枳橙四倍體與二倍體干旱生理應答及水通道蛋白基因表達分析
2021-08-04 09:14:02李曉妍陳芝琳鐘八蓮
中國南方果樹 2021年4期
關鍵詞:差異
彭 瀅,李曉妍,陳芝琳,彭 婷,2,鐘八蓮,2,肖 璇,2
(1 贛南師范大學生命科學學院,江西贛州,341000;2 國家臍橙工程技術研究中心,江西贛州,341000)
干旱是影響柑桔產量最重要的非生物脅迫因素之一。我國柑桔大部分集中在南部和東南部地區,受季節性干旱脅迫的問題較突出[1]。柑桔作為嫁接繁殖為主的果樹,選擇抗逆性和適應性更強的砧木對生產有重要影響[2]。與二倍體相比,多倍體植株通常在形態、組織結構、生理代謝等方面發生明顯變化,具有更強的抗逆性[3-5]。近年來柑桔研究者通過實生選種方法已發掘大量多倍體砧木資源[6-7]。研究表明,枳四倍體成熟葉片可積累更多的檸檬酸、蘋果酸、延胡索酸等初生代謝產物[8],干旱脅迫下枳四倍體具有更高的活性氧清除能力并可積累更多的可溶性糖[9]。近年來,部分水通道蛋白(aquaporin,AQPs)已被證實參與植物對水分的運輸[10-11]。根據序列的同源性及亞細胞定位差異可將AQPs分為4大類[12]:(1)細胞質膜水孔蛋白(PIP),(2)根瘤菌和豆科植物的共生膜上的類Nod26 膜水孔蛋白(NIP),(3)液泡膜上小分子堿性膜水孔蛋白(SIP),(4)泡膜水孔蛋白(TIP)。目前,關于柑桔四倍體砧木資源耐旱性鑒定、生理應答及水通道蛋白基因表達方面研究仍較少。枳橙[Citrussinensis(L.) Osb.×Poncirustrifoliata(L.) Raf.]是甜橙與枳的雜種,具多胚性,為柑桔主要砧木之一。筆者以枳橙二倍體為對照,對干旱脅迫下枳橙四倍體植株葉片失水表型、生理響應以及水通道蛋白基因表達差異進行分析,對枳橙四倍體耐旱能力進行評價和初步解析。
1 材料與方法
1.1 試驗材料及處理
研究于2019年6月在國家臍橙工程研究中心進行。供試材料為2年生的枳橙二倍體和同源四倍體。枳橙同源四倍體為本課題組從枳橙實生苗篩選獲得,經SSR分子標記鑒定其來源可能為二倍體母本珠心組織基因組加倍[13]。采用盆栽控水自然干旱方式進行干旱脅迫處理。塑料圓盆直徑15 cm、高22 cm,基質為果園10 cm深處園土和營養土等體積混和而成。6月下旬盆栽定植于遮雨網室,自然光、溫條件,試驗前每天澆透水1次,連續3 d,最后一次澆透水后進行自然干旱脅迫處理。分別在最后一次澆透水后當天(0 d)及3 d、6 d、9 d、12 d、15 d及復水5 d后取樣(連續自然干旱15 d后開始復水,復水后每天均澆透水1次)。每個取樣日,于上午9點定時采摘當年新梢從上向下數第4—6節位葉片(干旱脅迫前均為完全舒展的健康葉片),進行相關生理生化指標測定及總RNA提取。葉片離體脫水試驗取正常澆水的健康葉片,具體參照Wei等[9]方法進行。每3株采摘葉片混為一組,重復測定3次。
1.2 測定方法
超氧化物歧化酶(Superoxide dismutase,SOD)活性、丙二醛(malondialdehyde,MDA)含量、脯氨酸(Proline,Pro)含量及相對電導率的測定參照李合生[14]的方法,過氧化物酶(Peroxidase,POD)活性采用愈創木酚法[15]。
RNA提取參照Trizol試劑盒(Life Technologies,Inc.,Carlsbad,CA,USA)方法進行。采用NanoDrop 2000超微量核酸蛋白分析儀(Thermo Scientific,USA)測定RNA濃度。采用TaKaRa的PrimeScriptTMRT reagent Kit將RNA反轉錄為cDNA,統一稀釋至150 ng/μL,作為熒光定量的模板。以β-actin基因為內參[16]。實時定量PCR所用水通道蛋白基因引物參照馮芳芳[11],由北京擎科生物技術有限公司合成,具體見表1。應用 SYBR Green 熒光染料法在Roche公司Light Cycler?480熒光定量PCR儀上進行擴增分析。采用2-△△ct法分析基因相對表達量[17]。

表1 針對各水通道蛋白基因進行Real-time qPCR 的引物序列
葉片離體脫水試驗中相對失水率計算公式如下:相對失水率(%)=[(W0-Wn)/W0]×100%,W0為葉片離體失水處理前葉片鮮質量,Wn為葉片離體失水處理n小時后的質量。
1.3 數據分析
試驗數據用Excel 2016進行整理,用SPSS 21.0進行顯著性差異分析。
2 結果與分析
2.1 干旱脅迫下的形態差異
干旱處理前,兩種倍性枳橙的葉片均呈綠色、自然舒展狀(圖1A);干旱8 d,二倍體植株失水明顯,整株葉片呈萎蔫狀,部分葉片失綠黃化;與之不同的是,四倍體植株僅呈輕度萎蔫狀,葉片黃化不明顯(圖1B);干旱15 d,二倍體植株的葉片失水嚴重,整株葉片黃化枯萎(圖1C),四倍體植株僅梢端嫩葉萎焉嚴重。從干旱脅迫后盆栽植株及葉片表型來看,四倍體枳橙比二倍體枳橙具更強的耐旱性。

注:A.干旱0 d,B.干旱8 d,C.干旱15 d;在圖A、B、C中,左為二倍體,右為四倍體。
2.2 離體葉片的失水率差異
葉片離體脫水試驗結果看出,隨著葉片離體脫水時間的增加,二倍體和四倍體枳橙葉片相對失水率均不斷上升,但同一時間四倍體和二倍體的相對失水率差異均不顯著(圖2)。

圖2 離體不同時間后枳橙二倍體和四倍體葉片的相對失水率
2.3 干旱脅迫下的葉片相對電導率差異
干旱處理前期,兩種倍性葉片的相對電導率相差不顯著;隨著干旱的進行,二倍體和四倍體葉片的相對電導率均呈上升趨勢,且從干旱9 d開始有顯著性差異(p<0.05),二倍體葉片的相對電導率始終高于四倍體;復水后5 d,兩者的相對電導率均迅速下降,且兩者差異不顯著(見圖3)。說明,在干旱脅迫下四倍體葉片的細胞質膜受損要小于二倍體。
2.4 干旱脅迫下的葉片MDA含量差異
隨著干旱脅迫的進行,二倍體和四倍體葉片的MDA含量均呈上升趨勢,說明兩種倍性植株葉片組織細胞膜受損程度不斷加重。從干旱脅迫12 d開始,二倍體葉片的MDA含量顯著高于四倍體(p<0.05)。復水后5 d,兩種倍性葉片的MDA含量都迅速下降,但二倍體仍顯著高于四倍體(見圖4)。

注:“*”表示差異達顯著水平(p<0.05);“**”表示差異達極顯著水平(p<0.01)。圖4至圖7同。
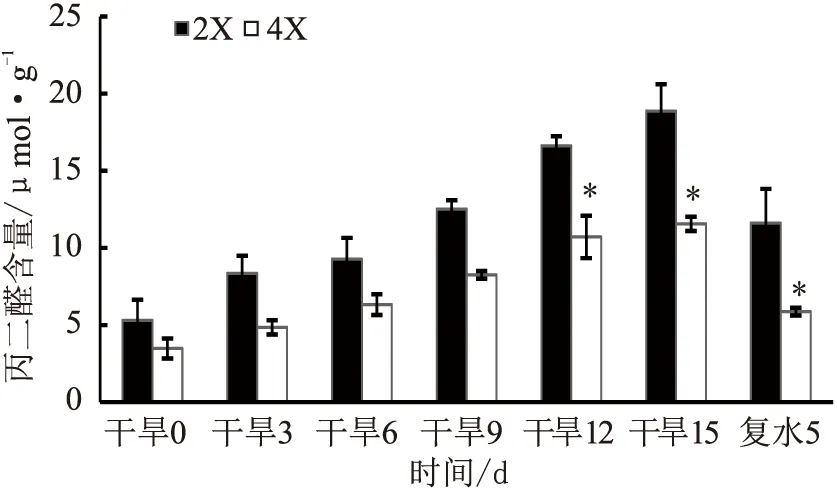
圖4 干旱脅迫對兩種倍性枳橙葉片MDA含量的影響
2.5 干旱脅迫下的葉片活性氧去除相關酶活性差異
隨著干旱脅迫的進行,兩種倍性枳橙葉片SOD活性表現為先升后降;二倍體在干旱6 d達到最大值,四倍體在干旱9 d達到最大值;自干旱脅迫9 d開始,四倍體的SOD活性始終高于二倍體(見圖5)。隨著干旱脅迫的進行,兩種倍性枳橙葉片POD活性總體也呈先升后降趨勢;在干旱6 d均達到最大值,但此時四倍體的POD活性顯著高于二倍體,其他時期均無顯著差異(見圖6)。復水后5 d,兩種酶活均接近于正常狀態,四倍體恢復速率較二倍體快(見圖5和圖6)。

圖5 干旱脅迫對兩種倍性枳橙葉片SOD活性的影響
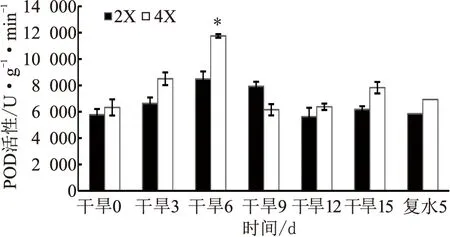
圖6 干旱脅迫對兩種倍性枳橙葉片POD活性的影響
2.6 干旱脅迫下的葉片Pro含量差異
隨著干旱脅迫的進行,兩種倍性的葉片Pro含量均有所上升;復水后5 d,兩者Pro含量均有所下降。在干旱9 d時四倍體的Pro含量高于二倍體(p<0.05),其他時期兩者無顯著差異(見圖7)。

圖7 干旱脅迫對兩種倍性枳橙葉片Pro含量的影響
2.7 干旱脅迫下的葉片水通道蛋白基因表達差異
植物水分跨膜運輸主要是由水通道蛋白(AQPS)所主導。從干旱脅迫下二倍體和四倍體葉片中9個水通道蛋白基因表達模式來看,干旱脅迫會引起水通道蛋白基因表達的變化,且PIP、NIP、TIP、SIP4種類型基因在兩種倍性葉片中表達量的變化情況不一致。在干旱脅迫條件下,二倍體葉片中PtrPIP1;2、PtrPIP1;3、PtrPIP2;2、PtrNIP1;2、PtrTIP1;3、PtrTIP4;1、PtrSIP1;1、PtrSIP1;2表達量總體呈下調趨勢,其中PtrTIP4;1表達量持續下調,其他都是先上調再下調;PtrNIP1;1表達量則呈總體上調趨勢。在干旱脅迫條件下,四倍體葉片中PtrNIP1;1、PtrNIP1;2、PtrTIP4;1、PtrSIP1;2表達量總體下調;PtrPIP1;2、PtrPIP1;3、PtrPIP2;2、PtrTIP1;3、PtrSIP1;1表達量上調,除了PtrSIP1;1外,均在干旱15 d達到最大值。在干旱脅迫15 d時,9個水通道蛋白基因中,只有PtrNIP1;1的表達量是二倍體高于四倍體,其余8個基因均是二倍體中的表達量低于四倍體(見圖8)。
3 討論
干旱脅迫下,植物在表型上會出現一系列的失水癥狀。本研究中,干旱脅迫下二倍體枳橙葉片萎蔫、黃化比四倍體枳橙更為嚴重,表明四倍體枳橙的耐旱性更強。兩者在耐旱性上的差異,可能與其在干旱應答生理及水分運輸相關的水通道蛋白基因表達差異有關。
3.1 干旱脅迫下生理響應差異對耐旱性的影響
四倍體植物在形態、生理代謝等方面的變化通常表現出比二倍體更加耐旱的特性[18-19]。相對失水率可反映植物葉片的持水能力,相對失水率越小,其持水能力則越高,耐旱性越強。相對電導率則反映植物組織細胞質膜在逆境脅迫中受損的程度,從而作為植物抗逆性的一個生理指標。MDA是膜脂過氧化的主要產物之一,MDA含量越高同樣表明細胞膜受傷害程度越大[20]。本研究表明,在干旱處理過程中,四倍體枳橙葉片的相對失水率、相對電導率及MDA含量均低于二倍體,說明四倍體的質膜受損程度較二倍體要輕,四倍體的抗旱性更強。這與鄧敏捷等[21]在泡桐四倍體抗旱性研究中的結果類似。

注:不同小寫字母表示相同材料不同干旱時間處理之間的相對表達量差異達顯著水平(p<0.05)。
干旱脅迫下,植物體內活性氧含量會急速增加,可導致膜脂過氧化甚至膜蛋白的降解[22]。超氧化物歧化酶(SOD)和過氧化物酶(POD)是植物體內清除活性氧重要的保護酶之一,能清除活性氧自由基,減輕活性氧對植株造成的傷害,維持細胞膜的穩定[23]。脯氨酸(Pro)含量的增加可有效調節細胞質內滲透過程,防止細胞脫水,是重要的抗旱生理指標[24]。本研究表明,在干旱脅迫過程中,二倍體枳橙和四倍體枳橙葉片的SOD、POD含量表現為先升后降趨勢,但干旱脅迫后四倍體的SOD及POD活性基本都高于二倍體,說明干旱脅迫下四倍體的活性氧清除能力比二倍體更強;干旱脅迫后期SOD、POD酶活性有所下降,其原因可能與干旱脅迫導致細胞受損有關[25]。本研究中,干旱處理前6 d,兩種倍性枳橙葉片Pro含量均緩慢增加且兩者含量差異不顯著;在干旱9 d時Pro含量迅速增加,四倍體Pro顯著高于二倍體,表明干旱脅迫下四倍體滲透調節能力更強。這些結果與尚小紅等[26]研究四倍體木薯耐旱的生理響應類似,進一步表明四倍體枳橙對干旱脅迫有更強的調節適應能力。
3.2 干旱脅迫下水通道蛋白基因表達對耐旱性的影響
植物水通道蛋白基因(AQPs)主要參與植物體內的水分運輸、滲透調節以及對養分的吸收[27]。受干旱環境的影響,AQPs可增強細胞的水分跨膜運輸能力[28]。本研究對枳橙葉片部分AQP基因進行了分析,結果顯示干旱脅迫中大部分基因在四倍體中是顯著上調且表達量高于二倍體,這一結果與薛浩等[29]類似。植物水通道蛋白與生理特征是密切相關的[30],在逆境脅迫下兩者共同調控和發揮作用,應對環境變化。
在干旱脅迫下,二倍體和四倍體兩種倍性枳橙生理應答及水通道蛋白基因表達分析結果,與兩種倍性枳橙葉片離體脫水及盆栽植株自然干旱鑒定的結果基本一致。植物的抗旱性是一個復雜的過程,本研究從干旱生理應答與水通道蛋白基因表達角度對枳橙二倍體和四倍體抗旱能力差異進行比較分析,進一步加深了對枳橙四倍體抗旱機制的認識。關于植物水通道蛋白基因在枳橙干旱脅迫中的具體功能及作用機制還有待進一步的深入解析。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
