基于轉錄組測序分析壯骨止痛方治療去勢大鼠骨質疏松癥的基因表達差異
2021-10-09 08:53:48陳紅瓊楊珊珊陳瑤郁潔雷曉明
中醫藥信息 2021年6期
關鍵詞:差異
陳紅瓊,楊珊珊,陳瑤,郁潔,雷曉明
(湖南中醫藥大學,湖南 長沙 410208)
骨質疏松癥是一種以骨量減少,骨組織微結構損壞,導致骨脆性增加,易發生骨折為特征的全身代謝性骨骼疾病[1]。其主要臨床表現為疼痛、腰膝酸軟、脊柱變形、易于骨折[2]。研究表明,骨骼代謝受到許多因素調節,包括激素、年齡、遺傳以及環境等[3]。其主要發病機制是成骨細胞與破骨細胞之間骨重建過程的失衡,骨形成與骨吸收兩者之間的失衡[4],但目前骨質疏松癥發病的分子機制尚不明確。骨質疏松癥沒有根治的方法,現代臨床醫學治療原發性骨質疏松癥普遍應用抑制骨吸收類藥物、促骨形成類藥物和骨礦化類藥物,這些藥物在臨床上取得了顯著的治療效果,但長期應用會帶來較多不良反應[5]。
壯骨止痛方是湖南省名中醫莫新民教授的經驗方,以古方補骨脂丸為基礎化裁而來,由淫羊藿、補骨脂、女貞子、枸杞、骨碎補、牛膝和狗脊組成,具有補益肝腎、壯骨止痛功效。前期研究已證明壯骨止痛方可以多途徑抑制骨吸收,促進骨代謝,提高雌激素水平,對于絕經后骨質疏松癥具有良好的防治作用[6-7]。但其治療骨質疏松癥的具體機制仍不清楚,因此,本研究基于轉錄組學測序分析壯骨止痛方治療去勢大鼠骨質疏松癥的差異表達基因以及對相關通路的影響,尋找相關的基因靶點,以期為臨床應用壯骨止痛方提供實驗依據。
1 材料
1.1 主要儀器
KL05A 型醫用離心機(湖南凱達科學儀器有限公司);991 型超低溫冰箱(賽默飛世爾科技有限公司);Illumina 測序平臺(廣州市唯譽智合科技有限公司)。
1.2 藥品與試劑
壯骨止痛膠囊(四川美大康藥業股份有限公司,批準文號:Z20050118);戊巴比妥鈉(YaErbio公司,批號:20200422);青霉素鈉(華北制藥股份有限公司,批號:F9102109);紅細胞裂解液(合肥志宏生物技術有限公司,批號:700577116);大鼠淋巴細胞分離液試劑盒(北京索萊寶科技有限公司,批號:20200616)。
1.3 動物
SPF 級雌性SD 大鼠12 只,體質量180~220 g,購自湖南斯萊克景達實驗動物有限公司,動物合格證號(1107272011003527),于湖南中醫藥大學動物實驗中心飼養,許可證號:SCXK(湘)2019-0009,飼養室溫度24~26 ℃,相對濕度50%~70%。人工12 h 晝/夜循環照明,分籠飼養。每日定時清洗籠舍,大鼠能自由攝食及飲水。實驗前適應性喂養1 周,造模前禁食12 h,自由飲水。
2 方法
2.1 分組、造模與給藥
將12 只SD 大鼠隨機分為實驗組、模型組,每組6 只。兩組均采用國內外公認的雌性大鼠去卵巢3 個月造成絕經后骨質疏松模型[8-9]。方法:大鼠用2%戊巴比妥鈉(0.2 mL/100 g)腹腔麻醉,在無菌條件下,從大鼠腰椎兩側摘除雙側卵巢。術后連續3 d 大腿肌內注射青霉素納,每只大鼠4 萬U/天,術后5 d 拆線。術后1 周開始灌胃給藥,實驗組給予壯骨止痛膠囊(567 mg/kg),模型組給予同體積的蒸餾水(1 mL/150 g),每日1次,連續13周。
2.2 樣本制備
13 周后,所有大鼠腹腔注射2% 戊巴比妥鈉(0.2 mL/100 g)麻醉,經腹腔主靜脈取血后,2 h內用大鼠淋巴細胞分離液試劑盒按說明收集PBMC,紅細胞裂解液裂解PBMC 中殘余紅細胞,隨機將各組每2 只大鼠的PBMC 合并為1 個樣本,每組3 個樣本,于液氮速凍后放-80 ℃保存,備用。
2.3 RNA提取及測序
將兩組的PBMC 樣本,共6 個樣本,送至廣州市唯譽智合科技有限公司進行RNA 提取、建庫及轉錄組測序,測序平臺為Illumina測序平臺。
3 信息分析
3.1 差異表達基因分析
采用DEGSeq 分析差異表達基因,差異基因篩選主要參考差異倍數(Fold change 值)以及q值(Padj 值,矯正之后的Pvalue 值)作為相關指標,通常選取|log2 Fold change|≥1 和q<0.05 的差異基因作為顯著差異基因。
3.2 GO和KEGG富集分析
基于篩選出的差異表達基因(|log2 Fold change|≥1,q<0.05),應用超幾何檢驗對差異表達基因進行GO和KEGG 富集分析,以校正后的q<0.05 為閾值,滿足此條件的GO詞條和KEGG通路被認為具有顯著性。
4 結果
4.1 差異表達基因分析結果
實驗共獲得20 276 個轉錄組mRNA,與模型組相比,實驗組共有10 166 個表達上調,10 110 個表達下調,見圖1。為差異表達基因及樣本垂直聚類結果的熱圖。DEGs 分析顯示,差異明顯的基因有149 個(|log2 Fold change|≥1,q<0.05),67個基因上調,82個基因下調(見圖2);進一步篩選顯著差異基因(|log2 Fold change| ≥2,q<0.01),共選出18 個顯著差異基因,其中上調基因15 個,下調基因3 個,具體信息見表1。

表1 實驗組與模型組外周血單個核細胞的差異表達基因
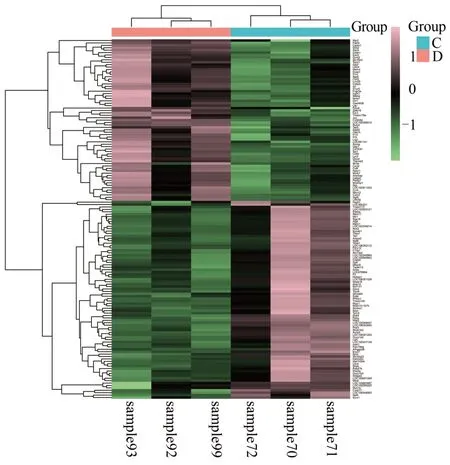
圖1 實驗組與模型組差異基因的熱圖
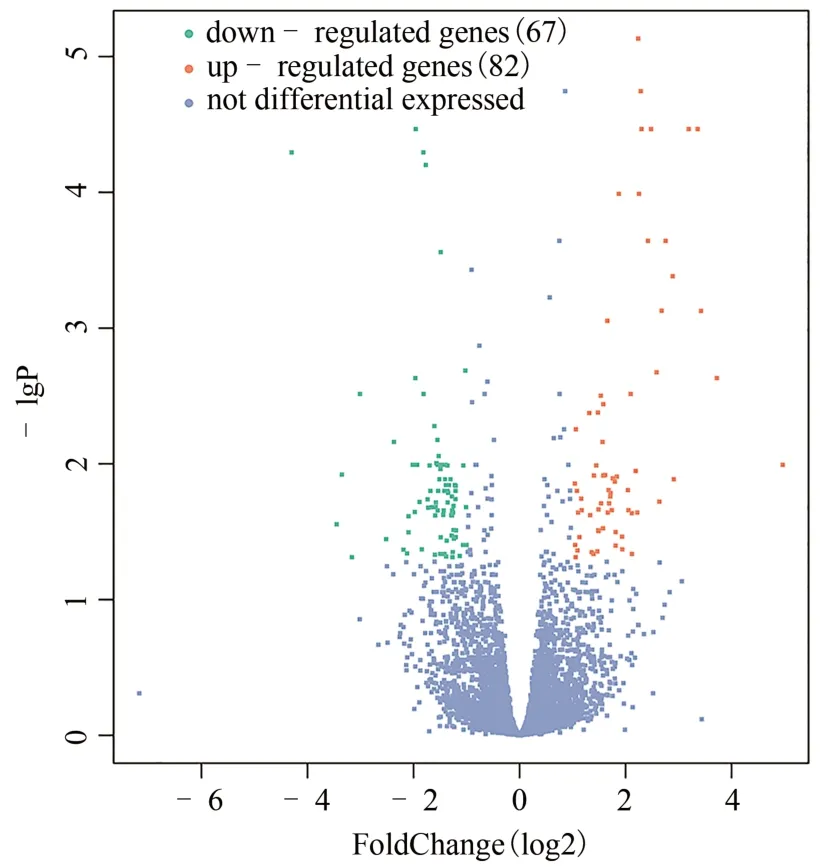
圖2 實驗組與模型組差異表達基因的火山圖
4.2 差異表達基因的GO富集分析
對差異基因(|log2 Fold change|≥1,q<0.05)進行GO 富集分析,結果顯示有45 個富集項(見圖3),差異表達基因主要富集在細胞部分、膜部分、細胞器、細胞過程、生物調節、代謝過程、綁定分子類、催化活性等;進一步校正檢驗后的P值之后,以q<0.05 為閾值,發現滿足此條件的差異表達基因中顯著富集的GO 條目有316 條,功能富集到生物過程的GO 條目有303 條(見表2,取顯著性最高的前10條)、細胞組分有4條(見表3)、分子功能有9條(見表4)。

表2 差異表達基因顯著性GO富集結果-生物過程
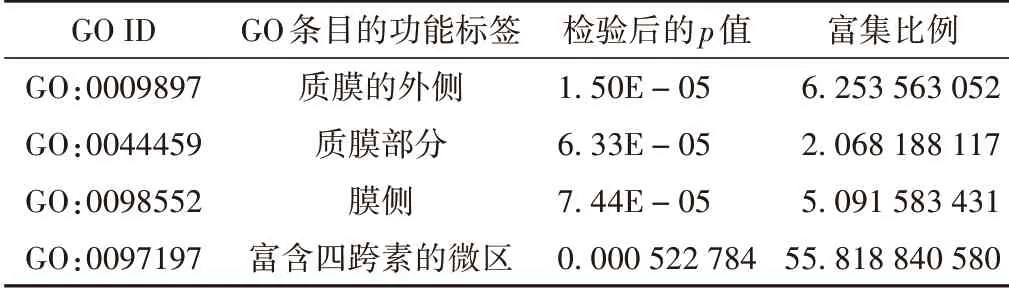
表3 差異表達基因顯著性GO富集結果-細胞組分

表4 差異表達基因顯著性GO富集結果-分子功能
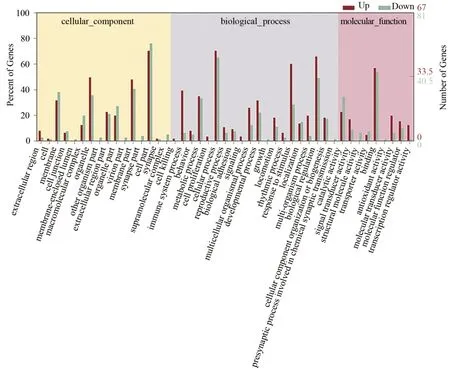
圖3 實驗組與模型組差異表達基因的GO統計柱狀圖
4.3 差異表達基因的KEGG富集分析
對差異基因(|log2 Fold change|≥1,q<0.05)進行KEGG富集分析,共富集到170條生物通路,結果顯示:上調基因主要參與細胞因子與細胞因子受體的相互作用、MAPK 信號通路、TNP 信號通路、吞噬體、趨化因子信號通路、癌癥通路、癌癥轉錄調控失調等通路(見圖4);下調基因主要參與嘌呤代謝、內質網蛋白加工、cGMP-PKG信號通路、黏著斑、IL-17信號通路、軸突引導等通路(見圖5);進一步校正檢驗P值之后,以q<0.05 為閾值,得到TNP 信號通路、IL-17 信號通路、沙門氏菌感染三條顯著性富集的通路見表5。
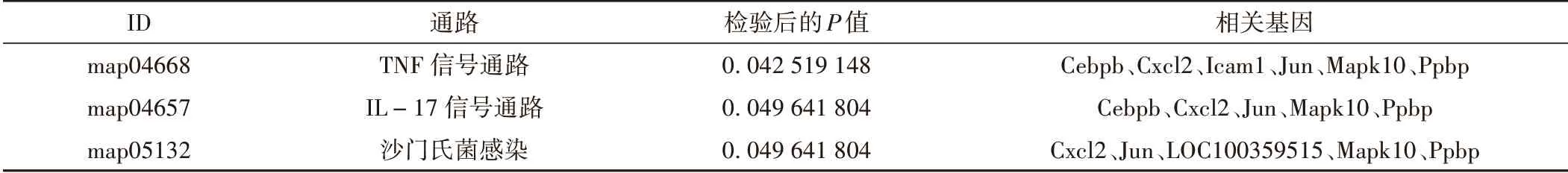
表5 差異表達基因的顯著性富集通路
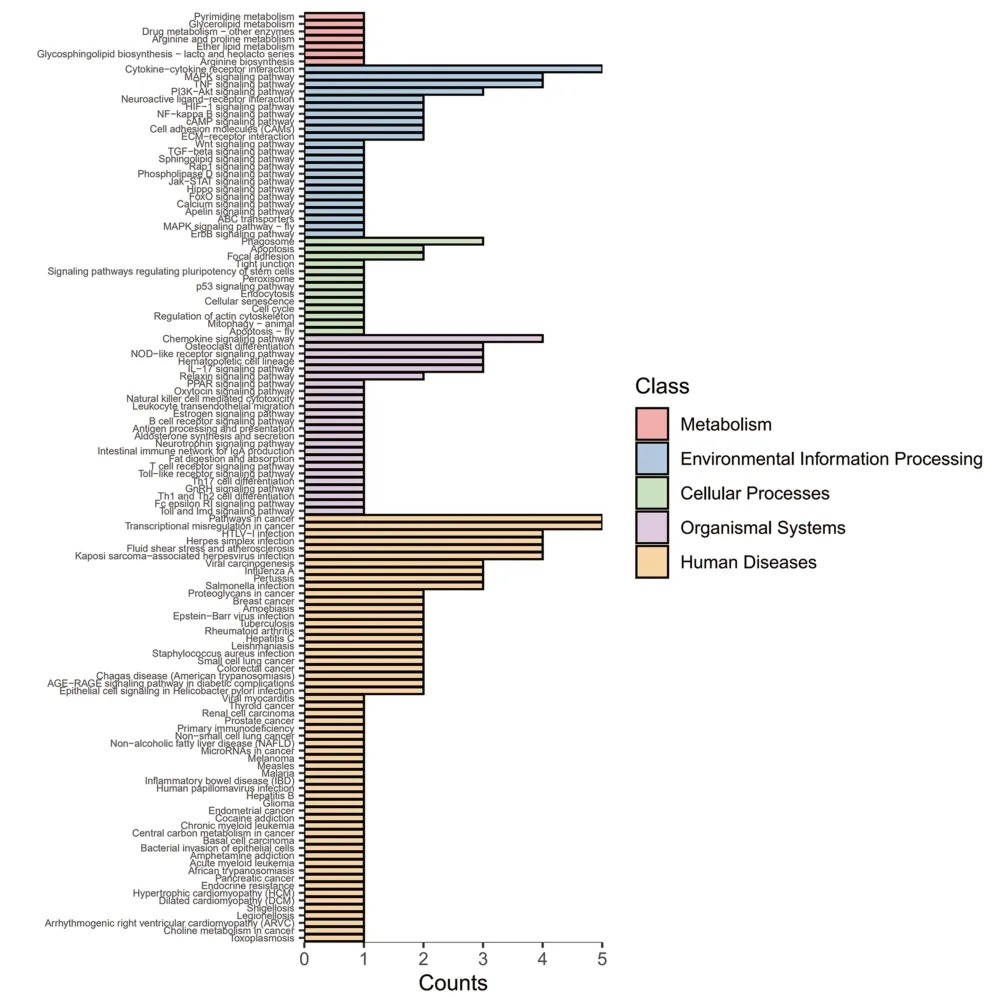
圖4 上調差異表達基因的KEGG分類
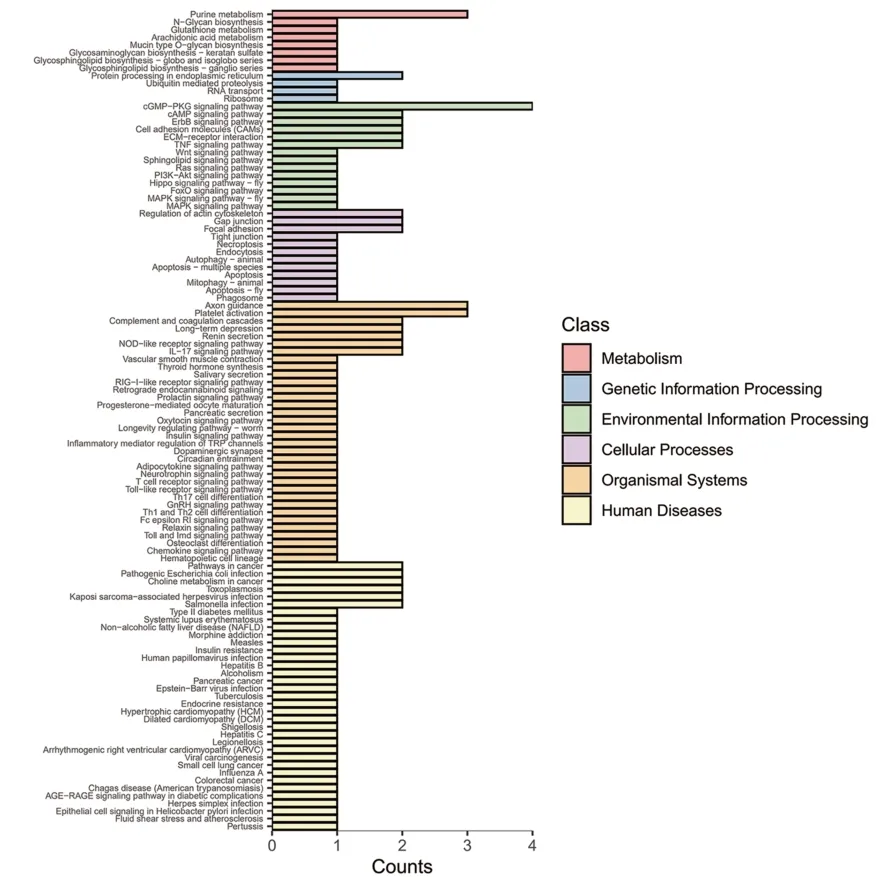
圖5 下調差異表達基因的KEGG分類
5 討論
本研究采用高通量測序技術對去勢大鼠骨質疏松模型組與使用壯骨止痛方干預的實驗組的外周血單個核細胞進行轉錄組學測序,差異表達基因分析結果表明,實驗組與模型組差異表達基因有149 個,以|log2 Fold change| ≥1 和q<0.05 作為篩選條件,上調基因有67 個,下調基因有82 個。GO 富集分析涉及生物過程、細胞組分以及分子功能三大類,差異表達基因主要富集在細胞部分、膜部分、細胞器、細胞過程、生物調節、代謝過程、綁定分子類和催化活性等;校正P值后,在生物過程分類中,細胞遷移、免疫系統過程、防御反應所占比例最多,其次是應激反應、調節免疫系統的過程、細胞運動性、調節對外界刺激的反應、炎癥反應等;在細胞組分分類中,細胞膜部分所占比列最多;在分子功能分類中,細胞因子受體活性所占比列最多,其次是整聯蛋白結合及受體結合類分子功能。KEGG 富集分析結果顯示,顯著性富集的通路為TNF 信號通路、IL-17 信號通路、沙門氏菌感染三條通路,涉及相關差異基因有Cebpb、Cxcl2、Icam1、Jun、Mapk10、Ppbp、LOC100359515。研究發現,腫瘤壞死因子成員之一(TNF-α)可以通過NF-κB 信號轉導通路促進成骨細胞的分化和細胞外基質的礦化[10],白介素17(IL-17)可以顯著抑制骨保護素(OPG)和增強NF-κB 配體受體活化劑(RANKL),并且IL-17 可以通過TNF 受體相關因子6(TRAF6)、TANK 結合激酶1(TBK1)、c-Jun N 端激酶(JNK)或NF-κB 調節RANKL 和OPG 的表達[11],而RANKL/RANK/OPG 系統是調節破骨細胞分化、成熟和凋亡最重要的信號通路[12]。但是暫無沙門氏菌感染在骨質疏松領域的相關報道。
在差異表達基因分析中,差異表達基因主要與脂質代謝相關疾病、骨質疏松疾病、炎癥、腫瘤等疾病有關。如顯著性最高的G0S2,其參與機體內脂質代謝過程,與肥胖、糖尿病胰島素抵抗等脂質代謝相關疾病密切相關[13];Dgat2在脂質代謝旺盛的組織中的表達水平最高,如肝臟、腎臟、白色脂肪組織等[14];Fpr3參與多種生理和病理過程,包括防御反應、炎癥、神經退行性疾病、腫瘤、糖脂代謝紊亂相關疾病等[15];Nefh 與肌肉萎縮,阿爾茨海默病、帕金森綜合征有關;Niacr1 可使小鼠結腸炎和結腸癌風險增加。但目前以上幾種基因在骨質疏松領域還沒有相關報道,其表達是否會影響骨質疏松癥的發生還需進一步驗證。
已有骨質疏松相關報道的差異基因如Ccrl2 參與了chemerin 信號通路調節,研究發現,chemerin 與其受體CMKLR1、Ccrl2 在骨髓充質干細胞和前成骨細胞的成脂化或成骨化的過程中起就決定性作用[16],但目前有關Ccrl2 在骨質疏松領域的報道很少;Csf3r 是一種蛋白質編碼基因,其在破骨細胞上表達,在成骨細胞上不表達,與骨質疏松癥的形成有一定關系[17],但在慢性中性粒細胞白血病領域的研究更常見;LOC100359515 是誘導型一氧化氮合酶(iNOS)編碼基因,在骨質疏松癥的發病機制中起著重要作用。iNOS 產生一氧化氮(NO),可導致骨骼形成和雌激素缺失引起的骨吸收之間的不平衡[18],這可能是治療骨質疏松癥的重要靶點;Nlrp12 通過抑制NF-κB 誘導的破骨形成在骨中發揮保護作用,并在破骨刺激下下調,調節Nlrp12 水平可控制OC 前體中NF-κB 信號,改變骨穩態和骨溶解反應[19];破骨細胞相關受體基因Oscar 在破骨細胞分化中起關鍵作用,可以使骨密度降低,是調控骨密度的重要候選基因[20]。在本研究中Nlrp12、Oscar 是上調基因,其可能是壯骨止痛方治療骨質疏松癥的關鍵靶點。
目前,關于骨質疏松癥的具體發病機制尚不明確,有關骨質疏松癥的分子機制研究也比不多,并且沒有根治的方法。近年來隨著科學技術的迅速發展,高通量測序技術已經廣泛應用于基因表達轉錄水平研究,是研究疾病基因功能的重要手段之一。本研究通過基因差異表達分析和富集分析,篩選了壯骨止痛方干預去勢大鼠骨質疏松癥相關的基因和通路,但由于樣本量不多,隨機誤差和假陽性難以排除,需要進一步研究,且本研究是通過高通量測序結合基因數據庫進行生物信息學的預測,還需后續的實驗去驗證。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
