2G型肢帶肌營養(yǎng)不良癥斑馬魚疾病模型的構(gòu)建
2021-09-22 02:24:20李麗萍鐘靜牛玉娟孫源超吳傳鴻李美航周建峰丁永和
精準(zhǔn)醫(yī)學(xué)雜志 2021年4期
關(guān)鍵詞:基因突變
李麗萍 鐘靜 牛玉娟 孫源超 吳傳鴻 李美航 周建峰 丁永和
(青島大學(xué),山東 青島 266071 1 基礎(chǔ)醫(yī)學(xué)院生物化學(xué)與分子生物學(xué)系; 2 生物醫(yī)學(xué)研究院)
2型肢帶肌營養(yǎng)不良癥(LGMD2)是一類具有肢帶肌無力、肌肉萎縮臨床表現(xiàn)的常染色體隱性遺傳的家族性疾病[1],根據(jù)致病基因的不同可分為不同的亞型,其中LGMD2G是LGMD2的亞型之一,是由定位于17q12區(qū)域的編碼Telethonin蛋白的TCAP基因突變導(dǎo)致的[2],在中國、印度和歐洲等國家人群中均有發(fā)現(xiàn)[2,3-7]。LGMD2G的主要臨床特征是遠(yuǎn)端肢體肌無力,患者在20歲左右開始出現(xiàn)下肢無力癥狀,約40%的患者在40歲左右失去了獨立行走的能力[8],生活質(zhì)量受到嚴(yán)重影響。鑒于LGMD2G發(fā)病機制尚不明確,目前臨床上尚沒有針對LGMD2G的有效治療手段。
由于人類遺傳學(xué)研究的限制,LGMD2G疾病相關(guān)分子機制的研究主要依賴于動物模型。斑馬魚用于科學(xué)研究已有80多年的歷史[9],因具有體積小、繁殖能力強、易于基因改造的優(yōu)點,目前廣泛應(yīng)用于藥物的篩選[10],在動物疾病模型的構(gòu)建中也顯示出了其特有的優(yōu)勢,然而目前LGMD2G斑馬魚疾病模型的構(gòu)建及致病機制的研究還處于初步探索階段。本研究利用CRISPR/Cas9基因編輯技術(shù)致斑馬魚tcap基因突變來構(gòu)建LGMD2G的活體動物模型,并進(jìn)一步利用該模型初步探討tcap基因在LGMD2G中的功能以及潛在作用機制,旨在為進(jìn)一步研究LGMD2G的致病機制以及進(jìn)行治療藥物的篩選奠定基礎(chǔ)。
1 材料與方法
1.1 動物和材料
Tübingen 品系斑馬魚由中國海洋大學(xué)分子醫(yī)學(xué)生物學(xué)實驗室提供,人工養(yǎng)殖于自動水循環(huán)系統(tǒng)中。T7核酸內(nèi)切酶、胚胎牛血清(FBS)、T7體外轉(zhuǎn)錄試劑盒、DNase Ⅰ(美國Thermo 公司),異丙醇、20×SSC 緩沖液、氯仿-異戊醇等(上海生工生物工程股份有限公司),KOD(plus)和KOD F(x)DNA聚合酶(日本TOYOBO 公司),反轉(zhuǎn)錄試劑盒、實時熒光定量PCR(RT-qPCR)試劑盒(加拿大ABMgood公司),酚紅、多聚甲醛(美國Sigma 公司),蘇木精-伊紅(HE)染色試劑盒(北京索萊寶科技有限公司),地高辛(DIG) 抗體、DIG 標(biāo)記的 NTP(瑞士Roche 公司)。
1.2 實驗方法
1.2.1小向?qū)NA(sgRNA)合成和顯微注射 依據(jù)從美國國家生物技術(shù)信息中心(NCBI)網(wǎng)站下載的斑馬魚tcap基因序列設(shè)計CRISPR/Cas9靶點,靶點的序列為5′-ATTGATGCTGCGCCAGTCAG-
3′。根據(jù)靶點序列設(shè)計sgRNA合成所需引物,以pT7-gRNA質(zhì)粒為模板進(jìn)行PCR擴增,用酚-氯仿抽提法抽提PCR產(chǎn)物,利用T7體外轉(zhuǎn)錄試劑盒合成sgRNA,用體積分?jǐn)?shù)為0.01的瓊脂糖膠進(jìn)行核酸電泳,檢測sgRNA是否制備成功。sgRNA制備成功后用酚紅將sgRNA稀釋為400 mg/L,將Cas9蛋白與400 mg/L的sgRNA各取1 μL混勻,將sgRNA稀釋為終濃度200 mg/L,將1 nL上述稀釋液于顯微鏡下注射至野生型(WT)斑馬魚胚胎卵黃(發(fā)育時期為1細(xì)胞期)中,然后胚胎置于培養(yǎng)液中于28.5 ℃恒溫培養(yǎng)箱中培養(yǎng)。
1.2.2tcap基因純合突變斑馬魚篩選 將于顯微鏡下注射后的斑馬魚胚胎培養(yǎng)2~3 d,然后進(jìn)行基因組提取。通過Prime 5.0軟件設(shè)計篩選tcap基因突變斑馬魚的引物序列如下:F-5′-GTTTGGATA-TGTGGGTG-3′,R-5′-AAATGTAGCCTACTGG-TG-3′;PCR擴增以后進(jìn)行T7核酸內(nèi)切酶酶切,選擇能切開的PCR產(chǎn)物進(jìn)行DNA測序,篩選獲得F0代tcap基因突變斑馬魚。成年后將其與WT斑馬魚交配獲得F1代tcap基因突變斑馬魚。對成年后的F1代tcap基因突變斑馬魚進(jìn)行篩選并確定突變類型,將相同突變類型的F1代tcap基因突變雌雄斑馬魚交配后,再次篩選獲得F2代tcap基因突變斑馬魚,即為tcap基因純合突變斑馬魚。然后通過GENEDOC軟件分析tcap基因純合突變斑馬魚的tcap基因所編碼的telethonin蛋白氨基酸序列的變化情況。
1.2.3斑馬魚骨骼肌組織石蠟切片以及HE染色成年WT和tcap基因純合突變斑馬魚安樂死后切取骨骼肌組織,置于體積分?jǐn)?shù)為0.04的多聚甲醛溶液中固定,經(jīng)水洗、梯度脫水、透明、透蠟后進(jìn)行石蠟包埋和切片;將切片后的組織進(jìn)行脫蠟和梯度復(fù)水,以蒸餾水沖洗組織切片5 min后用蘇木精染料染色3 min;然后經(jīng)含體積分?jǐn)?shù)0.01的鹽酸乙醇溶液沖洗后再以蒸餾水沖洗12 min,伊紅染料染色30 s;經(jīng)乙醇和二甲苯處理后封片,置于顯微鏡下觀察并拍照。
1.2.4斑馬魚最大游泳速度檢測 選取體長(BL)和體質(zhì)量相近的成年WT斑馬魚(A組)和tcap基因純合突變斑馬魚(B組),每組10條,禁食24 h,后使用新一代斑馬魚專用游泳速度測試系統(tǒng)(Loligo系統(tǒng),丹麥),參照相關(guān)文獻(xiàn)[11],測量A組和B組斑馬魚的最大游泳速度,為了減少斑馬魚不同個體間BL差異所產(chǎn)生的誤差,將最大游泳速度單位換算為BL/s。
1.2.5斑馬魚胚胎的分組及其處理 依據(jù)文獻(xiàn)[12]對受精后24 h(24hpf)的原基-5期和受精后72 h(72hpf)的突口期斑馬魚胚胎進(jìn)行區(qū)分,同時依據(jù)處理方式不同,將WT斑馬魚胚胎分為空白對照組(C組)、漂白劑處理組(D組)、剪切損傷處理組(E組)和加蘭他敏處理組(F組)。再將WT斑馬魚胚胎和M1型tcap基因純合突變斑馬魚胚胎分為WT斑馬魚胚胎空白對照組(G組)、WT斑馬魚胚胎加蘭他敏處理組(H組)、M1型tcap基因純合突變斑馬魚胚胎空白對照組(I組)和M1型tcap基因純合突變斑馬魚胚胎加蘭他敏處理組(J組),每組30枚胚胎。C、G和I組對72hpf的WT斑馬魚胚胎或者tcap基因純合突變斑馬魚胚胎進(jìn)行正常培養(yǎng),沒有進(jìn)行任何處理;D組用含體積分?jǐn)?shù)0.022 5的漂白劑溶液處理24hpf的WT斑馬魚胚胎2 min,正常培養(yǎng)至72hpf收集胚胎;E組用刀片在顯微鏡下?lián)p傷72hpf的WT斑馬魚胚胎尾部骨骼肌組織,3 h后收集胚胎;F組、H組和J組用濃度為0.5 mmol/L的加蘭他敏溶液處理24hpf的WT斑馬魚胚胎或者tcap基因純合突變斑馬魚胚胎48 h后,收集胚胎。
1.2.6斑馬魚整胚原位雜交及RT-qPCR實驗 參照RONG等[13]報道的方法進(jìn)行原位雜交,檢測C~F組WT斑馬魚胚胎中tcapmRNA顯色情況;對于C~J組斑馬魚胚胎進(jìn)行RT-qPCR檢測:將斑馬魚胚胎于200 μL Trizol試劑中碾碎,然后依次加入Trizol試劑、氯仿、異丙醇、體積分?jǐn)?shù)0.75乙醇溶液,分離并提取總的RNA;采用5×All-In-One-RT-MasterMix反轉(zhuǎn)錄試劑盒進(jìn)行反轉(zhuǎn)錄獲得cDNA,以此cDNA為模板采用SYBR Green Master Mix試劑盒進(jìn)行RT-qPCR反應(yīng),以β-actin作為內(nèi)參照,檢測斑馬魚胚胎中tcap、p53和p21 mRNA的表達(dá)水平。RT-qPCR引物序列見表1。

表1 RT-qPCR引物名稱及其序列
1.3 統(tǒng)計學(xué)處理
使用GraphPad Prism 8.0軟件進(jìn)行數(shù)據(jù)統(tǒng)計分析,計量資料以均數(shù)±標(biāo)準(zhǔn)差表示,兩組間比較采用t檢驗,多組間比較采用單因素方差分析,以P<0.05為差異有統(tǒng)計學(xué)意義。
2 結(jié) 果
2.1 tcap基因純合突變斑馬魚篩選結(jié)果
對設(shè)計合成的sgRNA進(jìn)行核酸電泳,顯示為一條長約100 bp的單一核酸條帶,表明成功合成了斑馬魚tcap基因編輯所需的sgRNA;于顯微鏡下注射sgRNA與Cas9蛋白復(fù)合物對WT斑馬魚胚胎tcap基因進(jìn)行編輯,并最終篩選獲得了兩種基因型不同的tcap基因純合突變斑馬魚,分別為缺失堿基GCGCCAGTC和增加堿基TCTATCTATCT的M1型以及缺失堿基CTGCGCCAGT的M2型(圖1);通過GENEDOC軟件分析發(fā)現(xiàn),這兩種tcap基因純合突變斑馬魚都會產(chǎn)生telethonin截短蛋白,從而導(dǎo)致tcap基因功能喪失。由于M1型tcap基因純合突變斑馬魚所編碼的telethonin截短蛋白氨基酸個數(shù)最少,因此后續(xù)均使用M1型tcap基因純合突變斑馬魚進(jìn)行實驗。
2.2 WT與M1型tcap基因純合突變斑馬魚骨骼肌組織HE染色結(jié)果
HE染色實驗結(jié)果顯示,成年WT斑馬魚的骨骼肌肌原纖維排列規(guī)則(圖2A),而成年M1型tcap基因純合突變斑馬魚的骨骼肌肌原纖維排列明顯紊亂(圖2B)。
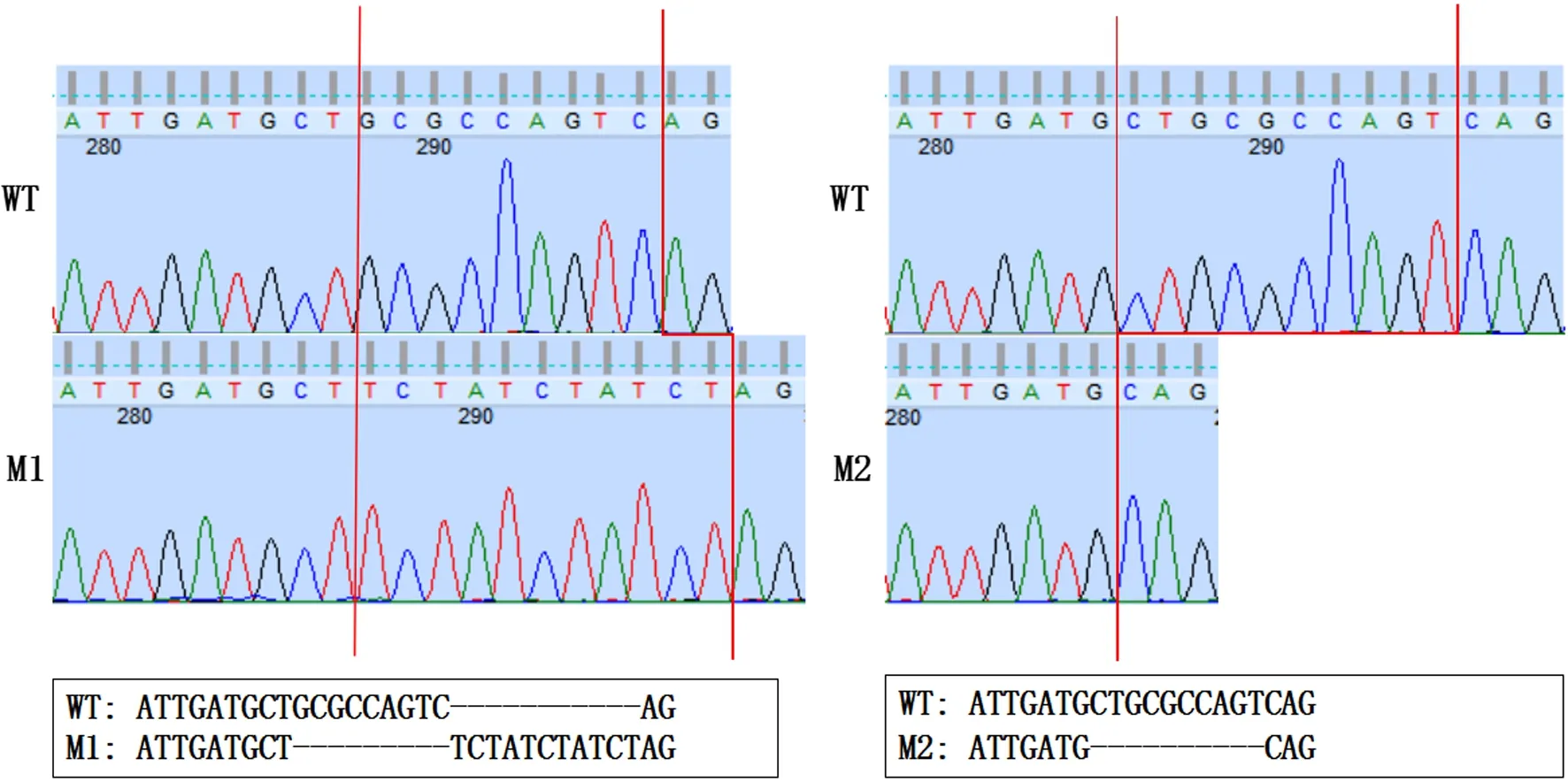
紅色線條標(biāo)記突變區(qū)

A:WT斑馬魚,B:M1型tcap基因純合突變斑馬魚,箭頭指向紊亂的肌原纖維,HE染色,400倍
2.3 WT與M1型tcap基因純合突變斑馬魚最大游泳速度檢測結(jié)果
檢測結(jié)果顯示,A組和B組斑馬魚的最大游泳速度分別為(22.78±2.25)、(18.67±2.95)BL/s,兩組比較差異具有顯著性(t=3.32,P<0.05)。
2.4 不同處理方式下WT斑馬魚胚胎中tcap mRNA表達(dá)情況
原位雜交實驗結(jié)果顯示,與C組比較,D~F組WT斑馬魚胚胎中tcapmRNA的紫色顯色區(qū)域明顯增加(圖3)。RT-qPCR實驗結(jié)果顯示,C~F組WT斑馬魚胚胎中tcapmRNA的相對表達(dá)量分別為1.00±0.00、9.64±0.42、8.20±2.52、9.88±1.78,組間比較差異具有顯著性(F=21.70,P<0.05),其中與C組相比,D~F組tcapmRNA相對表達(dá)量明顯升高(t=4.95~35.44,P<0.05)。
2.5 各組斑馬魚胚胎中p53、p21 mRNA相對表達(dá)量比較結(jié)果
RT-qPCR實驗結(jié)果顯示,G~J組斑馬魚胚胎中p53 mRNA的相對表達(dá)量分別為1.00±0.00、1.03±0.27、0.75±0.11、0.46±0.11,p21 mRNA相對表達(dá)量分別為1.00±0.01、0.82±0.15、0.73±0.04、0.36±0.06,組間比較差異均具有顯著性(F=8.45、33.04,P<0.05);H組與G組比較,斑馬魚胚胎中p53和p21 mRNA相對表達(dá)量均無顯著差異(P>0.05);J組與H、I組比較,斑馬魚胚胎中p53、p21 mRNA相對表達(dá)量明顯下降(t=3.24~8.94,P<0.05)。
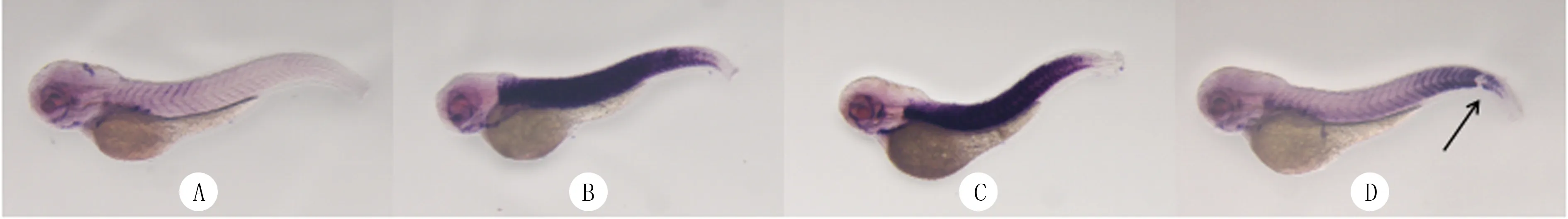
A、B、C、D分別對應(yīng)C、D、E、F組,箭頭指示剪切位點,原位雜交,40倍
3 討 論
在基因編輯過程中,一般先通過基因編輯工具誘導(dǎo)染色體DNA形成一個斷裂缺口,然后細(xì)胞啟動DNA修復(fù);此時DNA斷裂的地方會在修復(fù)過程中插入或者刪去堿基,進(jìn)而導(dǎo)致靶基因產(chǎn)生移碼或缺失突變[14]。CRISPR系統(tǒng)最早在大腸桿菌中被發(fā)現(xiàn)[15]。經(jīng)CRISPR系統(tǒng)改造而來的CRISPR/Cas9基因編輯技術(shù),具有效率高、能對基因組進(jìn)行精確編輯等優(yōu)勢,從而廣泛應(yīng)用于動物模型的構(gòu)建[14]。本研究將CRISPR/Cas9基因編輯技術(shù)應(yīng)用于斑馬魚活體動物模型,成功獲得了tcap基因功能缺失的純合突變斑馬魚。
人類的TCAP基因定位于17號常染色體,其編碼的Telthonin蛋白共由167個氨基酸組成,相對分子質(zhì)量為19 000。該蛋白具有獨特的β-折疊結(jié)構(gòu),存在于肌節(jié)Z盤[16-17]。Telethonin蛋白存在于骨骼肌、心肌和腸胃道平滑肌中,其主要功能是維持肌組織的結(jié)構(gòu)穩(wěn)定性。TCAP的隱性無義或缺失突變會導(dǎo)致LGMD2G。LGMD2G患者會出現(xiàn)遠(yuǎn)端肢體肌肉力量逐漸減弱的情況,最終失去行動能力[2,8]。本研究HE染色實驗結(jié)果顯示,在tcap基因純合突變斑馬魚中能檢測到明顯的骨骼肌肌原纖維紊亂表型,提示tcap基因的功能喪失直接影響了斑馬魚骨骼肌功能,從而支持tcap維持骨骼肌肌肉結(jié)構(gòu)穩(wěn)定性的理論。本研究中斑馬魚最大游泳速度測試結(jié)果顯示,tcap基因功能缺失純合突變斑馬魚的運動能力顯著降低。這些表型與在LGMD2G患者中出現(xiàn)的肌肉萎縮和肌肉力量減弱或缺失的癥狀是基本一致的[6],提示本研究成功構(gòu)建了斑馬魚LGMD2G模型。
根據(jù)文獻(xiàn)報道,漂白劑處理導(dǎo)致的機械刺激會使斑馬魚胚胎骨骼肌組織機械壓力過載,從而導(dǎo)致tcap基因相對表達(dá)量增加[18]。加蘭他敏是一種乙酰膽堿酯酶抑制劑,能夠使肌肉組織受到生物刺激;剪切損傷處理會使斑馬魚胚胎骨骼肌組織產(chǎn)生損傷應(yīng)激反應(yīng)。為了進(jìn)一步探討tcap基因突變導(dǎo)致斑馬魚LGMD2G的致病機制,本研究檢測了WT斑馬魚的tcap基因在以上處理方式下的表達(dá)情況,原位雜交和RT-qPCR實驗結(jié)果顯示,除漂白劑處理外,加蘭他敏處理和剪切損傷處理也會誘導(dǎo)WT斑馬魚胚胎tcapmRNA相對表達(dá)量增加。MDM2蛋白既能降解P53蛋白又能降解Telethonin蛋白[19-20],提示Telethonin蛋白可能參與了P53信號通路的調(diào)控。本研究的RT-qPCR實驗結(jié)果顯示,tcap基因純合突變斑馬魚胚胎經(jīng)加蘭他敏處理后,其骨骼肌中p53以及p21 mRNA相對表達(dá)量明顯降低,提示斑馬魚LGMD2G的發(fā)病機制很可能與tcap基因缺失后肌組織營養(yǎng)不良導(dǎo)致p53信號通路異常有關(guān)。
研究發(fā)現(xiàn),Telethonin能與肌LIM蛋白(MLP)進(jìn)行相互作用,而小鼠MLP蛋白表達(dá)降低會誘發(fā)肥厚型心肌病和心力衰竭[21]。TCAP基因缺失是否會影響MLP蛋白功能導(dǎo)致骨骼肌功能異常,從而誘發(fā)LGMD2G還有待進(jìn)一步研究。除此之外,多種信號通路相關(guān)蛋白如肌聯(lián)蛋白、骨形態(tài)發(fā)生蛋白10和離子通道蛋白Nav1.5等都能與Telethonin蛋白等進(jìn)行相互作用[22-25],這些蛋白是否也參與了LGDM2G的發(fā)生發(fā)展目前尚無研究報道,表明LGMD2G的致病機制研究還處于初步探索階段。
總之,本研究首次成功構(gòu)建了LGMD2G的斑馬魚疾病模型,并初步發(fā)現(xiàn)tcap基因功能缺失導(dǎo)致p53基因下調(diào)可能是斑馬魚LGMD2G的致病機制之一。但是關(guān)于斑馬魚tcap基因通過p53基因如何調(diào)控下游相關(guān)信號通路進(jìn)而導(dǎo)致LGMD2G還有待進(jìn)一步研究。本研究為LGMD2G治療藥物的篩選提供了一定的理論基礎(chǔ)。
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
中國醫(yī)學(xué)影像學(xué)雜志(2021年6期)2021-08-13 08:43:36
中國生殖健康(2020年2期)2021-01-18 02:51:26
小學(xué)生導(dǎo)刊(2018年13期)2018-06-29 03:49:00
中國生殖健康(2018年2期)2018-01-12 13:57:51
現(xiàn)代檢驗醫(yī)學(xué)雜志(2016年4期)2016-11-15 02:01:14
中國現(xiàn)代醫(yī)學(xué)雜志(2015年26期)2015-12-23 11:04:22
鄭州大學(xué)學(xué)報(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:44
中華皮膚科雜志(2014年4期)2014-12-19 12:55:49
中國神經(jīng)精神疾病雜志(2014年1期)2014-03-01 03:23:22
