傳統中藥,系石竹科植物孩兒參Pseudostellariaheterophylla(Miq.)Paxex Pax et Hoffm的干燥塊根,含有多糖類、皂苷類、磷脂類、環肽類等活性成分,具有增強免疫力、降血糖、抗疲勞等多種功效[9].隨著近年來太子參需求量的逐年上升,逐步轉向人工種植,主要分布于溫暖濕潤的氣候環境中[10].高溫、干旱等逆境容易引起太子參生長發育受阻,從而導致減產、病害等問題.因此,研究NF-Y基因家族的表達模式及對干旱脅迫的響應,對于深入理解其在太子參生長發育過程中的功能和作用,解析NF-Y蛋白在植物抵抗逆境脅迫中的作用機制具有重要意義.
1 材 料
供試材料為貴州施秉常用種,轉錄組數據由上海美吉生物醫藥科技有限公司測序完成.數據庫1:采用課題組前期構建的太子參轉錄組數據庫(葉、莖、根表皮、根木質部)[11];數據庫2:參照路丙社等[12]的方法,設置30%,44%,58%,72%,86%及100%的土壤水分梯度,相對含水量=土壤含水量/田間持水量×100%,取塊根進行轉錄組測序,de Novo組裝;數據庫3:采用0.2 mg/L NAA(萘乙酸)誘導組培苗生根[13],然后轉入MS+45 g/L蔗糖,MS+60 g/L蔗糖及MS+75 g/L蔗糖的培養基中培養,取塊根進行轉錄組測序,de Novo組裝.
2 方 法
2.1 NF-Y基因家族的鑒定
從NCBI中下載甜菜(BetavulgarisL.)、擬南芥(Arabidopsisthaliana)、水稻(OryzasativaL.)等的NF-Y蛋白家族成員.通過BlastP分別比對3個數據庫,從中鑒定太子參NF-Y家族同源序列,期望值設為小于0.001,結合數據庫原有的注釋結果,剔除注釋錯誤的序列,將3個數據庫中鑒定的NF-Y蛋白序列統一編號.
2.2 系統進化樹的構建及蛋白保守序列比對
采用muscle 3.8軟件(http://www.drive5.com/muscle/)對太子參及甜菜、擬南芥及水稻進行多序列比對,分析結果用MEGA X(https://www.megasoftware.net/)分析,采用鄰接法(neighbor-joining method)構建NF-Y的系統進化樹,檢驗參數Booistrap重復1 000次.利用MEME在線程序(http://meme-suite.org/)對NF-Y蛋白家族的保守基序進行分析,預測太子參NF-Y蛋白家族的蛋白質保守基序(Motif),參數設置為默認值,并用WebLogo軟件(http://weblogo.berkeley.edu/logo.cgi)展示保守基序.
2.3 表達分析
采用RSEM軟件(http://deweylab.biostat.wisc.edu/rsem/)進行基因表達分析,采用RPKM值(reads per kb permillion reads)表示基因的相對表達水平,采用Cluster 3.0軟件對NF-Y基因的表達水平進行標準化,最后采用Heatmap圖展示基因的表達.
2.4 蛋白互作分析
采用美吉生物云平臺(https://cloud.majorbio.com/)中的STRING數據庫(http://string-db.org/)對NF-Y蛋白互作進行分析.參考擬南芥的蛋白互作關系,構建太子參NF-Y蛋白的互作關系網絡.采用confidence score值評價互作關系的可信度,默認值0.4,值越大代表互作關系越可靠.
3 結 果
3.1 太子參NF-Y家族基因的鑒定與生物信息學分析
采用擬南芥、水稻等模式植物的NF-Y蛋白檢索3個太子參轉錄組數據庫,結合注釋信息及手工分析,共鑒定出24個NF-Y基因(表1),分別歸入3個亞家族,包括9個NF-YA,10個NF-YB及5個NF-YC.其中數據庫1中獲得9個NF-YA,6個NF-YB及4個NF-YC;數據庫2中獲得8個NF-YA,8個NF-YB及4個NF-YC;數據庫3中獲得8個NF-YA,7個NF-YB及4個NF-YC.PhNF-YA8僅在數據庫1中檢索到,PhNF-YB5,PhNF-YB6,PhNF-YB7,PhNF-YB9及PhNF-YC5未在數據庫1中檢索到,暗示這些基因可能是誘導型基因.
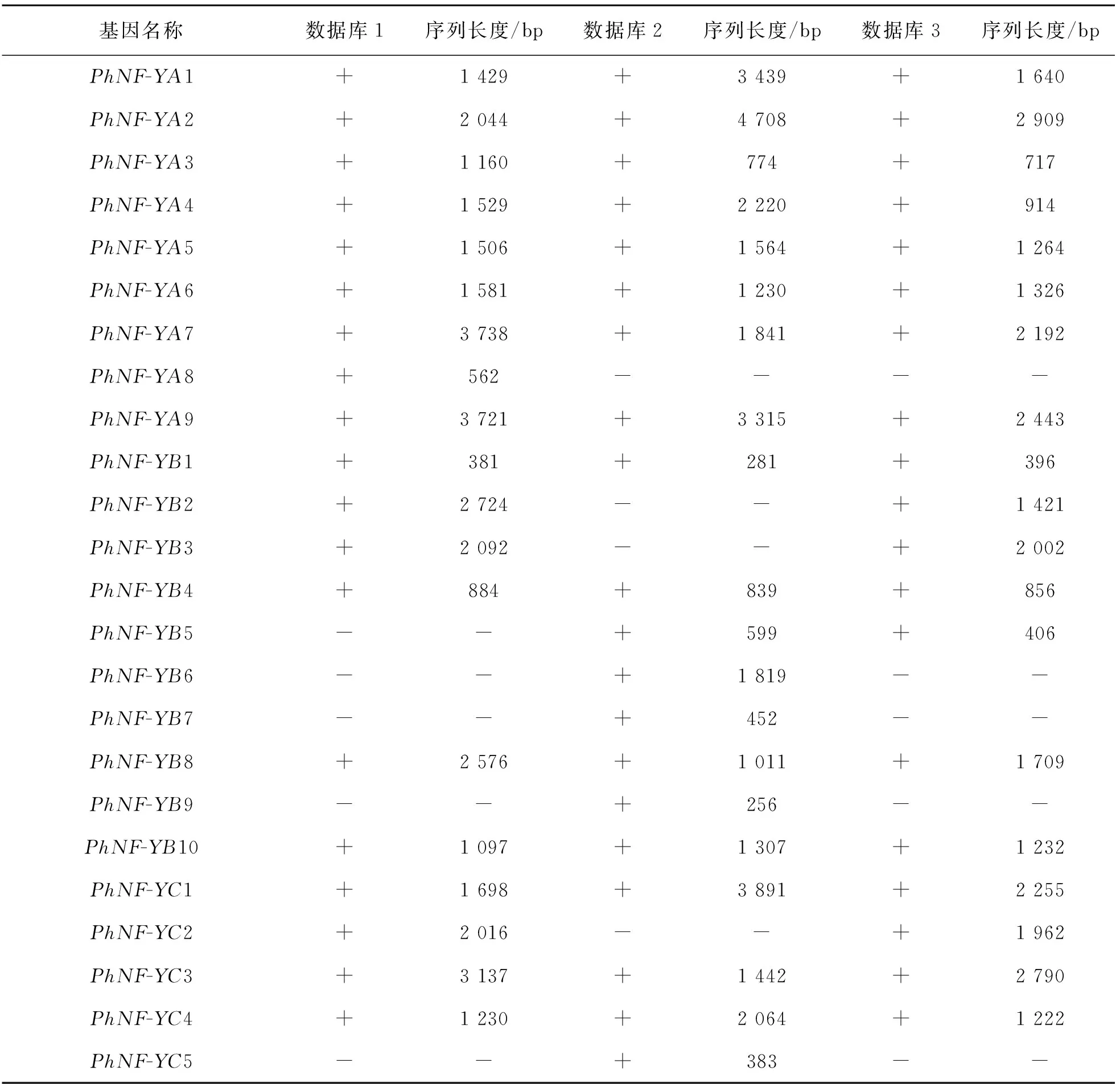
表1 太子參NF-Y基因家族的篩選與鑒定
3.2 太子參NF-Y家族基因蛋白的保守基序及進化分析
為了推測24個太子參NF-Y蛋白的功能,分別構建了NF-YA,NF-YB及NF-YC氨基酸序列的系統進化樹(圖1).太子參的9個NF-YA蛋白分別歸入3個類群,其中PhNF-YA2,PhNF-YA3,PhNF-YA7,PhNF-YA9與AtNF-YA3,AtNF-YA5,AtNF-YA6,AtNF-YA8歸為第1類群,PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA8與AtNF-YA1,AtNF-YA9歸為第2類群,PhNF-YA6與AtNF-YA2,AtNF-YA10歸入第3類群.
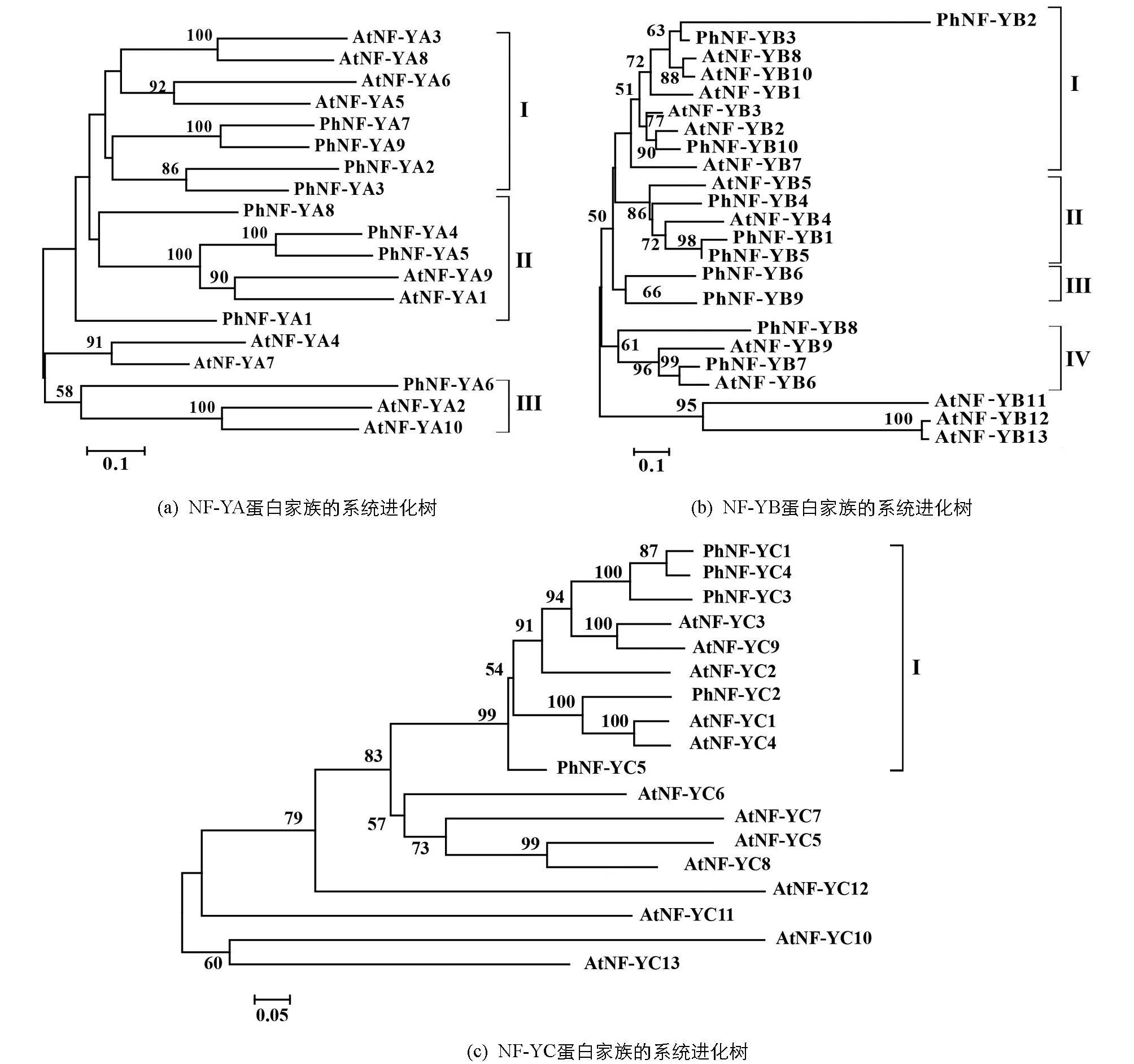
圖中分支點的數字表示基于1 000次重復該節點的自展支持率;標尺代表遺傳距離.
太子參的10個NF-YB蛋白分別歸入4個類群,其中PhNF-YB2,PhNF-YB3,PhNF-YB10與AtNF-YB1,AtNF-YB2等6個擬南芥NF-YB蛋白歸入第1類群,PhNF-YB1,PhNF-YB4,PhNF-YB5與AtNF-YB4,AtNF-YB5歸入第2類群,PhNF-YB6及PhNF-YB9歸入第3類群,PhNF-YB7,PhNF-YB8與AtNF-YB6,AtNF-YB9歸入第4類群.太子參的5個PhNF-YC蛋白歸入同一個類群.
為了解太子參NF-Y蛋白的序列特點,分析了太子參NF-Y蛋白的保守基序(圖2),發現PhNF-YA,PhNF-YB及PhNF-YC亞家族中均具有一段高度保守的序列,暗示這些序列與太子參NF-Y蛋白的功能密切相關.

圖2 太子參NF-Y蛋白的保守基序
3.3 不同組織中NF-Y家族基因的表達分析
為了進一步闡明太子參NF-Y蛋白的功能,分析了這些基因在葉、莖、塊根皮部及塊根木質部中的表達模式(圖3a),結果顯示9個PhNF-YA基因中PhNF-YA4,PhNF-YA5,PhNF-YA7,PhNF-YA9僅在葉中有較高表達,PhNF-YA2,PhNF-YA6在葉及莖中均有較高表達,PhNF-YA1在塊根木質部有較高表達,PhNF-YA3在塊根皮部有較高表達.6個PhNF-YB基因中PhNF-YB1和PhNF-YB4在塊根皮部及莖中有較高表達,PhNF-YB10僅在葉中有較高表達,PhNF-YB3在塊根木質部中有較高表達,PhNF-YB8在塊根木質部及葉中有較高表達.4個PhNF-YC基因中PhNF-YC2在葉中有較高表達,其余基因表達量較低.19個基因的蛋白互作關系分析結果顯示(圖3b),PhNF-YA4及PhNF-YC2等9個PhNF-Y蛋白的互作關系在太子參葉片的生長發育過程中具有重要作用.
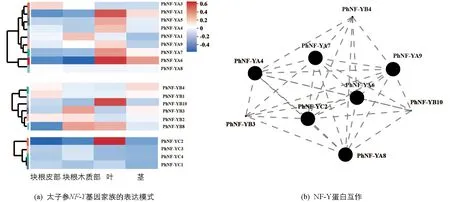
圖3 太子參NF-Y基因家族在不同組織中的表達及蛋白互作
3.4 塊根中NF-Y家族基因對土壤水分脅迫的響應
為了研究太子參NF-Y蛋白對水分脅迫的響應機制,分析了不同土壤水分脅迫下塊根中NF-Y基因的表達模式(圖4a),結果顯示8個PhNF-YA基因中PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA7,PhNF-YA9在干旱脅迫下(30%)有較高表達,PhNF-YA1,PhNF-YA2,PhNF-YA3在低度干旱條件下(58%)有較高表達,PhNF-YA6隨著土壤水分含量的上升表現出上調的趨勢.10個PhNF-YB基因中PhNF-YB2,PhNF-YB3及PhNF-YB8在干旱脅迫下(30%)有較高表達,PhNF-YB10在適度干旱脅迫下(44%)有較高表達,PhNF-YB1在低度干旱條件下(58%)有較高表達.5個PhNF-YC基因中PhNF-YC1,PhNF-YC3,PhNF-YC4在干旱脅迫下(30%)有較高表達,PhNF-YC2與土壤水分的增加趨勢基本一致,PhNF-YC5僅在澇滯環境下(100%)有較高表達.23個PhNF-Y蛋白的互作關系表明(圖4b),PhNF-YA1,PhNF-YB4,PhNF-YC3及PhNF-YC4等11個PhNF-Y蛋白積極響應土壤水分的脅迫,調控植物的生長發育.
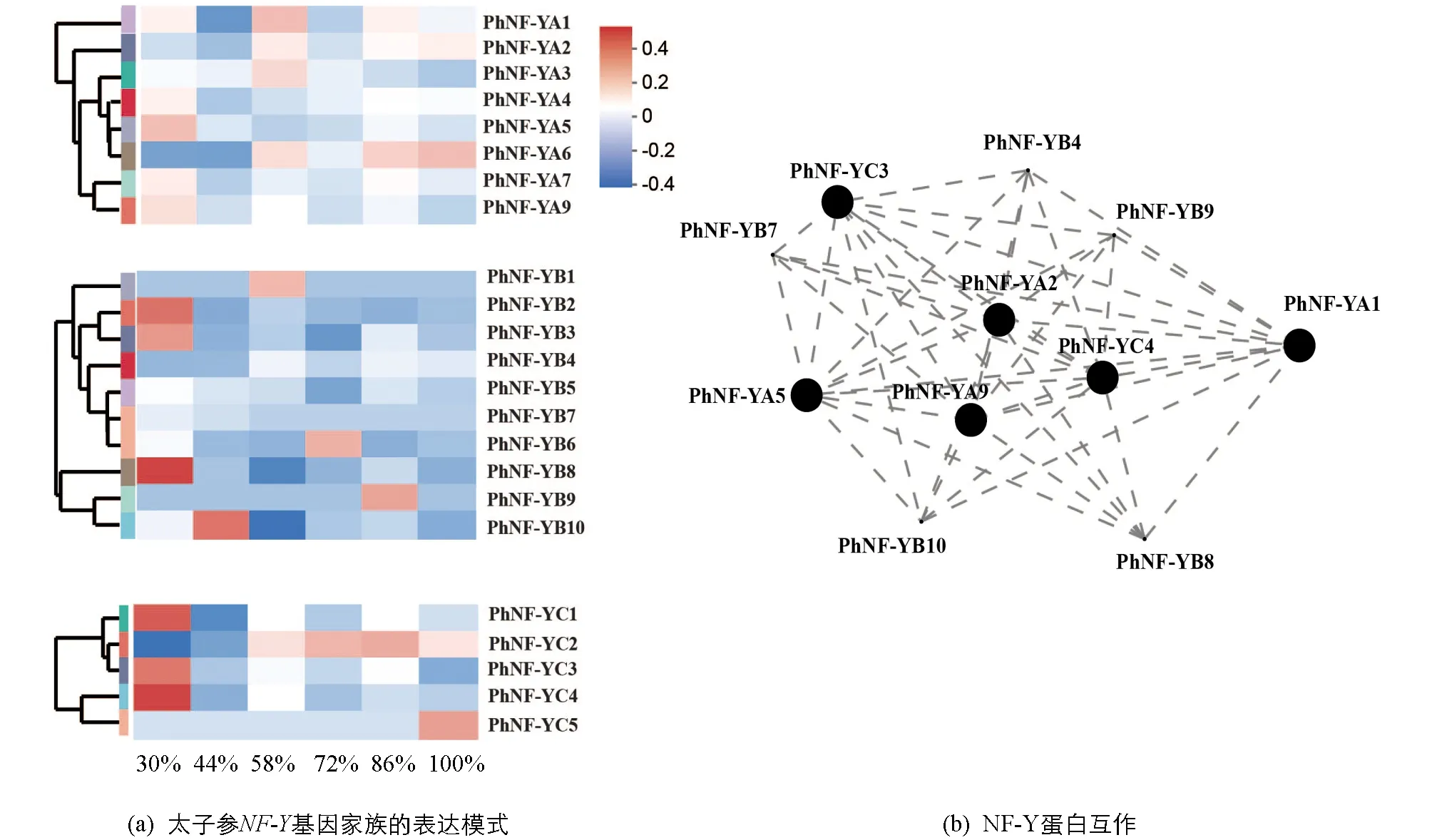
圖4 太子參塊根中NF-Y基因家族在土壤水分脅迫下的表達及蛋白互作
3.5 根中NF-Y家族基因對蔗糖脅迫的響應
為了研究太子參NF-Y蛋白對蔗糖誘導的響應機制,利用組培苗觀察不同蔗糖濃度培養條件下太子參根部NF-Y蛋白的表達模式(圖5a),結果顯示,隨著培養基中蔗糖濃度的升高,3個PhNF-YA基因的表達呈現逐漸升高的趨勢,7個PhNF-YB基因中PhNF-YB5的表達量隨蔗糖濃度的升高而升高,PhNF-YB1,PhNF-YB2的表達呈先上升后下降的趨勢,PhNF-YB3,PhNF-YB4,PhNF-YB8,PhNF-YB10的表達呈先下降后上升的趨勢.4個PhNF-YC基因中PhNF-YC3及PhNF-YC4基因的表達受高濃度蔗糖誘導,PhNF-YC1在高濃度蔗糖條件下表達受到抑制.19個PhNF-Y蛋白的互作關系表明(圖5b),PhNF-YA1,PhNF-YB4,PhNF-YC2等10個PhNF-Y蛋白積極響應蔗糖脅迫,協同調控植物的生長發育.
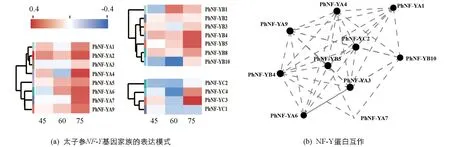
圖5 太子參根中NF-Y基因家族在蔗糖誘導下的表達及蛋白互作
4 討 論
已有證據表明NF-Y蛋白在葉綠體形成及光合作用中具有重要作用.擬南芥AtNF-YA5,AtNF-YB9和AtNF-YC9形成的復合體可與GCR1,GPA1和Pirin1組成信號傳遞鏈,參與調控植株中葉綠素a/b結合蛋白基因的表達[14].過量表達小麥TaNF-YB3后顯著提高葉綠素含量、光合速率及早期生長率[15].水稻Os-NF-YB2/3/4可影響多個核酮糖-1,5-二磷酸羧化酶/氧合酶小亞基和葉綠素a/b結合蛋白基因的表達[16].本研究發現太子參PhNF-YA4,PhNF-YA5等6個基因,PhNF-YB8,PhNF-YB10及PhNF-YC2在葉中高量表達,且可能具有復雜的相互作用關系,暗示這些基因在協同調控葉的生長發育及光合作用中具有重要作用,但具體機制尚不明確.擬南芥NF-YA1/5/6/9共同調控雄配子發育、胚胎發育、種子形態及萌發等[17-18],單個基因或兩個基因的突變并不能發生表型的改變,但過表達其中任一基因均能影響種子的發育,暗示了太子參PhNF-YA亞家族也有功能冗余的可能性.此外,PhNF-YA1及PhNF-YB3,PhNF-YB8在塊根木質部中有較高表達,且進化分析結果顯示PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA8與AtNF-YA1,AtNF-YA9聚為一類,暗示了這些太子參NF-Y基因可能與太子參塊根的發育有一定關系.
NF-Y蛋白在水分脅迫過程中具有重要作用.研究表明擬南芥大部分NF-YA基因呈上調表達,而小麥中NF-YA則相反[19-20].過表達AtNF-YB1和ZmNF-YB2可顯著減少轉基因植株的葉片水分流失及提高抗旱能力[21].部分NF-Y基因的轉錄受ABA途徑調控,轉錄后受到miR169的調控[22].本研究中太子參PhNF-YA1,PhNF-YA4,PhNF-YA5,PhNF-YA7,PhNF-YA9,PhNF-YB2,PhNF-YB3,PhNF-YB8及PhNF-YC1,PhNF-YC3,PhNF-YC4在干旱脅迫下均顯著上調,且進化分析結果顯示PhNF-YB2,PhNF-YB3,PhNF-YB10與AtNF-YB1,AtNF-YB2等6個擬南芥NF-YB蛋白歸入一類,暗示了這些太子參NF-YB蛋白與抗旱有密切聯系.此外,蛋白間互作關系暗示PhNF-YC3,PhNF-YC4在干旱脅迫響應過程中可能具有核心調控作用.值得注意的是,PhNF-YA6及PhNF-YC2的表達隨土壤水分的增加而增加,揭示了這些基因在太子參感知土壤水分變化的過程中具有重要作用.
蔗糖及其衍生物不僅是光合同化物與能量的運輸和貯藏形式,也是植物中一種重要的信號分子,調控基因表達及影響器官的生長發育[23-24].本研究分析了太子參NF-Y蛋白對蔗糖脅迫的響應,發現了大部分基因能夠響應蔗糖濃度的變化,暗示了環境因子可通過影響組織中蔗糖的分布來調控NF-Y蛋白的表達,進而影響器官的形態建成.
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
財經(2017年15期)2017-07-03 22:40:49
財經(2017年2期)2017-03-10 14:35:35
山東工業技術(2016年15期)2016-12-01 05:31:22
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
