pH 急性脅迫對棘胸蛙蝌蚪行為、尾部皮膚色差及耗氧率、排氨率和窒息點的影響
2021-10-27 08:28:06劉哲宇申屠琰朱衛東王志錚
浙江海洋大學學報(自然科學版) 2021年3期
劉哲宇,申屠琰,楊 程,朱衛東,王志錚
(1.浙江海洋大學水產學院,舟山 316022;2.余姚市水產技術推廣中心,余姚 315400)
就養殖生產而言,養殖生物實質上就是一臺以維系存活并促進其生長為主要運動方式的“產肉機器”,其對養殖環境的感應與適應在宏觀水平上常通過對其行為、體色、呼吸、排泄等的調節來反映或體現。pH既是水生動物最敏感的環境因子之一[1],也是評價養殖水環境優劣的必檢項[2]。因水生生物的光合作用和呼吸活動,池塘水體pH 常波動于6.6~10.2 之間[3-4],故池養水產動物常處于pH 脅迫狀態。研究表明,養殖水體易變的pH,不僅可直接影響水生動物的載氧能力并麻痹其呼吸中樞,而且還可通過影響水中物質的存在形式和遷移過程間接毒害水生動物,從而導致機體出現多種損傷癥狀[5-6]。因此,探究并確定目標養殖對象適宜的水環境pH 范圍一直是水產養殖動物生理生態學研究的常規命題之一。棘胸蛙Quasipaa spinosa 素有“百蛙之王”之美譽,是一種我國南方丘陵山區極具養殖開發潛力的特有經濟蛙種。為切實弄清棘胸蛙蝌蚪期養殖水體的適宜pH 范圍,本文作者在探究該蛙蝌蚪尾部皮膚和肝臟對養殖水體pH 抗氧化應答特征的同時[7],較系統開展了pH 急性脅迫對該蛙蝌蚪行為、尾部皮膚色差及耗氧率、排氨率和窒息點的影響研究,旨為該蛙蝌蚪的平原化安全高效培育提供科學依據。
1 材料與方法
1.1 材料
本研究所用棘胸蛙蝌蚪的來源、實驗前馴養處理和實驗個體的選擇要求與規格,以及實驗水源和實驗用水的pH 配置方法均完全同劉哲宇等[7]。
1.2 方法
1.2.1 蝌蚪行為及尾部皮膚色差值的觀測
以直徑32 cm的塑料盆為實驗容器單元(實驗實際容積為5 L),以2 000 ind·m-3為蝌蚪實驗密度,以96 h 為實驗周期,設置5.5、6.5、7、7.5、8.5 和9.5 等6 個pH 處理梯度(每一梯度設3 個重復),完全按朱衛東等[8]的方法觀察并統計蝌蚪水層分布、集群狀況及個體移動率,并于實驗終了時刻測定各pH 處理梯度組實驗蝌蚪(每一梯度各任測5 ind)尾部皮膚黑、白斑的表觀色澤。
1.2.2 蝌蚪耗氧率、排氨率的測定
以直徑32 cm的塑料盆為實驗容器單元(實驗實際容積為5 L),以2 000 ind·m-3為蝌蚪實驗密度,以24 h為實驗周期,設置5.5、6.5、7、7.5、8.5 和9.5 等6 個pH 處理梯度(每一梯度設3 個重復),按楊程等[9]的方法,以凌晨2:00 為實驗起始時點,測算各pH 處理梯度組的OR (耗氧率)、NR (排氨率) 和O:N (氧氮比)。
1.2.3 蝌蚪窒息點的測定
以容量為500 mL的容量瓶(實驗實際容積為50 mL)為呼吸室,設置5.5、6.5、7、7.5、8.5 和9.5 等6 個pH 處理梯度(每一梯度設3 個重復,每個重復均放入實驗蝌蚪4 ind),將蝌蚪置于對應實驗pH 處理組的呼吸室后即刻開始實驗,并按楊程等[9]的方法測定各pH 處理梯度組的窒息點。
1.3 數據處理使用
2 結果
2.1 行為
觀察發現,棘胸蛙蝌蚪的活動區域、需氧壓力和運動頻率均與其所處水環境pH 存在密切關聯,主要表現為:(1)實驗期間,pH 5.5~6.5 實驗組的蝌蚪散游于水體近表面,pH 7.0 和9.5 實驗組的蝌蚪散布于整個水體,均可見浮頭現象,而pH 7.5~8.5 實驗組蝌蚪則集群游動于整個水體,且未見有浮頭現象;(2) pH 5.5~7.0實驗組蝌蚪的移動頻率明顯高于其它實驗組。綜上可知,就分布格局、集群行為、需氧壓力和移動頻率觀,棘胸蛙蝌蚪偏好弱堿性環境,其適宜pH 范圍為7.5~8.5。
2.2 尾部皮膚色差
由表1 可見,盡管棘胸蛙蝌蚪尾部皮膚黑、白斑的表觀色澤A、B 值均無組間差異(P>0.05),但其L 值卻與所處水環境pH 存在密切關聯,主要表現為:(1)黑、白斑L 值均以pH 5.5 和6.5 兩實驗組為最大(P<0.05),pH 8.5 實驗組為最小(P<0.05);(2)pH 7.5 和9.5 兩實驗組的黑、白斑L 值均無組間差異(P<0.05),且均顯著小于pH 7.0 實驗組(P<0.05)。綜上可知,用于表征尾部皮膚亮度的L 值呈酸性環境>中性環境>堿性環境(P<0.05),對pH 環境具指示作用,以pH 8.5 為最低(P<0.05)。
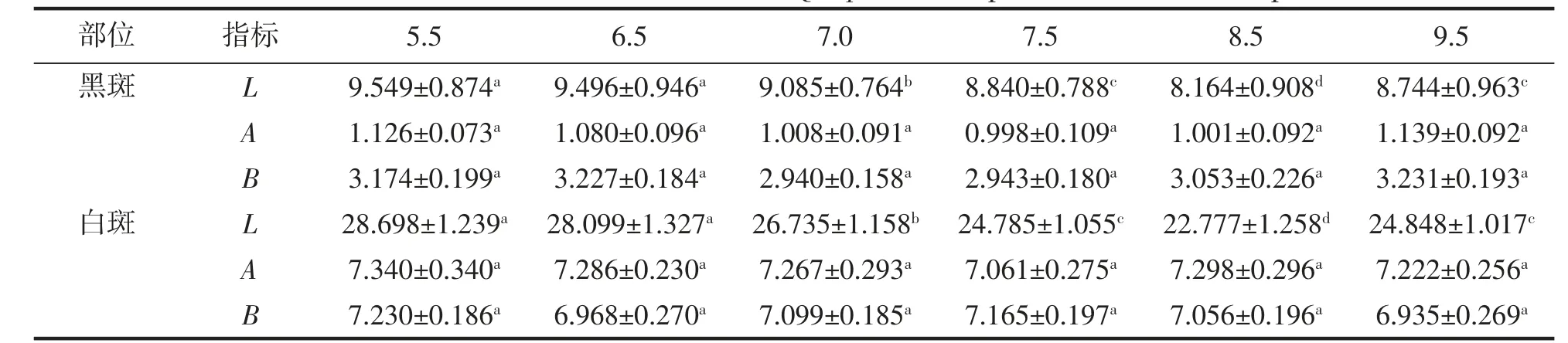
表1 不同pH 條件下棘胸蛙蝌蚪尾部表觀色澤的L、A、B 值Tab.1 The Hunter L,A,B values of Q.spinosa tadpole tail in different pH
2.2 OR(耗氧率)
由圖1 可見,pH 對蝌蚪OR的影響主要表現為:(1)從時段OR 看,堿性環境下的OR 較酸性和中性環境高。其中,pH 5.5 與6.5 實驗組間存在顯著差異的時段僅為06:00-10:00,兩者間的時段相似性達83.3%,而其與其它實驗組的相似性則均為0;pH 6.5 與7.0 實驗組間的時段相似性為100%,兩者與pH 7.5 實驗組間的相似性分別為16.7%和33.3%,與pH 8.5 實驗組間的相似性分別為33.3%和50%,而與pH 9.5 實驗組間的相似性則分別為50%和67.7%;pH 7.5 與8.5 實驗組間以及pH 8.5 與9.5 實驗組間的時段相似性均為100%,而pH 7.5 與9.5 實驗組間的時段相似性則降為83.3%,表明本研究所涉堿性環境與酸性和中性環境間時段OR 差異最大的為pH 7.5 實驗組,而與pH 7.5 實驗組時段OR 最為接近的則為pH 8.5實驗組;(2)從晝均、夜均和日均OR 看,各實驗組均呈夜均>晝均≈日均(P<0.05),與所涉酸性和中性范圍內類似,堿性范圍內的組間夜均、晝均和日均OR 相似性也均為100%,但其中僅pH 7.5 和8.5 兩實驗組的夜均、晝均和日均OR 均顯著高于酸性和中性環境(P<0.05),表明盡管本研究所設pH 范圍內蝌蚪的耗氧晝夜節律尚未發生根本性改變,不同pH 性質范圍內的組間夜均、晝均和日均OR 均較穩定,但仍改變不了弱堿性環境下蝌蚪夜均、晝均和日均OR 均明顯高企的事實。綜上可知,棘胸蛙蝌蚪的最適pH 為7.5,適宜范圍為7.5~8.5,當所處水環境pH 偏離7.5~8.5 范圍時,其時段及夜均、晝均和日均OR 均會出現不同程度的下降。
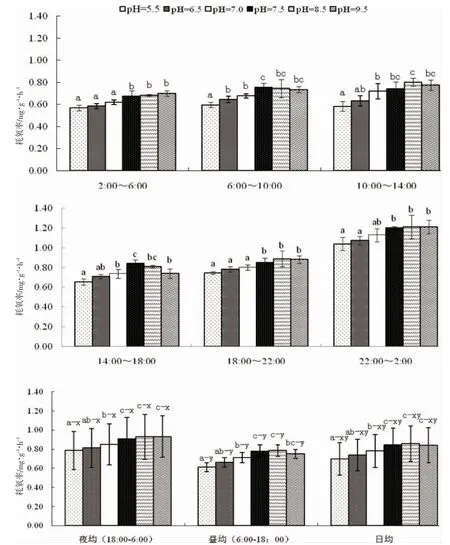
圖1 pH 對棘胸蛙蝌蚪耗氧率的影響Fig.1 Effects of pH on oxygen consumption rate of Q.spinosa tadpole
2.3 NR (排氨率)
由圖2 可見,pH 對蝌蚪NR的影響主要表現為:(1)從時段NR 看,各實驗組間的時段NR 均具較高的相似性。其中,pH 5.5 與6.5 實驗組間的時段NR 相似性為100%,兩者與pH 7.0、7.5、9.5 實驗組間的相似性均為88.3%,與pH 8.5 實驗組間的相似性分別為33.3%和66.7%;pH 7.0、7.5 和9.5 實驗組間的時段NR相似性均為100%,三者與pH 8.5 實驗組間的相似性分別為66.7%、88.3%和88.3%,表明與酸性和中性環境間時段NR 差異最大的組別為pH 8.5 實驗組;(2)從晝均、夜均和日均NR 看,各實驗組均呈夜均>晝均≈日均(P<0.05),與所涉酸性和中性范圍內類似,中性和堿性范圍內的組間夜均、晝均和日均NR 相似性也均為100%,其中與酸性環境夜均、晝均和日均NR 差異最大的組別為pH 8.5 實驗組,表明本研究所設pH范圍內蝌蚪的排氨晝夜節律尚未發生根本性改變,不同pH 性質范圍內的組間夜均、晝均和日均NR 均較穩定,但仍改變不了pH 由堿性轉為酸性時夜均、晝均和日均NR 均出現一定程度下降的事實。綜上可知,棘胸蛙蝌蚪的最適pH 為8.5。
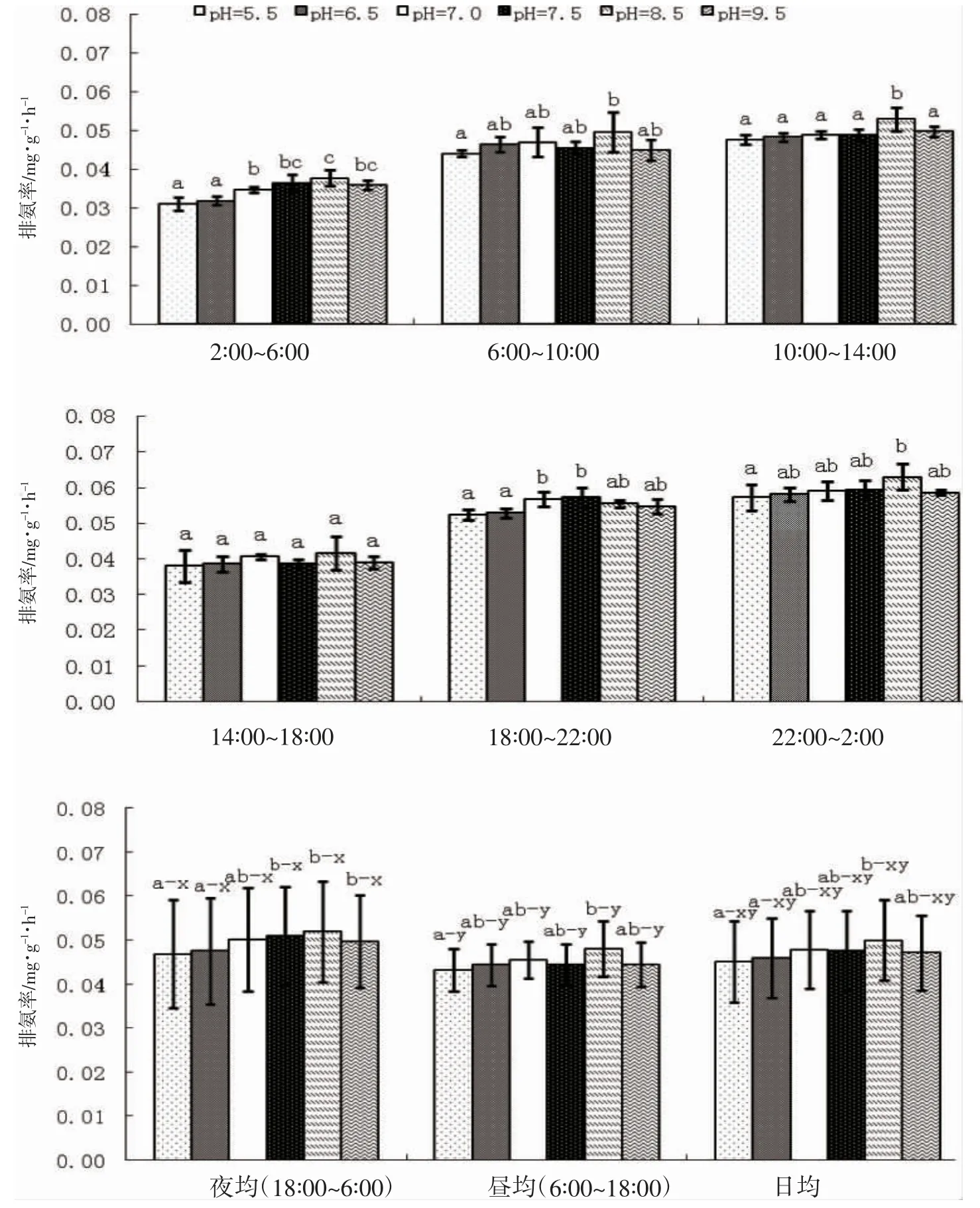
圖2 pH 對棘胸蛙蝌蚪排氨率的影響Fig.2 Effects of pH on ammonia excretion rate of Q.spinosa tadpole
2.4 O:N (氧氮比)
由圖3 可見,pH 對蝌蚪O:N 值的影響主要表現為:(1)諸實驗組中,夜均、晝均和日均O:N 值除pH 5.5實驗組均低于16 外,其余均波動于16.00~20.07 之間;(2)晝均O:N 值以pH 5.5 實驗組最小(P<0.05),pH 7.5 實驗組最大(P<0.05),其余實驗組間均無顯著差異(P>0.05);(3)夜均O:N 值隨pH 上升呈階梯式穩步上升趨勢,階梯式上升的臨界點分別為pH 6.5 和7.5(P<0.05);(4)日均O:N 以pH 7.5 和9.5 兩實驗組最大(P<0.05),pH 5.5 實驗組最小(P<0.05),其余實驗組間均無顯著差異(P>0.05);(5)除pH 7.5、8.5 兩實驗組的O:N值呈晝均>夜均(P<0.05)外,其余實驗組間均呈夜均≈晝均(P>0.05)。綜上可知,諸實驗組中晝均、夜均和日均O:N 值均以pH 5.5 實驗組為最小(P<0.05),僅pH 7.5 實驗組的晝均、夜均和日均O:N 值均不低于其它實驗組,唯pH 7.5~8.5 范圍內的O:N 值具晝夜差異(P<0.05)。
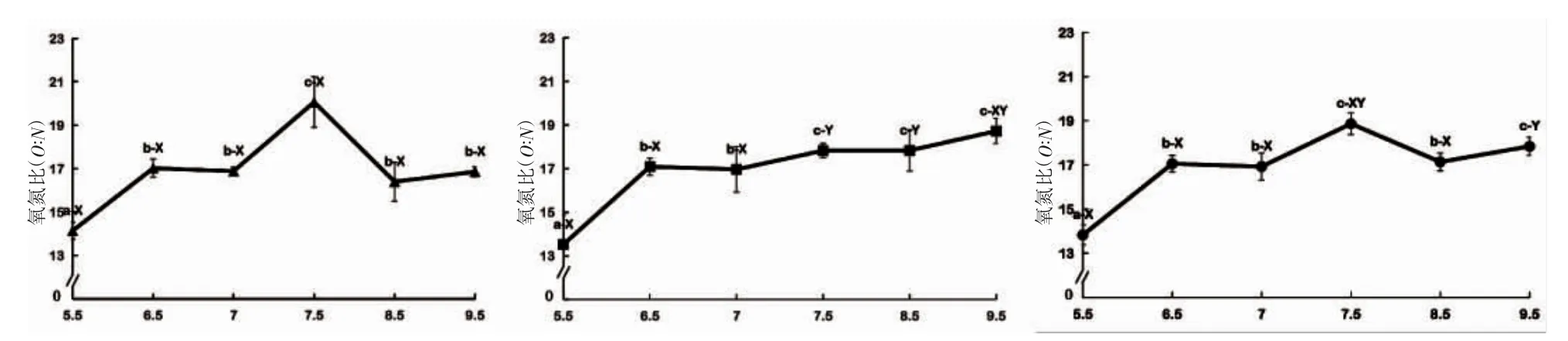
圖3 pH 對棘胸蛙蝌蚪氧氮比的影響Fig.3 Effects of pH on ammonia excretion rate of Q.spinosa tadpole
2.5 窒息點
由圖4 可見,蝌蚪窒息點的水中氧含量隨pH 上升呈先降后升走勢,并于pH 7.5 時到達谷底(P<0.05);其中,與pH 6.5 實驗組無顯著差異的僅為pH 9.5 實驗組(P>0.05),而與pH 7.0 實驗組無顯著差異的僅為pH 9.5 實驗組(P>0.05)。由此可知,蝌蚪所處水環境pH 偏離7.5 越遠,其窒息點水中氧含量也就越高(P<0.05)。
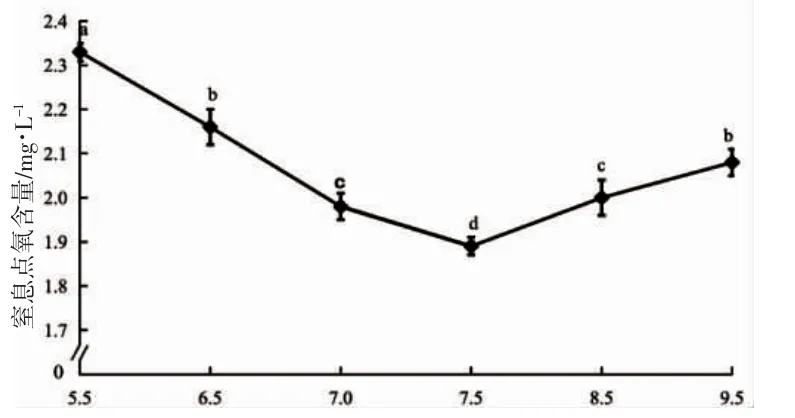
圖4 pH 對棘胸蛙蝌蚪窒息點的影響Fig.4 Effects of pH on the suffocation point of Q.spinosa tadpole
3 討論
3.1 棘胸蛙蝌蚪對pH 急性脅迫的應答特征
蛙類是重要的環境指示生物,對水質變化極為敏感[10]。據報道,薄且布有大量毛細血管的皮膚為無尾類蝌蚪的主要呼吸器官并兼具滲透壓調節等多種功能[11-12],處于低氧水域中的蝌蚪皮膚往往有變薄和毛細血管增多的傾向[13-14]。這與棘胸蛙蝌蚪尾部皮膚L 值隨水溫升高而顯著增加(P<0.05)的情形[8]相吻。故,可將尾部皮膚L 值作為指示棘胸蛙蝌蚪供氧壓力的顯性指標,即供氧壓力隨尾部皮膚L 值的增大而增加,反之亦然。觀察發現,仿生態條件下飼養的棘胸蛙蝌蚪具聚群活動的習性[15]。無疑,本研究pH 5.5~7.0范圍內蝌蚪所表露的固有集群行為消失、移動頻率加大、運動無序性和浮頭個體明顯增加的現象,與表1所示尾部皮膚L 值顯著大于pH 7.5~9.5 (P<0.05),圖1 所示夜均和日均OR 均顯著低于pH 7.5~9.5 (P<0.05),以及圖2 所示pH 由堿性轉為酸性時夜均、晝均和日均NR 均出現一定程度下降的情形相對應,表明pH 5.5~7.0 范圍內高企的皮膚供氧壓力已對其機體代謝產生負面影響,尤以對皮膚具高腐蝕性的酸性環境為甚。
研究發現,水生動物OR 和NR 呈晝低高夜的變化特征與其晝伏夜出的生活習性密切相關[16-17]。由圖1、圖2 所示,各實驗組的OR 和NR 均呈夜均>晝均(P<0.05),與其固有生活習性相符[8-9,18-19],表明它們的耗氧和排氨晝夜節律均未發生根本性改變。但各實驗組間在O:N(圖3)和窒息點(圖4)的應答機制上卻不盡相同,主要表現為:(1)O:N 值系反映機體代謝底物利用狀況的重要生理指標,當取值3~16 時代謝底物完全為蛋白質,50~60 時為等量的脂肪和蛋白質,無窮大時則完全為脂肪和糖類[20]。由圖3 可見,除pH 5.5 實驗組的代謝底物完全為蛋白質外,其余各實驗組均仍保持其常態下以蛋白質為主、脂肪為輔的代謝底物利用模式[9.18],諸實驗組中僅pH 7.5 實驗組的晝均、夜均和日均O:N 值均不低于其它實驗組,唯pH 7.5~8.5 范圍內的O:N 值具晝夜差異呈晝均>夜均(P<0.05),表明棘胸蛙蝌蚪機體代謝過程中脂肪動員能力的強弱以及是否具有晝夜差異,系應答其pH 是否處于適宜范圍的重要依據,即pH 環境越適宜,固有生活習性就越穩定,機體脂肪動員能力就越強,體內產氨量就越小,反之亦然,由此也進一步印證了牛春格等[18]所述該蛙蝌蚪具通過增強脂肪動員以抑制蛋白質水解和氨基酸代謝水平,從而主動減少體內產氨量的耐氨策略的可靠性;(2)窒息點系評價水生動物耐低氧能力的重要指標。由圖4 可見,pH 偏離7.5 越遠,窒息點水中氧含量就越高,表明棘胸蛙蝌蚪血液載氧能力就越弱,皮膚供氧壓力也就越大,故窒息點亦可作為該蛙蝌蚪應答其pH 是否處于適宜范圍的重要依據。綜上,上述有關O:N 和窒息點對pH 急性脅迫的應答特征,既與pH 偏離7.5 時,棘胸蛙蝌蚪尾部皮膚ATP 酶和肝臟GSH 酶活力均呈顯著上升(P<0.05),尾部皮膚與肝臟的SOD 酶和CAT 酶活力均呈顯著下降(P<0.05)的結果相符[7],也揭示了皮膚供氧壓力與脂肪動員能力對pH 急性脅迫的互助應答效應,即皮膚供氧壓力越小,脂肪動員能力也就越強,固有生活習性也就越穩定。
至于,pH 9.5 實驗組尾部皮膚L 值顯著大于pH 6.5 實驗組(P<0.05),而與pH 7.5 實驗組無顯著差異(P>0.05)(表1),窒息點水中氧含量與pH 6.5 實驗組無顯著差異(P>0.05),而顯著高于pH 7.5 實驗組(P<0.05)(圖4),且其固有集群行為消失、運動無序性增加并伴有浮頭現象的原因,一方面可能與該實驗組蝌蚪皮膚的受腐蝕程度弱于pH 6.5 實驗組有關,另一方面應與其喪失O:N 值晝夜節律導致皮膚供氧壓力有所增加有關。由此,進一步表明O:N 值和窒息點在表征pH 適宜范圍上較尾部皮膚L 值更具準確性。無疑,pH 7.5 較pH 8.5 更適宜棘胸蛙蝌蚪的原因亦同此理,盡管pH 7.5 實驗組尾部皮膚的L 值顯著大于pH 8.5實驗組(P<0.05)(表1)。
3.2 pH 急性脅迫下棘胸蛙蝌蚪的內穩態保持機制
通過內穩態保持機制維系或擴大生態幅,是動物積極響應外界環境脅迫以有效消除或降低由此引發的負面影響的重要手段,盡管該機制存在較明顯的局限性。行為調節和生理調節作為動物內穩態保持機制的主要途徑,兩者在響應外界環境變化并共同保持動物內穩態上往往會形成“表里互通”的代償效應。就本研究而言,pH 急性脅迫下維系OR 和NR 晝夜生理節律是棘胸蛙蝌蚪通過行為調節和生理調節保持內穩態的前提(圖1、圖2),而維系皮膚供氧水平和機體脂肪動員能力間的適配性則是pH 急性脅迫下棘胸蛙蝌蚪內穩態保持機制的內核,這與水溫[8-9]、氨氮[18,21]急性脅迫對棘胸蛙蝌蚪行為和生理影響效應的研究結果相一致。
如上所述,盡管水溫、氨氮和pH 間在理化性質上存在較明顯的差異,但它們在棘胸蛙蝌蚪適宜范圍或最適點的形成機制上卻具明顯的相似性,即:(1)隨水溫上升,當蝌蚪夜均、晝均和日均O:N 值均抵達峰值(P<0.05),窒息點水中含氧量脫離穩定階段并顯著增加(P<0.05),各測定時段尾部皮膚ATP 酶活力持續處于峰值(P<0.05)時,表明該水溫(24.5 ℃)已偏離最適水溫(23 ℃)[8-9]。也就是說,隨水溫繼續增加,即使OR和NR 晝夜節律不發生改變,但其皮膚供氧能力卻已無法有效保障機體進一步動員脂肪的需求;(2)隨氨氮質量濃度增加,當蝌蚪各測定時段尾部皮膚CAT 酶和肝臟GSH 酶、CAT 酶活力均持續處于峰值(P<0.05),晝均O:N 值處于峰值區氨氮質量濃度上限時,表明此時已處于蝌蚪維系低毒興奮效應和耐氨策略的臨界,該氨氮質量濃度(14.70 mg·L-1)為其安全質量濃度的上限[18,21]。也就是說,隨氨氮質量濃度的繼續增加,即使其OR 和NR 晝夜生理節律尚未發生根本性改變,但其皮膚供氧能力卻已難以挽回脂肪動員能力即將顯著回落的結果;(3)由表1、圖3 和圖4 可見,當窒息點水中含氧量抵達谷底(P<0.05),晝均O:N 值處于峰值(P<0.05),各測定時段尾部皮膚ATP 酶和肝臟GSH 酶活力均持續處于谷底(P<0.05),尾部皮膚和肝臟的SOD 酶、CAT 酶活力均處于峰值(P<0.05)時,該pH 為蝌蚪的最適pH(7.5),即當環境pH 偏離該值時,即使其OR 和NR 晝夜生理節律未發生根本性改變,也難以改變因皮膚供氧壓力的持續增加而導致其機體脂肪動員能力顯著下降的事實。綜上可知,就以蛋白質為主和脂肪為輔的代謝底物利用模式的棘胸蛙蝌蚪而言,維系皮膚供氧水平和機體脂肪動員能力間的適配性既是保障其機體生理活動正常運行的重要基礎,更是其啟動內穩態保持機制的源動力。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
發明與創新(2016年38期)2016-08-22 03:02:52
