濃香型白酒包包曲微生物種群多樣性及形成機制
2021-10-09 00:41:12張倩韓保林李子健謝軍余東鄒永芳郭輝祥文靜張玲玲羅惠波黃丹
食品與發(fā)酵工業(yè) 2021年18期
關(guān)鍵詞:差異
張倩,韓保林,2,李子健,謝軍,2,余東,鄒永芳,郭輝祥,文靜,張玲玲,羅惠波,2*,黃丹,2*
1(四川輕化工大學(xué) 生物工程學(xué)院,四川 自貢,643000)2(釀酒生物技術(shù)及應(yīng)用四川省重點實驗室,四川 宜賓,644000)3(舍得酒業(yè)股份有限公司,四川 遂寧,629209)
包包曲,是濃香型白酒釀造過程中重要的糖化發(fā)酵劑[1],因其塊狀結(jié)構(gòu)一側(cè)具有較大凸起而得名。獨特的“包包”結(jié)構(gòu)使包包曲比一般大曲擁有更大的比表面積,更有利于霉菌等的生長[2]。同時,其外緊內(nèi)松的結(jié)構(gòu)特點[2],增大了物料的孔隙率,更有利于大曲內(nèi)部和外部的物質(zhì)能量交換和微生物的生長繁殖。包包曲獨特的結(jié)構(gòu)使熱量、水分和氧氣含量等參數(shù)從曲塊外部到內(nèi)部呈梯度分布[3],造成大曲不同部位微生物群落多樣性,從而導(dǎo)致了曲皮、曲心微生物群落的差異。
研究表明,大曲不同部位往往具有不同的質(zhì)量[4]和微生物群落結(jié)構(gòu),其差異甚至大于同一環(huán)境下不同制曲方式[5]。不同部位大曲的研究對加深大曲微生物群落的理解有著重要作用。曲皮、曲心的理化、生化特性和基于傳統(tǒng)可培養(yǎng)方式的微生物群落特征及差異已經(jīng)取得了一定的研究成果[4,6]。隨著高通量測序技術(shù)被普遍應(yīng)用于白酒釀造研究[7],曲皮、曲心微生物群落結(jié)構(gòu)與差異被進一步解析。JIN等[8]通過Illumina MiSeq平臺測序發(fā)現(xiàn)醬香型大曲中乳桿菌目(Lactobacillales)在曲皮中豐度更高,而芽孢桿菌目(Bacillales)則在曲心中豐度更高。近期,CHEN等[9]也通過不可培養(yǎng)的手段得出特香型大曲曲皮和曲心微生物間存在顯著差異的結(jié)論。然而,運用高通量測序技術(shù)對曲皮、曲心在大曲發(fā)酵過程中的微生物群落變化情況和差異的報道目前幾乎沒有。
本研究采用高通量測序技術(shù)探究了包包曲發(fā)酵過程中的曲皮和曲心微生物群落結(jié)構(gòu)變化規(guī)律,并與理化因子相關(guān)聯(lián),旨在比較大曲發(fā)酵過程中曲皮、曲心微生物群落的差異及與理化因子的關(guān)系,有助于進一步認識濃香型白酒包包曲微生物種群多樣性,探明其形成機制。
1 材料與方法
1.1 材料與試劑
樣品:濃香型包包曲樣品源自四川某濃香型白酒廠,分別在發(fā)酵第0、1、2、3、4、5、6、7、8、9、10、11、12、13、15、16、20、23、29天取樣。每次取樣選擇3塊大曲,將大曲表面約1 cm的部分劃為曲皮(QP),其余部分為曲心(QX),分別粉碎、混勻、裝袋后,于4 ℃和-80 ℃保存。
試劑:NaOH(分析純),成都市科龍化工試劑廠;乙二胺四乙酸二鈉(ethylenediaminetetraacetic acid disodium salt,Na2EDTA)、Na2HPO4、NaH2PO4、蝸牛酶、溶菌酶、十六烷基三甲基溴化銨(hexadecyl trimethyl ammonium bromide,CTAB)、十二烷基硫酸鈉(sodium dodecyl sulfate,SDS),生工生物工程(上海)股份有限公司;CHCl3(分析純),成都市科隆化學(xué)品有限公司。
1.2 儀器與設(shè)備
PT100溫度傳感器,華巨溫度傳感器有限公司;DHG-9245電熱干燥箱,上海一恒科學(xué)儀器有限公司;QL-861漩渦振蕩器,江蘇海門市麒麟醫(yī)用儀器廠;LYNX 6000高速冷凍離心機,美國Thermo公司。
1.3 包包曲理化指標(biāo)的測定
包包曲的水分、總酸、pH值采用大曲通用分析方法QB/T 4257—2011[10]測定。包包曲溫度采用溫度傳感器實時測定。
1.4 包包曲總DNA提取和測序
采用CTAB改良法提取大曲總DNA[11]。使用引物338F/806R(5′-ACTCCTACGGGAGGCAGCA-3′/5′-GGACTACHVGGGTWTCTAAT-3′)對細菌16S rRNA基因的V3~V4高變區(qū)進行擴增,內(nèi)部轉(zhuǎn)錄間隔區(qū)(internal transcribed spacer,ITS)5F/ITS1R(5′-GGAAGTAAAAGTCGTAACAAGG-3′/5′-GCTGCGTTC TTCATCGATGC-3′)對真菌的ITS1進行擴增。由上海派森諾生物科技有限公司在Illumina MiSeq 2500平臺上進行雙端測序。
1.5 包包曲微生物群落結(jié)構(gòu)分析
按照QIIME 2標(biāo)準(zhǔn)流程對原始序列進行處理。使用DADA2對序列進行質(zhì)量過濾、去噪、合并和去除嵌合體[12],使用多樣性插件計算α多樣性指數(shù)。在R(version 4.0.3)的vegan(version 2.5-7)軟件包中進行了層次聚類分析、相似性分析(analysis of similarities,ANOSIM)、mantel檢驗和理化因子與優(yōu)勢微生物的Pearson相關(guān)性分析。mantel test分析了基于bray-curtis距離的細菌、真菌ASV(amplicon sequence variant)相對豐度的距離矩陣和基于歐幾里得距離的理化因子相異性矩陣的Spearman相關(guān)性。使用canoco 4.5進行包包曲微生物群落與環(huán)境因子的冗余分析(redundancy analysis,RDA)。通過STAMP(2.1.3)進行Fisher′s精確檢驗得到兩樣本的顯著差異微生物。而線性判別分析效應(yīng)值(linear discriminant analysis effect size,LEfSe)是通過http://huttenhower.sph.harvard.edu/galaxy/root?tool_id=PICRUSt_normalize網(wǎng)站完成的。
2 結(jié)果與分析
2.1 包包曲發(fā)酵過程中曲皮、曲心理化因子的變化特點
溫度、水分、酸度、pH值是大曲發(fā)酵過程的重要參數(shù),在一定程度上可以反映大曲微生物的生長環(huán)境或代謝情況。曲心溫度符合“前緩中挺后緩落”的變化特點;曲皮溫度受到翻曲的影響,在第7~12天和第16~18 天有明顯的降低(圖1-a)。曲皮、曲心水分含量呈降低趨勢,并分別在第13~29天和第1~6天變化較小;出房時,曲皮、曲心水分含量分別為12.32%和16.23%。發(fā)酵初期,曲塊水分含量較高,霉菌菌絲沒有大量形成,曲塊處于缺氧環(huán)境。因此,微生物無氧發(fā)酵產(chǎn)生大量酸,尤其是曲心中總酸含量較高,曲皮、曲心pH值在第0~2天迅速下降,之后由于有機酸參與合成酯類及原料中蛋白質(zhì)的降解,導(dǎo)致了pH值的增加[13]。包包曲的塊狀結(jié)構(gòu)導(dǎo)致其水分、溫度等理化指標(biāo)從內(nèi)到外呈梯度變化,進而導(dǎo)致大曲發(fā)酵過程中,曲皮、曲心的理化因子存在明顯差異,并且這種差異是從發(fā)酵第2天開始的。
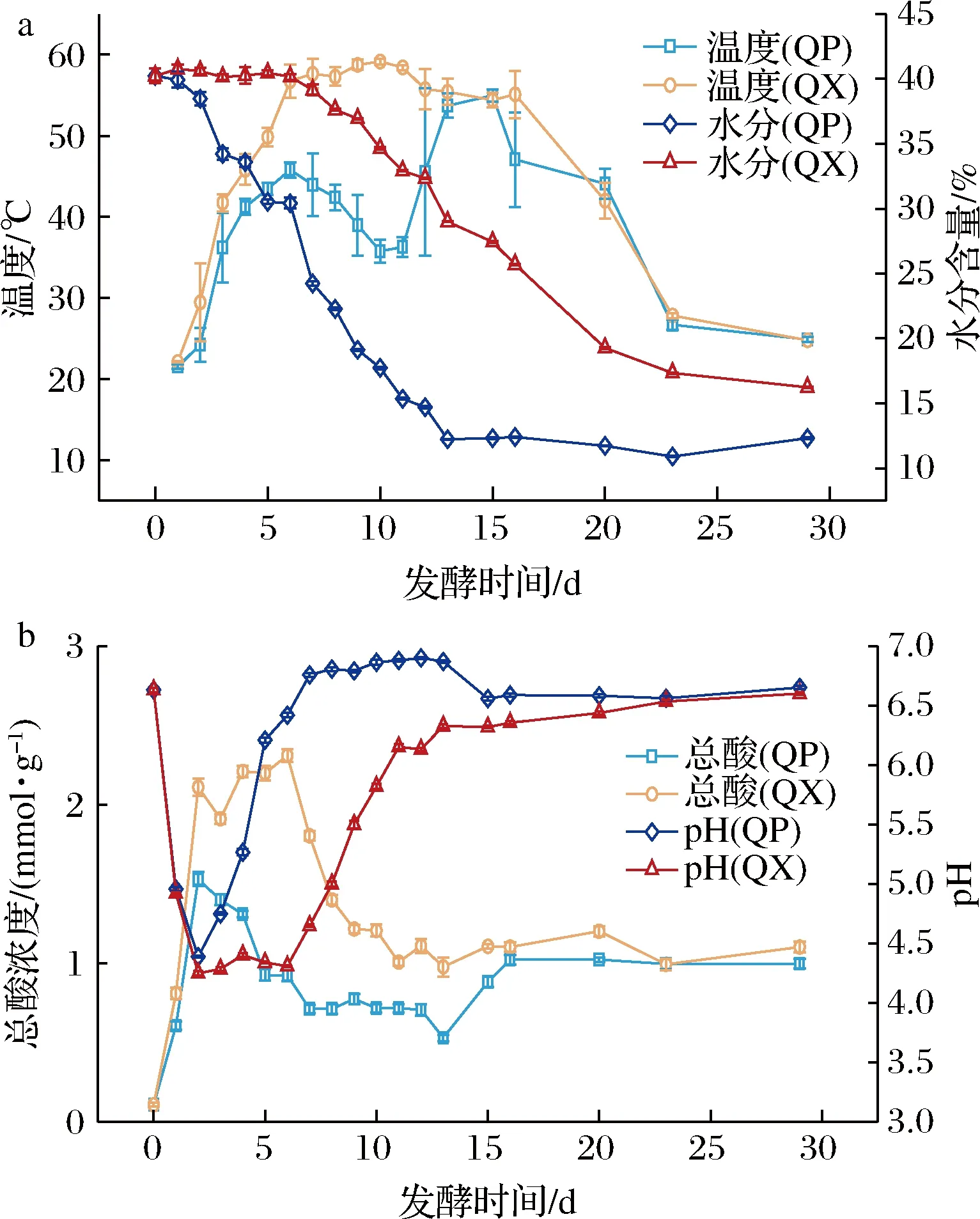
a-包包曲溫度和水分含量;b-包包曲總酸和pH值圖1 包包曲發(fā)酵過程中理化因子的變化規(guī)律Fig.1 Dynamics of physicochemical factors during Baobaoqu fermentation process
2.2 曲皮、曲心微生物群落測序深度評價和α多樣性分析
2.2.1 包包曲樣本微生物群落測序深度評價
Shannon指數(shù)稀釋曲線顯示在10 000條序列時趨于飽和(圖2),表明所有樣本的測序深度均滿足分析要求。
a-細菌;b-真菌圖2 包包曲樣品稀釋曲線Fig.2 Rarefaction curves of Baobaoqu samples
2.2.2 包包曲發(fā)酵過程中曲皮、曲心微生物群落α多樣性分析
包包曲發(fā)酵過程中,曲皮、曲心細菌群落豐富度指數(shù)Chao1和群落多樣性指數(shù)Shannon具有先降低再增高趨勢(圖3-a、圖3-b)。曲皮細菌豐富度和多樣性與曲心沒有顯著差異(P>0.05)。曲皮真菌豐富度和多樣性在第1~9天持續(xù)下降;而曲心真菌豐富度和多樣性在發(fā)酵第1~3天迅速上升,在第3天達到最大值(165.56和5.90);第7~29天,曲皮真菌豐富度和多樣性變化趨勢與曲心一致,且顯著高于曲心(P<0.05)(圖3-c、圖3-d)。
2.3 包包曲發(fā)酵過程中曲皮、曲心微生物群落演替規(guī)律
細菌16S rRNA和真菌ITS測序分別獲得了1 467 670 個和1 350 213個高質(zhì)量序列,按100%相似度聚類分別得到4 454個和504個ASV,覆蓋率均在99%以上。細菌、真菌ASV特征序列分別與Greengenes數(shù)據(jù)庫、UNITE數(shù)據(jù)庫比對進行物種注釋,共檢測到475個細菌屬和69個真菌屬。
包包曲發(fā)酵過程中,曲皮、曲心的主要細菌目為Bacillales、Lactobacillales、假單胞菌目(Pseudomonadales)(平均相對豐度>10%)。如圖4-a和圖4-b所示,曲皮、曲心優(yōu)勢細菌屬(平均相對豐度>1%)在包包曲發(fā)酵過程中的變化趨勢較為相似,但豐度具有明顯差異,特別是在第13~29天。發(fā)酵第1~6天,Lactobacillales各屬和假單胞菌屬(Pseudomonas)在群落中占優(yōu)勢;隨著溫度升高,嗜熱菌芽孢桿菌屬(Bacillus)、熱放線菌屬(Thermoactinomyces)在第7~12天豐度最高,相對豐度占70%以上;第13天之后,曲皮群落優(yōu)勢微生物為Bacillus、Thermoactinomyces、Pseudomonas、魏斯氏菌屬(Weissella)、明串珠菌屬(Leuconostoc)、不動桿菌屬(Acinetobacter)、未分類的根瘤菌屬(unclassifiedRhizobiaceae)等,而曲心中仍以Bacillus、Thermoactinomyces和Pseudomonas為主。并且,曲皮、曲心中的Bacillus分別在第3天和第8天達到優(yōu)勢豐度(相對豐度>1%),這可能是由于曲心中高酸度、高水分的環(huán)境抑制了酸敏感的Bacillus[14]的生長繁殖。
包包曲發(fā)酵過程中,曲皮、曲心的主要真菌目為散囊菌目(Eurotiales)和毛霉目(Mucorales)。而優(yōu)勢真菌屬在曲皮、曲心中的分布具有明顯差異,如圖4-c 和圖4-d所示。嗜熱子囊菌屬(Thermoascus)在曲心中更早出現(xiàn)優(yōu)勢,曲皮中根霉屬(Rhizopus)的豐度在第1~6天遠大于曲心。發(fā)酵第13~29天,曲皮中優(yōu)勢微生物除Thermoascus外,還有Rhizopus、根毛霉屬(Rhizomucor)、曲霉屬(Aspergillus)、畢赤酵母屬(Pichia)、嗜熱真菌屬(Thermomyces),而曲心中僅Thermoascus和Thermomyces在群落中占優(yōu)勢。
由圖4可知,出房時,曲皮、曲心微生物群落具有不同的結(jié)構(gòu)特點,曲皮微生物群落由多菌主導(dǎo),而曲心微生物群落主要由Pseudomonas和嗜熱菌Thermoactinomyces、Thermoascus、Thermomyces主導(dǎo)。Bacillus、Rhizopus、Rhizomucor是大曲中主要糖化酶和淀粉酶產(chǎn)生菌[15],其在曲皮中的優(yōu)勢可能導(dǎo)致了曲皮糖化力、液化力大于曲心。同時,這些優(yōu)勢微生物也是白酒釀造過程中的重要功能微生物。其中,Bacillus普遍具有良好的產(chǎn)淀粉酶和蛋白酶的功能,能夠代謝多種風(fēng)味物質(zhì)[16]。Thermoascus具有產(chǎn)熱穩(wěn)定性和高活力纖維素酶、蛋白酶的功能,具有產(chǎn)酒生香作用[11]。而Pseudomonas是大曲和糟醅的優(yōu)勢微生物[17],可以產(chǎn)生嗜熱脂肪酶[18]并抑制谷物病原體[19],也可能對發(fā)酵產(chǎn)生負面影響[20],其在釀造系統(tǒng)中的功能及影響需要進一步研究。曲皮、曲心中這些重要優(yōu)勢微生物的豐度差異,賦予了曲皮、曲心不同的微生物群落及代謝特點,也是曲皮、曲心理化因子存在較大差異的原因之一。
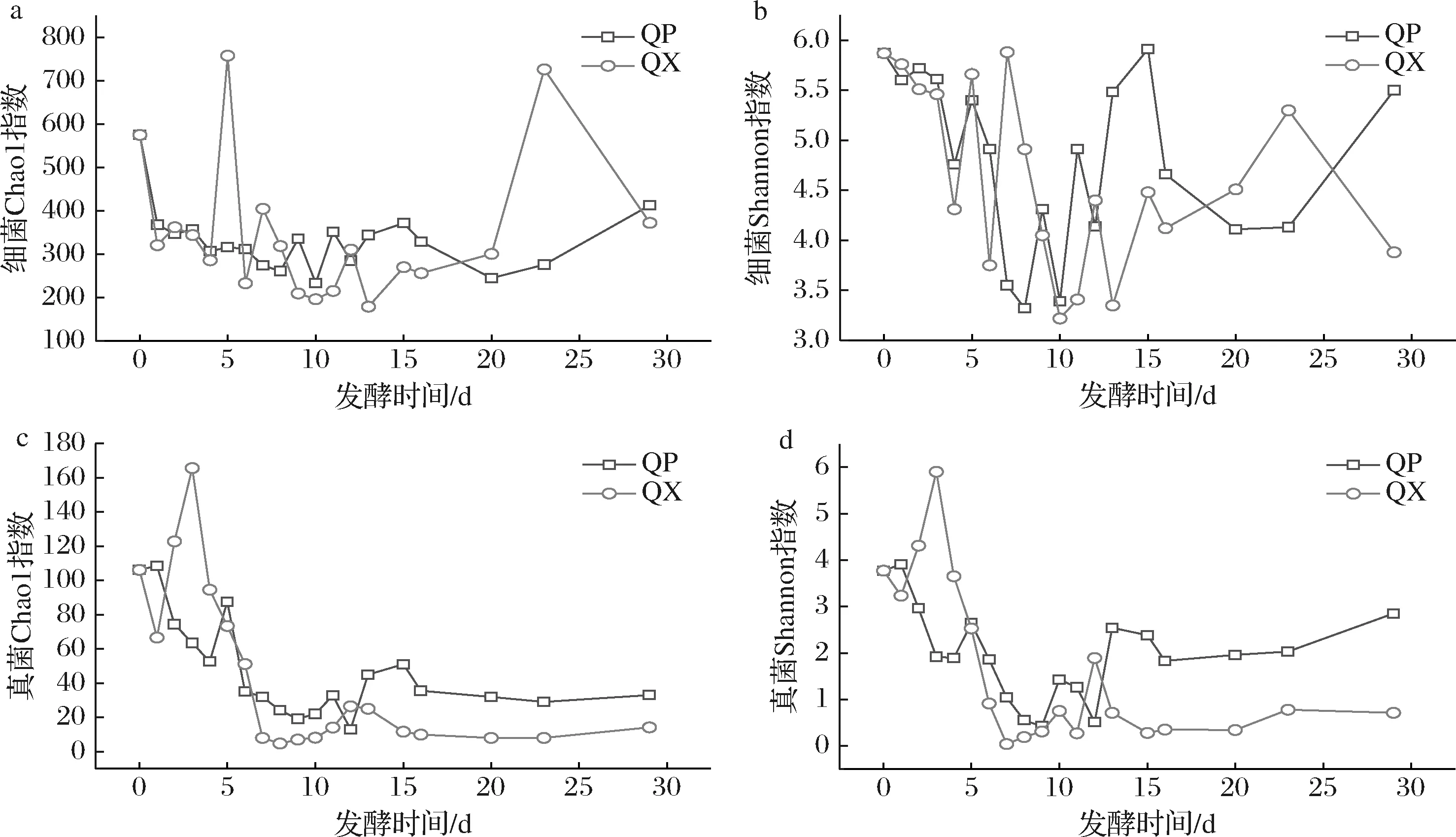
a,b-細菌多樣性指數(shù);c,d-真菌多樣性指數(shù)圖3 包包曲發(fā)酵過程中曲皮、曲心微生物群落α多樣性指數(shù)Fig.3 Microbial communities αdiversity index during Baobaoqu fermentation
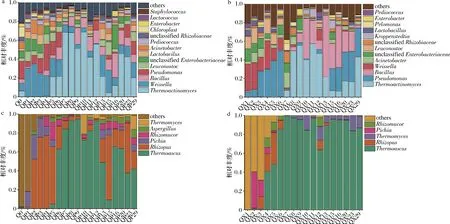
a-屬水平曲皮細菌群落結(jié)構(gòu);b-屬水平曲心細菌群落結(jié)構(gòu);c-屬水平曲皮真菌群落結(jié)構(gòu);d-屬水平曲心真菌群落結(jié)構(gòu)圖4 包包曲發(fā)酵過程中微生物群落演替規(guī)律Fig.4 Succession of microbial community during Baobaoqu fermentation
2.4 包包曲發(fā)酵過程中曲皮、曲心微生物群落的差異
2.4.1 包包曲發(fā)酵過程中曲皮、曲心樣品間微生物群落結(jié)構(gòu)的差異性和相似性
ANOSIM分析表明曲皮、曲心細菌群落(R=0.161,P=0.004)和真菌群落(R=0.131,P=0.01)存在顯著差異。基于bray-curtis距離對曲皮、曲心細菌和真菌微生物群落結(jié)構(gòu)的聚類分析,直觀展現(xiàn)了樣本間的差異性和相似性,如圖5-a和圖5-b所示。第1~6天和第7~29天包包曲細菌群落分別聚為2支(圖6-a),表明第1~6天和第7~29天大曲細菌群落結(jié)構(gòu)差異較大;而第1~5天和第6~29天大曲真菌群落結(jié)構(gòu)差異也較大(圖6-b)。從包包曲不同部位來看,曲皮、曲心細菌群落在第1~4天較為相似;而第7~12天的曲皮微生物群落結(jié)構(gòu)和第8~29天曲心微生物群落結(jié)構(gòu)較為相似。而QX7、QP13、QP15、QP29的細菌群落結(jié)構(gòu)和QP15真菌群落結(jié)構(gòu)與發(fā)酵前期樣品較為接近。曲皮、曲心微生物群落結(jié)構(gòu)聚類結(jié)果與理化因子變化具有相似的階段性,反映了理化因子與微生物群落間存在密切的相互影響關(guān)系,這種關(guān)系可能是包包曲微生物種群多樣性形成的重要原因。
2.4.2 包包曲發(fā)酵過程中曲皮、曲心微生物群落的差異微生物
利用LEfSe對包包曲發(fā)酵過程中曲皮、曲心間顯著差異微生物進行分析(閾值為4),結(jié)果如圖5-c和圖5-d所示。乳桿菌屬(Lactobacillus)、Weissella、Rhizopus、Rhizomucor、Pichia、Aspergillus的豐度在曲皮中更高,Thermoascus、Pseudomonadales、根瘤菌目(Rhizobiales)、酵母菌目(Saccharomycetales)、鎖擲酵母目(Sporidiobolales)的豐度在曲心中更高。曲心溫度更高可能導(dǎo)致了Thermoascus在曲心真菌群落中豐度更高、主導(dǎo)時間更長[21]。霉菌的好氧特性可能直接導(dǎo)致了其在曲皮中占優(yōu)勢。Lactobacillales的豐度變化趨勢與溫度一致,溫度可能是Lactobacillales在曲皮中豐度更高的重要原因。擲孢酵母屬(Sporobolomyces)的相對豐度極小,其相對豐度在曲皮中總是為0可能導(dǎo)致了Sporobolomyces在曲心中豐度顯著更大。而酵母菌易受環(huán)境因素影響,如低水分(<15%)和高溫(>50 ℃)[9],這可能導(dǎo)致了Pichia在曲皮中更豐富。
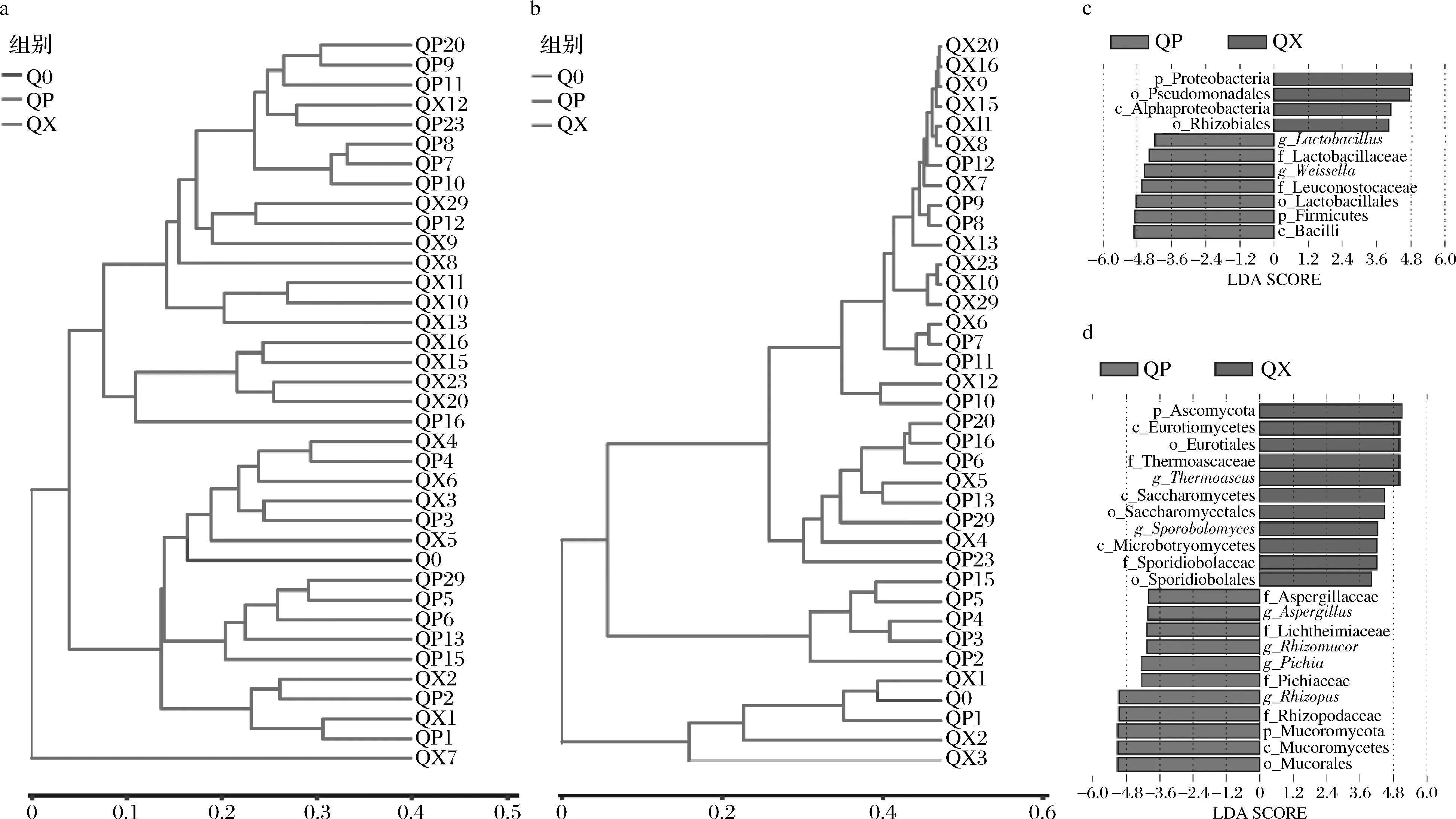
a-細菌群落結(jié)構(gòu)的聚類分析;b-真菌群落結(jié)構(gòu)的聚類分析;c-細菌顯著差異物種LDA得分值分布柱狀圖;d-真菌顯著差異物種LDA得分值分布柱狀圖圖5 包包曲發(fā)酵過程中曲皮、曲心微生物群落的差異Fig.5 Differences in microbial communities between QP and QX during Baobaoqu fermentation注:LDA-linear discriminant analysis
此外,包包曲出房時,曲皮、曲心優(yōu)勢微生物的豐度也具有顯著差異(P<0.05)。來自于大曲的Bacillus、Saccharomycetales、Aspergillus和Thermoascus等,是白酒主發(fā)酵前期的優(yōu)勢微生物,對白酒發(fā)酵過程的乙醇或風(fēng)味代謝有重要貢獻[20]。大曲中的Lactobacillales可直接參與白酒發(fā)酵[22],是白酒主發(fā)酵的主導(dǎo)菌群,還是酯類化合物積累的關(guān)鍵因素[23]。而大曲微生物代謝物質(zhì)也是白酒風(fēng)味的重要來源。因此,大曲微生物組成對白酒主發(fā)酵過程的微生物菌群結(jié)構(gòu)、豐度及微生物生長代謝都具有重要影響[24],認識和揭示包包曲多樣性及形成機制,對濃香型白酒主發(fā)酵過程具有重要意義。
2.5 包包曲發(fā)酵過程中曲皮、曲心微生物類群與理化因子的關(guān)系
利用RDA對包包曲發(fā)酵過程中曲皮、曲心細菌和真菌屬水平微生物群落與理化因子的關(guān)系進行分析,結(jié)果如圖6所示。曲皮、曲心細菌和真菌群落在第1~4天與水分、總酸呈正相關(guān),與溫度、pH值呈負相關(guān);而在第7~12天與溫度、pH值呈正相關(guān),與水分、總酸呈負相關(guān)。但第13~29天,曲心微生物群落主要受pH和溫度影響;而曲皮微生物群落在第15~20天仍主要受溫度和總酸含量的影響,在第23~29天主要受pH影響,曲皮、曲心樣本總是沿pH值的軸分布和聚集,表明pH值對曲皮、曲心微生物群落的影響較大。
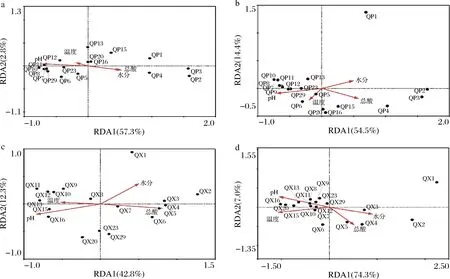
a-曲皮細菌;b-曲皮真菌;c-曲心細菌;d-曲心真菌圖6 包包曲發(fā)酵過程中微生物群落與理化因子的冗余分析Fig.6 Redundancy analysis of microbial communities and physicochemical factors during Baobaoqu fermentation
mantel test分析進一步表明,包包曲發(fā)酵過程中,pH值、總酸對曲皮、曲心的細菌和真菌組成具有顯著影響,溫度僅對真菌組成有顯著影響,而水分僅對曲皮微生物群落組成具有顯著影響(表1)。包包曲發(fā)酵過程中平均相對豐度前25的微生物屬與理化因子的Pearson相關(guān)性分析也表明pH值與優(yōu)勢微生物存在大量顯著正或負相關(guān)關(guān)系(圖7)。且pH值是影響微生物生長代謝的重要因素。Bacillus在曲心中與總酸呈負相關(guān),與2.3的結(jié)論一致,酸度可能是導(dǎo)致發(fā)酵前期Bacillus在曲皮中豐度更高的主要原因。曲皮、曲心的微環(huán)境差異是導(dǎo)致其微生物群落結(jié)構(gòu)差異的重要原因[4],如曲心中更高的溫度可能導(dǎo)致了嗜熱菌相對豐度在曲心中更具優(yōu)勢、更穩(wěn)定[25];而微生物群落代謝也會反過來影響環(huán)境因子,特別是酸度、溫度等理化特性[9]。pH值、總酸、溫度、水分是與曲皮、曲心微生物組成密切相關(guān)的重要影響因素。
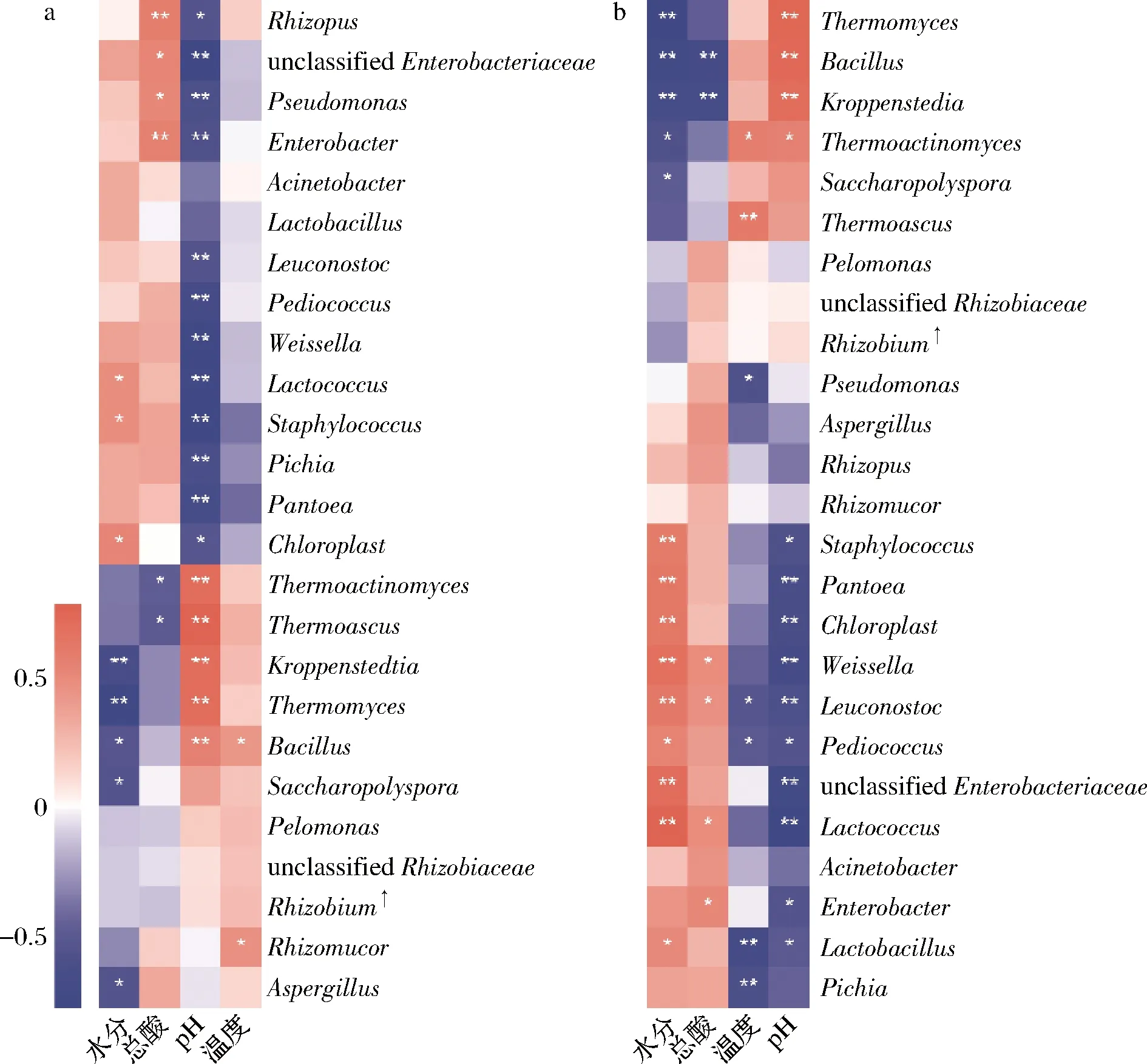
a-曲皮;b-曲心圖7 優(yōu)勢微生物與理化因子的Pearson相關(guān)性分析Fig.7 Pearson correlation analysis of dominant microorganisms and physicochemical factors注:*表示顯著相關(guān)(P <0.05);**表示極顯著相關(guān)(P<0.01);Rhizobium :Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium
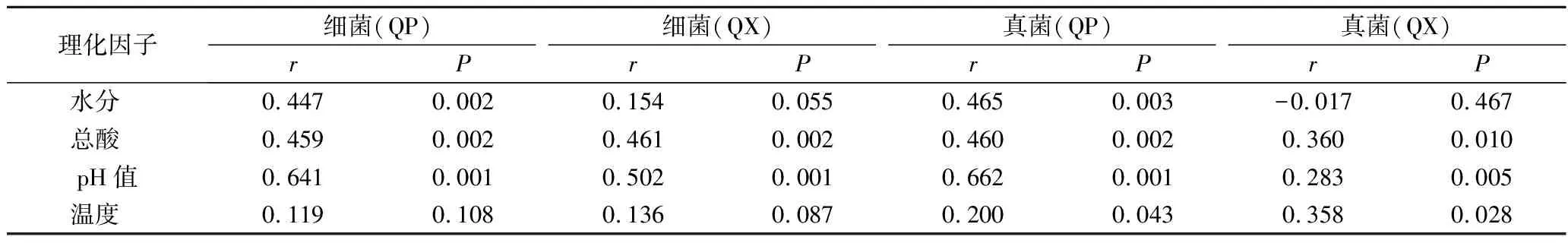
表1 包包曲發(fā)酵過程中細菌和真菌群落與理化因子的mantel test分析Table 1 Mantel test analysis of bacteria and fungi communities and physicochemical factors during Baobaoqu fermentation
3 結(jié)論與討論
通過研究濃香型包包曲發(fā)酵過程中曲皮、曲心理化特性和微生物群落組成,討論理化因子對微生物群落的影響,得出以下結(jié)論:曲皮、曲心的理化特性從發(fā)酵第2天開始出現(xiàn)差異,并且曲皮pH值高于曲心,水分、溫度和總酸低于曲心;高通量測序結(jié)果表明曲皮、曲心優(yōu)勢微生物類群相似,為Lactobacillales、Pseudomonadales、腸桿菌目(Enterobacteriales)、Bacillales、Eurotiales、Mucorales、Saccharomycetales,但曲皮、曲心優(yōu)勢微生物在發(fā)酵過程中的分布和豐度具有明顯區(qū)別;ANOSIM分析表明曲皮、曲心微生物群落存在顯著差異;LEfSe分析得出Lactobacillus、Weissella、Rhizopus、Rhizomucor、Pichia、Aspergillus的豐度在曲皮中顯著更高,Thermoascus、Pseudomonadales、Rhizobiales、Saccharomycetales、Sporidiobolales的豐度在曲心中顯著更高;RDA、mantel test分析及Pearson相關(guān)性分析共同表明,pH值、總酸、溫度、水分是與曲皮、曲心微生物組成密切相關(guān)的重要影響因素。本研究揭示了包包曲發(fā)酵過程中曲皮、曲心微生物群落多樣性差異及與大曲理化因子的相關(guān)性,有助于解析包包曲微生物種群多樣性的形成機制。
研究結(jié)果發(fā)現(xiàn)包包曲發(fā)酵過程中曲皮、曲心的理化性質(zhì)、微生物群落結(jié)構(gòu)、理化因子與微生物類群的關(guān)系間的差異都存在較明顯的階段性,這些階段間存在一定規(guī)律但尚不明晰。進一步通過引入其他與大曲微生物類群生長相關(guān)的因素,如氧氣濃度或加大樣本量等方式,可能有助于包包曲發(fā)酵過程中微生物群落變化規(guī)律的探究。而各環(huán)境因子是協(xié)調(diào)影響微生物群落的,微生物群落代謝也會反過來影響環(huán)境因子,量化的復(fù)合因素對微生物群落的研究或許能夠進一步解析包包曲微生物種群變化規(guī)律。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
