肝X受體調控脂代謝研究進展
2021-07-30 02:15:52韓汶潤張丹丹杜曉鸝董鵬志
中國藥理學與毒理學雜志 2021年5期
韓汶潤 ,張丹丹,朱 彥,杜曉鸝,董鵬志
(1.天津中醫藥大學組分中藥國家重點實驗室,天津 301608;2.天津國際生物醫藥聯合研究院中藥新藥研發中心,天津 300457)
膽固醇是維持細胞膜流動性和產生各種激素及膽汁酸的重要脂質。因此,維持膽固醇穩態(包括吸收、輸運、生物合成和外排)至關重要。膽固醇穩態的失調可能導致各種疾病,如動脈粥樣硬化[1]等。肝X受體(liver X receptor,LXR)是膽固醇和脂質代謝的重要調節因子。在動物模型中,LXR活化可減緩心血管疾病并減少動脈粥樣硬化的發生[2]。動脈粥樣硬化發展中的一個關鍵事件是巨噬細胞募集到血管壁的內皮層及巨噬細胞對氧化和(或)修飾膽固醇的過量攝取。巨噬細胞不斷積累氧化和(或)修飾的膽固醇及相關的炎癥反應,導致泡沫細胞形成并促進動脈粥樣硬化早期發展。逆轉巨噬細胞膽固醇積累的過程和抑制血管壁炎癥反應被認為是潛在的治療動脈粥樣硬化的新策略[3]。膽固醇逆轉運(reverse cholesterol transport,RCT)可促進巨噬細胞內膽固醇清除,減少泡沫細胞形成,從而阻斷動脈粥樣硬化發生。LXR可促進RCT,對體內脂質代謝平衡的調節十分重要。LXR除具有改善脂代謝的作用以外,還參與調節多種炎癥因子及炎癥介質的表達,同時調節體內糖代謝過程,發揮抗動脈粥樣硬化作用。LXR的多重脂調節功能揭示了開發LXR配體預防和治療心血管疾病的廣泛前景。但由于LXR激動劑易引發肝脂肪變性和高甘油三酯血癥等不良反應而限制了其發展。因此,需要尋求新的防治策略,如開發LXRβ激動劑和組織特異性激動劑等。為此,本文就LXR信號通路涉及的分子及其在調控脂代謝中的作用予以綜述,并對LXR的天然產物配體及合成配體進行簡要總結。
1 肝X受體概述
1.1 結構
LXR是核受體家族成員之一,最早在1995年由Willy等[4]從肝cDNA文庫中分離得到,因其在肝中表達豐富而得名。LXR具有典型的核受體結構,由4個結構域組成:①N端刺激轉錄的非配體依賴性功能活化域;②DNA結合域;③配體結合和受體二聚化所需的疏水配體結合域(ligand bound domain);④C端配體依賴性反式激活序列,也稱為轉錄活化域[5](圖1)。LXR包括LXRα和LXRβ 2種亞型,二者在DNA和配體結合域的氨基酸同源性約為77%[6]。LXRα主要在代謝活躍的肝、腎、小腸、脾及脂肪組織和巨噬細胞中高表達,其中在肝組織表達最高;而LXRβ在全身多種組織中均有表達[7]。
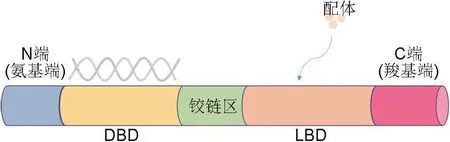
圖1 肝X受體(LXR)的核受體結構.DBD:DNA結合域;LBD:配體結合域.
1.2 轉錄激活
LXR屬于核受體Ⅱ類亞家族,需要與類視黃醇X受體(retinoid X receptor,RXR)形成LXR/RXR異二聚體才能發揮其調控功能。該異二聚體可被LXR與RXR的配體單獨或共同激活,共同激活時起到協同誘導作用[8]。LXR/RXR異二聚體與靶基因啟動子區域內的特定DNA序列即LXR反應元件(LXR response element,LXRE)結合,調節下游基因表達(圖2)。LXRE是由4個核苷酸隔開的2個保守的六核苷酸序列(5′-AGGTCA-3′)以直接重復的方式排列,稱為直接重復元件4(direct repeat 4)[6]。LXR的轉錄活性與LXR/RXR異二聚體密切相關,在無配體的情況下,LXR/RXR異二聚體與輔阻遏物結合時仍與目標基因的啟動子區域結合,從而抑制了目標基因的激活[9]。當配體與LXR或RXR結合后,LXR/RXR異二聚體的構象發生改變,導致輔阻遏物如核輔阻遏物1(nuclear co-repressor 1)及視黃酸和甲狀腺激素受體沉默介體(silencing mediator of retinoic acid and thyroid hormone receptor,也稱為核輔阻遏物2)解離,并募集輔助激活因子,如受體輔助激活因子1(receptor co-activator 1)和激活信號輔助因子2(activating signal co-integrator 2),從而啟動轉錄活性[10]。
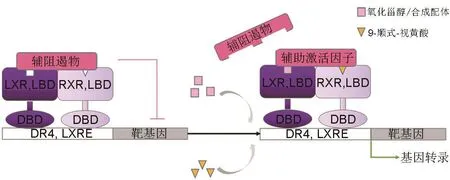
圖2 肝X受體的轉錄激活.DR4:直接反應元件4;LXRE:LXR反應元件;RXR:類視黃醇X受體.
2 肝X受體靶基因及其功能
LXR是調控體內膽固醇和脂質代謝平衡的重要轉錄因子。LXR主要通過幾方面的協同作用調控膽固醇穩態:①腸道飲食中膽固醇的吸收;②肝膽固醇的生物合成和攝取;③高密度脂蛋白(high density lipoprotein,HDL)顆粒的合成和重塑及隨后的逆向膽固醇轉運;④肝中將膽固醇轉化為膽汁酸并排泄到膽汁中[11]。此外,LXR與脂肪酸生物合成及外周血中甘油三酯水平相關。
LXR的2種亞型在體內甾醇穩態中發揮獨特作用。在低密度脂蛋白受體(low-density lipoprotein receptor,LDLR)-/-小鼠背景下,LXRα-/-和LXR β-/-單基因或雙基因敲除實驗表明,LXRα和LXRβ均有抗動脈粥樣硬化作用,但LXRβ貢獻弱于LXRα,且2種亞型的功能有所區別,即LXRα在激動劑作用下啟動轉錄,而LXRβ可能在配體存在時抑制轉錄[12]。
2.1 調控膽固醇逆轉運
LXR可刺激膽固醇從外周細胞排出到肝,然后通過膽汁酸分泌,最終通過糞便排出體外,這一過程被稱為RCT。LXR被激活后,可促進肝細胞及巨噬細胞的膽固醇外流,并通過細胞色素P450 7A1(cytochrome P450 7A1,CYP7A1)將游離膽固醇轉化為膽汁酸,或經腸膽固醇排泄清除含載脂蛋白B(apolipoprotein B,ApoB)的脂蛋白膽固醇。此外,LXR激活并誘導脂肪生成基因固醇調節元件結合蛋白1c(sterol regulatory element-binding protein 1c,SREBP1c)、脂肪酸合成酶(fatty acid synthase,FAS)、硬脂酰輔酶A去飽和酶1(stearoyl-coenzyme A desaturase 1,SCD1)和LDLR誘導型降解子(inducible degrader of the LDLR,IDOL)的表達(圖3)[13-14]。其中,依賴ATP結合盒轉運蛋白 A1(ATP-binding cassette transporter A1,ABCA1)的膽固醇通過HDL從巨噬細胞外排的過程,在人心血管疾病的生理和病理進程中發揮重要作用[15]。下面將簡要介紹LXR靶基因的生理功能。
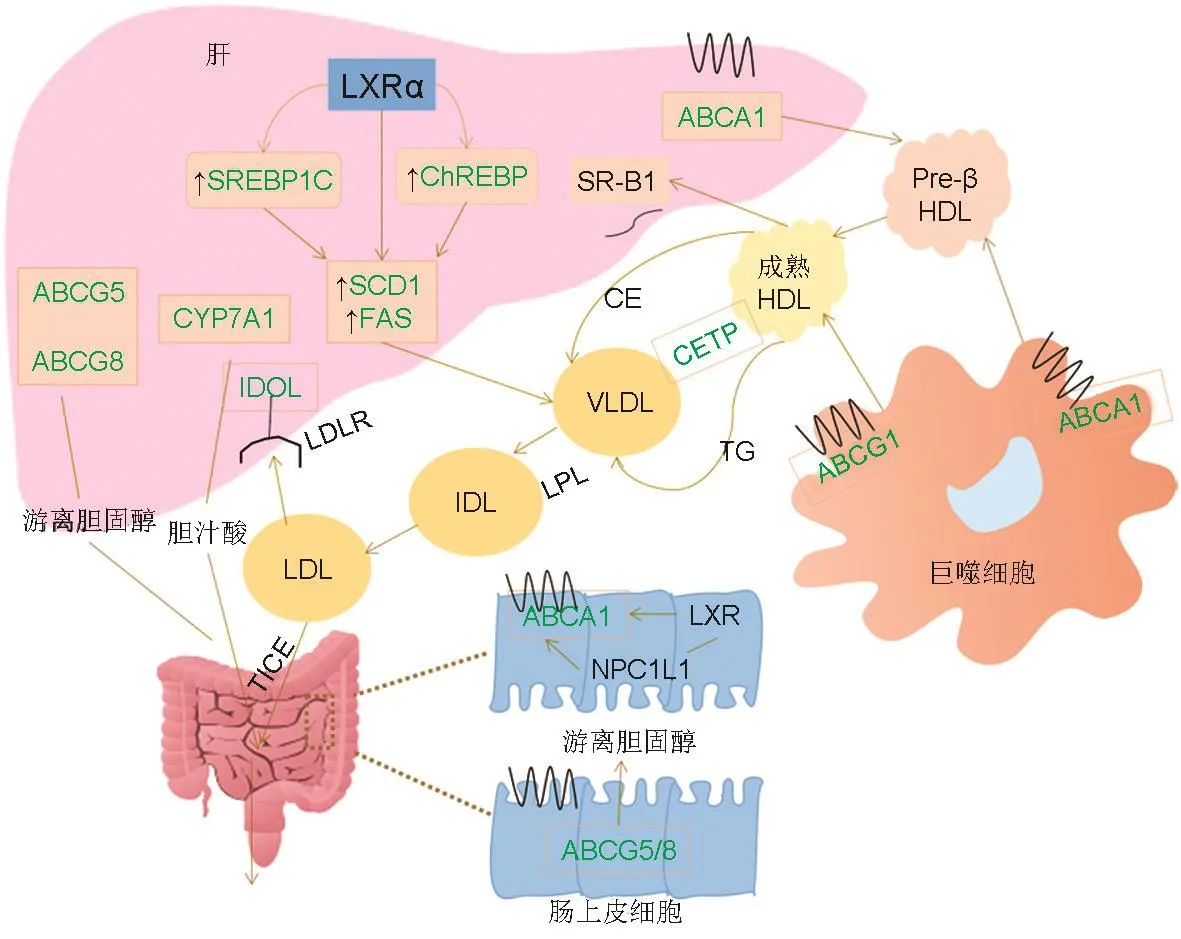
圖3 肝X受體調控膽固醇逆轉運.方框內所示為LXR靶基因,可被LXR激動劑調控.ABCA1:ATP結合盒轉運蛋白A1;SR-B1:清道夫受體B1;LXRα:肝X受體α亞型;SREBP1c:固醇調節元件結合蛋白1c;ChREBP:糖反應元件結合蛋白;SCD1:硬脂酰輔酶A去飽和酶1;FAS:脂肪酸合酶;CYP7A1:細胞色素P450 7A1(也稱為膽固醇-7α-羥化酶);TICE:經腸膽固醇排泄;IDOL:LDLR誘導型降解子;LDLR:低密度脂蛋白受體;LDL:低密度脂蛋白;HDL:高密度脂蛋白;Pre-β HDL:HDL前體;VLDL:極低密度脂蛋白;CE:膽固醇酯;CETP:膽固醇酯轉運蛋白;TG:甘油三酯;NPC1L1:類尼曼-匹克C1型蛋白;LPL:脂蛋白酯酶.
2.1.1 三磷酸腺苷結合盒轉運體家族
目前,已知的膽固醇外流途徑有:①游離膽固醇分子通過水相擴散結合到HDL顆粒;②清道夫受體B1(scavenger receptor class B type 1,SR-B1)介導途徑;③ABCA1介導磷脂和膽固醇從細胞向低脂Apo如ApoA1的主動外排途徑。而ABCA1參與的膽固醇外流是巨噬細胞RCT的主要途徑[16],這一過程促使細胞磷脂分子從細胞質小葉轉移到細胞質膜的外表面小體并促進ApoA1結合到細胞表面[17]。ABCA1廣泛參與腸、肝和巨噬細胞中的膽固醇和磷脂轉運,其功能缺失會導致嚴重后果。如在丹吉爾病(Tangier disease)中,ABCA1突變會阻斷膽固醇和磷脂在腸上皮細胞和肝細胞基底外側膜的轉運,致使因缺乏足夠的膽固醇和磷脂而影響ApoA1蛋白脂化。這些空載的ApoA1則被腎迅速清除出循環系統,進而導致嚴重的HDL缺乏綜合征[18]。
ABCG1也參與調控細胞內膽固醇穩態。生理條件下,ABCG1的受體包括較大HDL2顆粒(極大HDL和大HDL)與較小HDL3顆粒(中型HDL、小型HDL和極小型HDL)[19]。研究發現一種獨特現象,即ABCA1和ABCG1在刺激膽固醇外排和抑制動脈粥樣硬化方面發揮著協同作用[20]。一方面,ABCA1介導磷脂和膽固醇向低脂或無脂ApoA1的轉移,以生成新生的HDL顆粒;另一方面,上調ABCG1表達可促進膽固醇向HDL的流出。
而ABCG5和ABCG8也是LXR的直接靶基因,通常形成異二聚體ABCG5/ABCG8發揮作用。它們在內質網二聚化后,輸運至腸細胞的頂膜和肝細胞的小管膜,將細胞內固醇泵入腸腔和膽汁中,并最終通過糞便排出體外[21]。
2.1.2 載脂蛋白E
ApoE作為脂蛋白的組成部分,在維持血漿脂蛋白穩態中起著關鍵作用。ApoE主要由肝合成,其他器官和細胞,如腦、腎、脂肪細胞、平滑肌細胞和巨噬細胞也能產生ApoE[22]。ApoE主要通過ABCA1進行脂質化,而二者均受LXR的轉錄調控[23]。
2.1.3 膽固醇酯轉移蛋白
膽固醇轉移到HDL顆粒后,經卵磷脂膽固醇酰基轉移酶酯化形成膽固醇酯,最終形成成熟的HDL。肝脂肪酶和內皮脂肪酶分別介導甘油三酯和磷脂水解進而重塑HDL顆粒[24]。在表達膽固醇酯轉運蛋白的物種中,大部分HDL膽固醇酯可通過膽固醇酯轉運蛋白轉移至含ApoB的脂蛋白中,這些脂蛋白由肝細胞表面LDLR攝取從而促進RCT在體內運行[25]。
2.1.4 脂質轉運蛋白
磷脂轉運蛋白參與低脂HDL前體(pre-β HDL)的生成,為細胞膽固醇提供更多的受體,并參與維持循環系統中HDL的含量穩定[26]。脂蛋白脂酶是脂蛋白代謝的關鍵酶,專司循環脂蛋白中甘油三酯的水解,向外周組織釋放游離脂肪酸。在肝組織中,脂蛋白脂酶能促進HDL膽固醇的攝取,從而促進膽固醇的逆向轉運[27]。
2.1.5 CYP7A1
CYP7A1是膽固醇到膽汁酸的經典轉化途徑中的限速酶[28]。CYP7A1屬于細胞色素P450蛋白家族。與人不同,嚙齒類動物在CYP7A1基因啟動子中含有LXRE序列。此外,LXRα(而非LXRβ)是小鼠肝CYP7A1 mRNA表達的主要調節因子[29]。除膽汁酸的合成代謝外,LXR也參與調節膽汁酸的分解代謝。有研究表明,配體激活的LXRα通過與啟動子中的LXRE序列結合而上調人UDP葡萄糖醛酸轉移酶1家族多肽A3(UDP glucuronosyltransferase 1 family polypeptide A3)(葡糖醛酸結合膽汁酸最活躍的酶之一)基因的表達,從而使膽汁酸葡萄糖醛酸化,并最終轉化為可通過尿液排泄的代謝產物[30]。
2.2 調控膽固醇合成、分解和代謝
膽固醇的積累和清除分別由2種轉錄因子調節。在低膽固醇條件下,SREBP2促進膽固醇的合成和攝取,而細胞膽固醇水平過高時,LXR則促進膽固醇外流。研究表明,SREBP2作為ABCA1基因表達的正調節因子,通過誘導下游基因生成LXR的氧甾醇配體發揮作用[31]。而LXR則通過肝表達LXR誘導序列(liver-expressed LXR-induced sequence,LeXis)(一種非編碼 RNA鏈)[32]和內質網膜泛素連接酶環指蛋白145(ring finger protein 145,RNF145)[33]抑制膽固醇的生物合成。其主要機制是LeXis與異質核核糖核蛋白(RALY heterogeneous nuclear ribonucleoprotein)相互作用并影響后者與基因組DNA的相互作用[32]。RNF145誘導SREBP蛋白裂解激活蛋白將靠近外被體蛋白復合物Ⅱ(coat protein complex Ⅱ)結合位點的2個賴氨酸殘基泛素化,進而抑制其向高爾基體的轉運和SREBP2的后續加工[33]。
LXR還調節細胞內膽固醇的積累。研究發現,LXR通過調控IDOL抑制LDLR通路。已知IDOL是一種E3泛素連接酶,可觸發LDLR在細胞質內泛素化而被靶向降解[34]。LXR-IDOL途徑為SREBP2途徑提供了補充,即SREBP2途徑在低膽固醇條件下增加LDLR轉錄,而增加LDL膽固醇的攝取[35]。
類尼曼-匹克C1型蛋白1(Niemann-Pick C1-like1,NPC1L1)分布于腸上皮細胞刷狀緣膜,其功能包括介導腸細胞攝取游離膽固醇、將新分泌的膽汁膽固醇轉運回肝細胞,從而防止內源性膽固醇的過度流失。LXR可調控NPC1L1在小鼠和人腸細胞中的表達,降低腸道膽固醇的吸收[36]。
2.3 調控脂肪酸代謝
LXR激活后,通過誘導SREBP1c、糖反應元件結合蛋白(葡萄糖敏感轉錄因子,可將肝中過剩的糖類轉化為脂質)及SREBP1c靶蛋白FAS和SCD1的表達,促進脂肪酸的生物合成[13]。在脂肪組織中,LXR調節脂結合蛋白和代謝蛋白如ApoD和甲狀腺激素應答基因的表達,并可能通過誘導脂肪酸在線粒體中的β-氧化而促進其分解。此外,LXR通過誘導葡萄糖轉運蛋白4促進葡萄糖攝取。綜上,LXR影響脂肪組織中碳水化合物和脂質代謝,促進葡萄糖攝取,增強脂肪酸β-氧化及其分解[37]。
3 調控炎癥反應
除調節脂質穩態外,LXR同時參與調控多個炎癥信號通路。例如,LXR對白細胞介素18(interleukin-18,IL-18)和IL-1β的調節具有多重作用。LXR抑制脂多糖誘導的骨髓源性巨噬細胞IL-1β和IL-18表達水平,同時負向調節IL-18前體胱天蛋白酶1的水平,從而抑制IL-18的成熟,且LXR激活后通過調節干擾素調節因子8增強IL-18BP(IL-18的有效內源性抑制劑)的表達[38]。配體激活的LXR可以抑制典型炎癥基因的表達(如誘導型一氧化氮合酶、環氧合酶2和基質金屬蛋白酶9)和各種趨化因子對脂多糖、腫瘤壞死因子α和IL-1β刺激的反應[39]。
有報道稱,LXR通過以小泛素相關樣修飾物(small ubiquitin-like modifier,SUMO)信號通路依賴的方式與NF-κB轉錄復合物相互作用來抑制炎性基因表達[40]。而Ito等[41]報道,在無SUMO的條件下,LXR激活仍發揮抗炎作用,其機制主要是通過調控固醇轉運蛋白ABCA1的轉錄,改變膜膽固醇穩態,抑制Toll樣受體下游的NF-κB和絲裂原活化蛋白激酶介導的炎癥信號。
Kappus等[42]發現,在缺乏ABCA1 和ABCG1的條件下,激活LXR仍產生有效的抗動脈粥樣硬化作用,即LXR發揮抗炎作用不依賴于巨噬細胞ABCA1和ABCG1介導的膽固醇外流通路。而Westerterp等[43]研究表明,ABCA1和ABCG1缺陷導致骨髓細胞中的膽固醇積累并激活NOD樣受體蛋白3炎性小體,促進動脈粥樣硬化病變中的中性粒細胞浸潤和網狀病變。另有研究發現,LXR激動劑通過順式抑制(cis-repression)LXR和調節膽固醇代謝可抑制體內無菌性炎癥時中性粒細胞的遷移[44]。綜上,LXR通過多種途徑調節炎癥信號通路,其具體機制需要深入研究。
4 肝X受體配體
動脈粥樣硬化的發展可導致心臟病發作和卒中等嚴重后果。LXR激活可發揮幾種不同的抗動脈粥樣硬化作用,如增加外周膽固醇的流出、促進膽固醇向肝的逆向轉運及膽汁酸分泌、抑制炎癥、提高巨噬細胞的存活率并減少其對含ApoB脂蛋白的攝取[45],充分表明LXR是治療動脈粥樣硬化的潛在藥物靶標。綜上,通過配體激活LXR控制膽固醇通量從而預防和治療心血管疾病具有很好的應用前景。
LXR的整體結構尤其是配體結合口袋(ligandbinding pocket)可非常靈活地與結構高度不同的配體結合。激動劑在配體結合口袋中的結合導致LXR-RXR異二聚體的受體構象發生變化,從而影響異二聚體的轉錄活性(圖4)[46]。
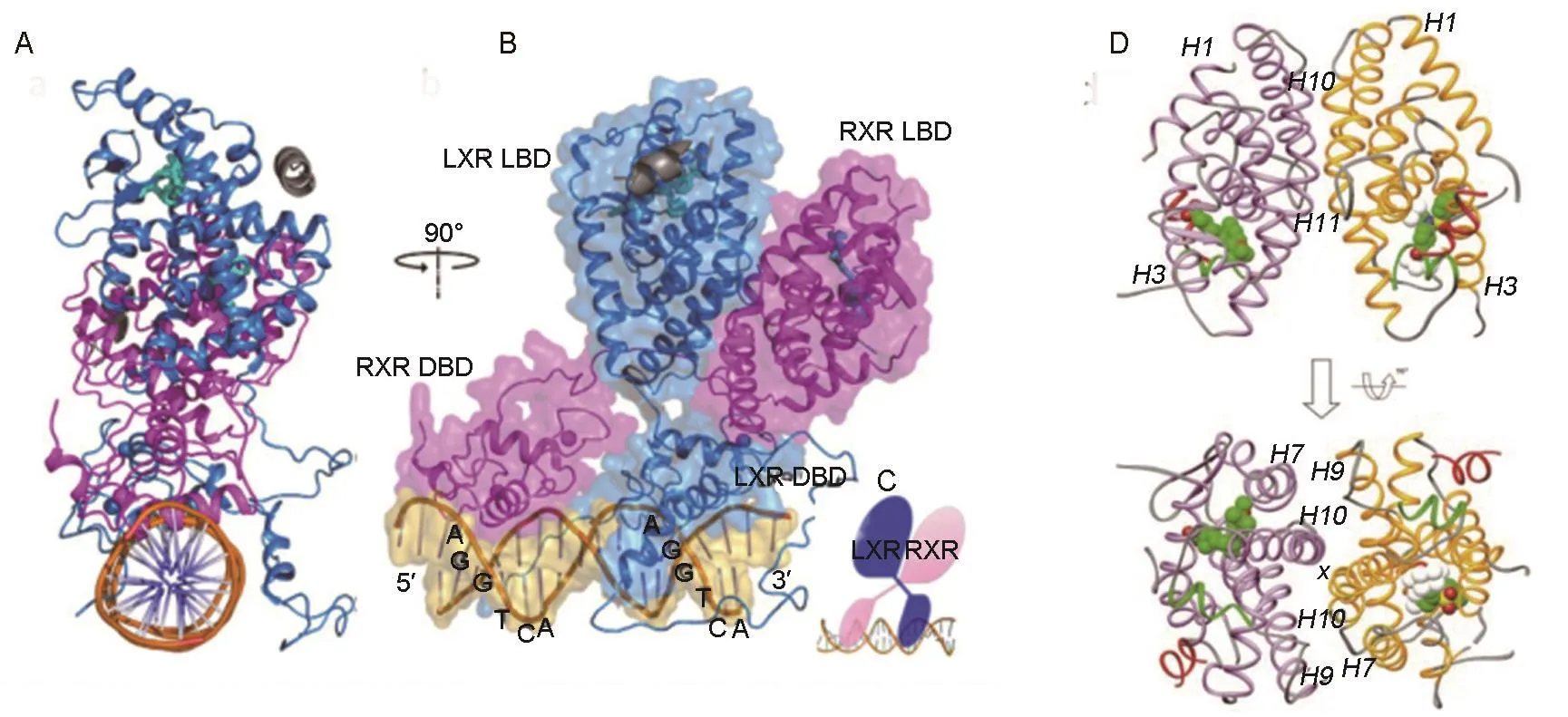
圖4 肝X受體/類視黃醇X受體異二聚體整體結構示意圖.A和B:RXRα/LXRβ沿DNA方向(5′到3′)結構和側面結構(藍色:LXRβ;品紅色:RXRα;青色:配體;灰色:輔因子肽);C:RXRα/LXRβ異二聚體中結構域位置;D:LXRα/RXRβ異二聚體LBD的結構(黃色:LXRα-LBD;紫色:RXRβ-LBD;綠色:LXR羧端配體依賴的轉錄活化域螺旋).
4.1 內源性配體
4.1.1 內源性激動劑
最初研究認為,LXR是一種孤兒受體。但后續研究證實,氧化甾醇是LXR的內源性配體。氧化甾醇是膽固醇的氧化衍生物,包括24(S)-和25-環氧膽固醇及22(R)-、24(S)-、20(S)-和 27-羥基膽固醇(但不包括膽固醇)均是LXR的配體。22(R)-和20(S)-羥基膽固醇是甾體激素合成過程中的中間產物。24(S)-羥基膽固醇在大腦中產生,是循環系統中最豐富的氧化甾醇,而24(S)-和25-環氧膽固醇主要存在于肝中[47]。
4.1.2 內源性拮抗劑
花生四烯酸和其他不飽和脂肪酸競爭性地阻斷LXR的激活,抑制T0901317誘導的內源性SREBP1c基因表達升高。前列腺素F2α(花生四烯酸的環氧合酶代謝產物之一)以劑量依賴性方式拮抗T0901317誘導的LXRα-LBD和LXRβ-LBD激活,并拮抗ABCA1和ABCG1啟動子的激活[48]。
4.2 天然產物配體
LXR的配體包括很多天然產物或其衍生物(表1)。研究發現,花青素可與LXRα和LXRβ結合,調控靶基因如ABCA1和ABCG5的表達,但對LXRα的親和力高于 LXRβ(LXRα EC50為 3.5 μmol·L-1,LXRβ為125.2 μmol·L-1)。單萜桉樹腦是在茶和草藥精油中發現的成分,可組織特異性激活LXRα和LXRβ。桉樹腦(50~200 μmol·L-1)處理RAW264.7巨噬細胞后,ABCA1和ABCG1 mRNA水平顯著升高;桉樹腦未顯著影響HepG2肝細胞中SREBP1c mRNA表達,而FAS和SCD1 mRNA表達顯著下降。芍藥苷是一種單萜苷,用于治療高脂血癥和高血糖。熒光素酶報告基因檢測發現,芍藥苷10 μmol·L-1可顯著激活HepG2細胞LXRα。槲皮素或槲皮素糖苷被胃腸道吸收后,形成槲皮素-3-O-葡萄糖苷。槲皮素-3-O-葡萄糖苷(50 μmol·L-1)可增加RAW264.7巨噬細胞中ABCA1 mRNA和蛋白水平[49]。
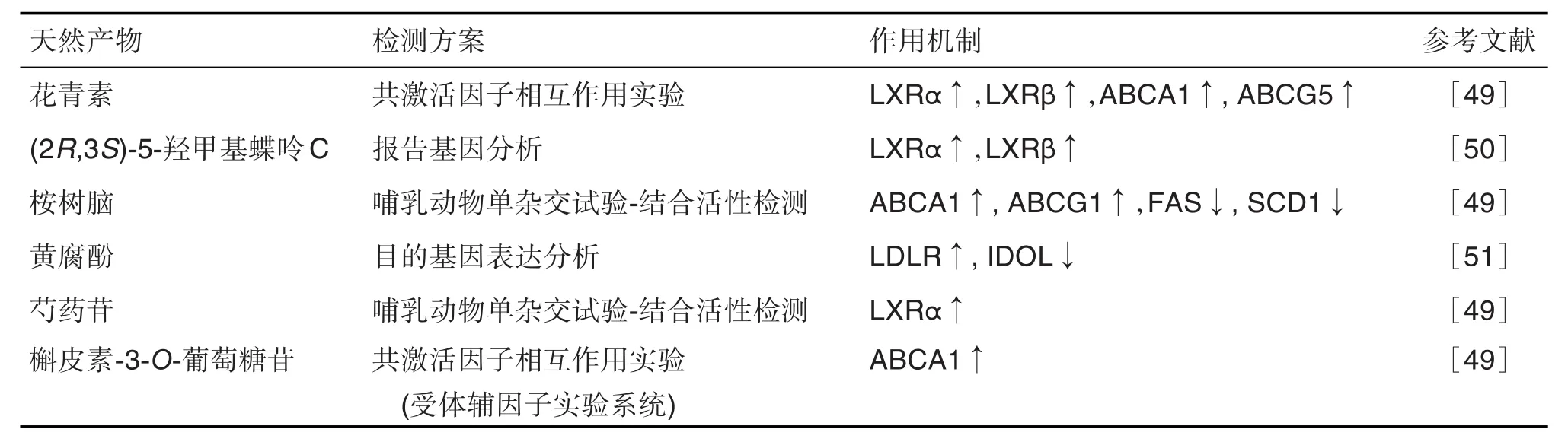
表1 肝X受體的天然產物配體
從植物氣生部分分離出的化合物,如蝶呤倍半萜(2R,3S)-5-羥甲基蝶呤C,亦可激活LXRα和LXRβ,比 GW3965(10 μmol·L-1)對 2 種 LXR 亞型的激活更明顯[50]。Chen等[51]研究發現,黃腐酚可增加細胞表面LDLR表達,進而增加HepG2細胞中LDL攝取;黃腐酚(10和20 μmol·L-1)可顯著降低IDOL mRNA和蛋白的表達。此外,與T0901317單一處理相比,用黃腐酚 20 μmol·L-1預處理 HepG2細胞后與T0901317共孵育,IDOL mRNA和蛋白質水平均降低,LXRα mRNA和蛋白水平無明顯改變,表明黃腐酚可抵消LXR的激活。
4.3 合成配體
經典LXR合成類激動劑有T0901317和GW3965等[52]。在非臨床動物模型中,合成的LXR激動劑可改善動脈粥樣硬化和廣泛的炎癥性疾病,但系統性LXR激活可引發肝脂肪變性和高甘油三酯血癥(LXRα在肝中誘導脂肪生成基因表達的后果)。近來研究出一些新的防治策略,包括LXRβ選擇性激動劑、組織選擇性LXR激動劑和轉抑制選擇性LXR激動劑等[14]。不同種類的LXR合成配體見表2。
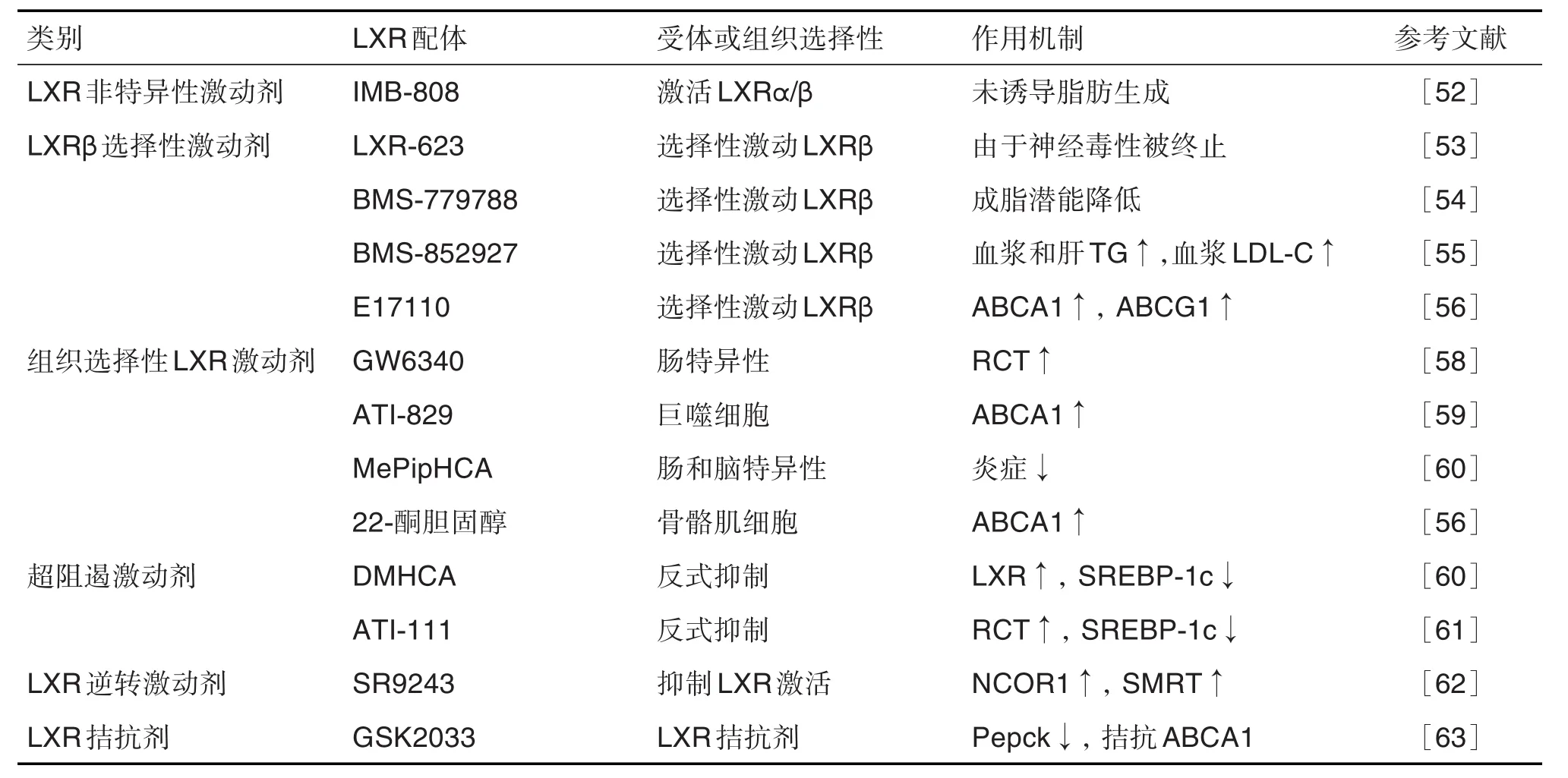
表2 肝X受體的合成配體(激動劑和拮抗劑)
4.3.1 非特異性激動劑
IMB-808激活LXRα和LXRβ,在多個細胞系中有效增加與RCT以及與膽固醇代謝途徑有關基因的表達,尤其顯著促進RAW264.7和THP-1巨噬細胞的膽固醇外流,并相應減少細胞脂質的積累,且不會上調脂肪生成基因的表達[52]。
4.3.2 肝X受體 β選擇性激動劑
LXRβ激動劑LXR623可增加外周血細胞中LXR靶基因ABCA1和ABCG1的表達,但最高劑量時可引起中樞神經系統和精神疾病等不良反應,導致其研發終止[53]。BMS-779788和BMS-852927在食蟹猴或小鼠中選擇性激活LXRβ,可顯著改善血脂異常,誘導RCT相關基因表達且具有較低的成脂潛能[54-55]。與其他LXR激動劑相同,LXRβ激動劑E17110在體外可增加ABCA1和ABCG1表達,并增強RAW264.7巨噬細胞中的膽固醇外流,表明其可能具有抗動脈粥樣硬化活性[56]。
4.3.3 組織選擇性激動劑
Li等[57]建立了62個甾醇衍生物庫并進一步設計、合成和測試了12個甾醇衍生物的生物學性質,證明化合物4和6是LXR激動劑,可以抑制巨噬細胞泡沫細胞的形成而不會誘導肝細胞或脂肪細胞中甘油三酯的累積。GW6340是腸特異性激動劑,顯著上調小腸中的ABCA1,ABCG5和ABCG8的表達,促進巨噬細胞RCT,但在肝中無此作用[58]。ATI-829可在THP-1巨噬細胞中強烈誘導ABCA1表達,且在HepG2肝癌細胞中未明顯上調SREBP1c表達。LDLR-/-小鼠給藥后,ATI-829激活了腸道和巨噬細胞中的LXR靶基因,但未激活肝中脂肪酸合成基因而未增加肝和量[59]。MePipHCA是一種基于甾醇的LXR激動劑,抑制葡聚糖硫酸鈉結腸炎和創傷性腦損傷2種模型小鼠炎癥而未引起肝脂質蓄積或肝損傷[60]。
4.3.4 其他
最近開發的LXR激動劑還有反向激動劑等類型[62]。Zhang 等[64]報道,含有合成 LXR 激動劑GW3965的納米顆粒〔(乳酸-共聚乙二醇)-b聚(乙二醇)共聚物〕具有抗炎作用,并在不引起肝脂肪變性的情況下抑制動脈粥樣硬化的發展。
5 結語
LXR在調控膽固醇代謝過程中起到核心作用。此外,LXR在調控糖代謝、脂質代謝、炎癥反應以及先天免疫等方面也發揮重要作用。隨著對LXR在動脈粥樣硬化疾病發展中的作用及合成LXR配體的深入研究,以LXR為靶點尋找激動劑,在治療高脂血癥和動脈粥樣硬化等疾病方面將具有很好的應用前景。LXR協調抑制炎癥的能力對于理解脂質代謝和炎癥之間的相互作用以及藥物發現具有重要意義,但LXR的這種協調作用仍然存在很多疑問。目前常用的LXR合成激動劑,多會引起脂肪在肝的積累以及血中甘油三酯水平升高。LXR的功能研究為動脈粥樣硬化和眾多炎癥性疾病的治療提供了潛在的機會。然而,由于嚙齒類動物以及非人類靈長類動物與人之間的種屬差異,難以預測LXR激動劑對人體的不良反應,致使LXR激動劑的研究進一步復雜化且困難重重。因此,深入研究LXR的作用機制,充分了解LXR調控脂質代謝的組織特異性,將有助于開發更加安全有效的LXR選擇性激動劑,也將為開發新型的抗動脈粥樣硬化藥物及多種心血管疾病的治療提供理論基礎。此外,將LXR激動劑與其他可減輕脂肪生成的藥物(如IDOL拮抗劑)進行聯合用藥也是一種可行方案。隨著LXR激動劑作用機制研究的不斷深入,綜合結構信息指導下的藥物研發,最終會實現靶向藥物的有效和安全性應用。
