大腸桿菌乙酸弱化對1,2,4-丁三醇合成的影響
2021-10-28 02:30:38狄瑩瑩王昕鈺馮奧陸信曜宗紅諸葛斌
食品與發(fā)酵工業(yè) 2021年19期
狄瑩瑩,王昕鈺,馮奧,陸信曜,宗紅,諸葛斌*
1(糖化學(xué)與生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室(江南大學(xué)),江蘇 無錫,214122) 2(江南大學(xué),工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,工業(yè)微生物研究中心,江蘇 無錫,214122)
1,2,4-丁三醇(1,2,4-butantriol,BT)是三羥基四碳多元醇,易溶于醇類及水,無色無味的油狀液體[1]。BT是重要的藥物合成中間體,可用于合成腫瘤和皮膚病藥物等;也可用于減少煙草中的有害物質(zhì)[2]。BT的三硝酸酯(1,2,4-butanetriol trinitrate,BTTN)是重要的增塑劑,具有比硝化甘油更好的熱穩(wěn)定性、可塑性和低敏感性,是導(dǎo)彈等軍事武器的重要推進(jìn)器成分等[3]。
近年來,在大腸桿菌(Escherichiacoli)中以木糖為底物開展了大量BT生物合成途徑研究。BT的生物合成最后一步需要NAD(P)H參與醇脫氫[3-4],同時(shí),葡萄糖和木糖2種底物的轉(zhuǎn)運(yùn)需要消耗ATP[5],這兩者可以通過TCA循環(huán)獲得,但是代謝會產(chǎn)生低級酸等副產(chǎn)物[6],即代謝溢流效應(yīng)。乙酸會影響細(xì)菌胞內(nèi)pH值、滲透壓和氨基酸的合成[7],從而降低生物量、ATP和還原力,同時(shí)外源基因表達(dá)受到嚴(yán)重影響[8]。而且乙酸通過乙酰輔酶A合成酶途徑(ACS,由acs基因編碼)轉(zhuǎn)化為乙酰輔酶A時(shí)會消耗大量ATP[9]。
目前,BT的代謝合成研究主要集中在弱化BT途徑直接副產(chǎn)物合成[4]、高活性脫羧酶篩選[10]以及建立無細(xì)胞體系[11-12]等,本論文敲除宿主細(xì)胞E.coli的乙酸合成關(guān)鍵基因磷酸轉(zhuǎn)乙酰化酶(pta)和丙酮酸氧化酶(poxB)弱化大腸桿菌乙酸途徑,減少乙酸溢流,增加丙酮酸和乙酰輔酶A節(jié)點(diǎn)流向草酰乙酸等下游路徑的碳流方法,探索提高1,2,4-丁三醇產(chǎn)量,獲得了較好的結(jié)果,為以E.coli為底盤細(xì)胞的類似化合物的合成生物學(xué)改造提供借鑒。
1 材料與方法
1.1 試劑與儀器
卡那霉素、壯觀霉素、異丙基-β-D-硫代半乳糖苷(IPTG)、阿拉伯糖,生工生物工程(上海)股份有限公司;蛋白胨、酵母提取物,英國Oxoid公司;DNA凝膠回收試劑盒、質(zhì)粒提取試劑盒,上海康朗生物科技有限公司;高保真酶、限制性內(nèi)切酶,大連寶生物工程有限公司(TaKaRa);一步法克隆試劑盒,南京諾唯贊生物科技有限公司;1,2,4-丁三醇,上海阿拉丁化學(xué)試劑有限公司;其他試劑均為國產(chǎn)分析純。
PCR儀、電轉(zhuǎn)儀,Eppendorf公司;液相色譜柱:Aminex HPX-87H column(300 mm×7.8 mm;9 μm),Bio-Rad公司;BIO-TECH 5L 發(fā)酵罐,上海保興生物設(shè)備工程有限公司;引物由上海亦欣生物科技有限公司合成。
1.2 菌株與質(zhì)粒
本研究使用的菌株和質(zhì)粒參見表1。
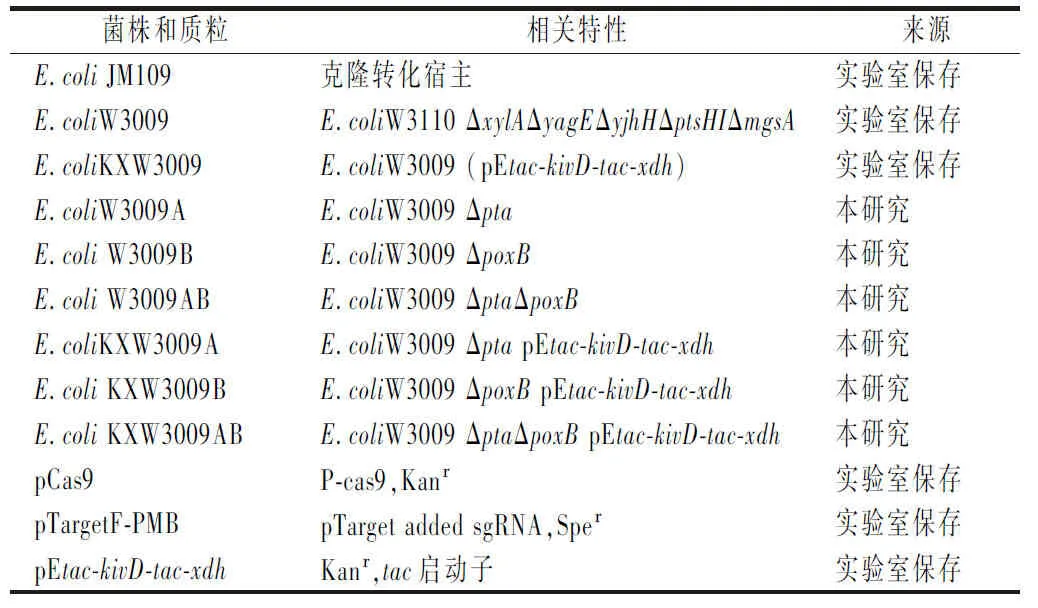
表1 本研究使用的菌株和質(zhì)粒Table 1 Strains and plasmids used in this study
1.3 引物
本研究使用的引物參見表2。

表2 本研究使用引物Table 2 Primers used in this study
1.4 培養(yǎng)基與培養(yǎng)方法
種子培養(yǎng)基(LB培養(yǎng)基)(g/L):NaCl 10.0,蛋白胨10.0,酵母提取物5.0。
發(fā)酵培養(yǎng)基(g/L):NaCl 15.0,蛋白胨15.0,酵母提取物7.5,CaCO310.0,木糖30.0,葡萄糖10.0。
5L發(fā)酵罐培養(yǎng)基(g/L):NaCl 15.0,蛋白胨15.0,酵母提取物7.5,葡萄糖、木糖濃度參照以下各圖圖例。
種子培養(yǎng):1%接種量(體積比),37 ℃、150 r/min培養(yǎng)12 h。
發(fā)酵培養(yǎng):250 mL三角瓶中裝液量為50 mL,1%接種量(體積比)。37 ℃、200 r/min培養(yǎng)至OD600 nm為0.6~0.8時(shí),加入終濃度為0.5 mmol/L IPTG進(jìn)行誘導(dǎo),同時(shí)加入木糖、葡萄糖,繼續(xù)培養(yǎng)至48 h。所有發(fā)酵設(shè)置3個(gè)平行實(shí)驗(yàn)。
上罐發(fā)酵培養(yǎng):5 L發(fā)酵罐裝液量為2.5 L。接種量為10%(體積比),培養(yǎng)溫度維持在37 ℃,轉(zhuǎn)速為400 r/min,通氣量 1.5 vvm,pH控制在7.0。所有發(fā)酵設(shè)置3個(gè)平行實(shí)驗(yàn)。
1.5 敲除菌株構(gòu)建
根據(jù)參考文獻(xiàn)[14]中的CRISPR/Cas9技術(shù)對E.coliW3009進(jìn)行基因敲除。在NCBI上查找E.coliW3110中pta和poxB基因序列設(shè)計(jì)引物(表2),分別構(gòu)建含有sgRNA的pTargetF-PMB[15](引物為Sg-PTA-F/Sg-PTA-R,Sg-PoxB-F/Sg-PoxB-R)和目的基因上、下游各500 bp的融合片段[14](引物為PTA-F1/PTA-R1,PTA-F2/PTA-R2;PoxB-F1/PoxB-R1,PoxB-F2/PoxB-R2)。將pCas9質(zhì)粒電轉(zhuǎn)入E.coliW3009中。加入終濃度為50 mmol/L的阿拉伯糖誘導(dǎo)pCas9質(zhì)粒的表達(dá),將pTargetF-PMB和融合的片段電轉(zhuǎn)入含有質(zhì)粒pCas9的E.coliW3009,涂布含有50 mg/L Kan 和100 mg/L Spe抗性的LB平板,30 ℃過夜培養(yǎng),然后通過菌落PCR(引物PTA-F-1/PTA-R-1,PoxB-F-1/PoxB-R-1)篩選陽性轉(zhuǎn)化子。敲除成功的菌株在對數(shù)生長期時(shí)加入終濃度為0.5 mmol/L IPTG誘導(dǎo)消除pTargetF-pMB質(zhì)粒;42 ℃連續(xù)傳代消除pCas9質(zhì)粒,最終獲得不帶抗性的敲除菌株。
1.6 檢測方法
發(fā)酵液中葡萄糖、木糖、BT和乙酸等代謝產(chǎn)物用HPLC示差法檢測,木糖酸在紫外下檢測。流動相為5.0 mmol/L的H2SO4,流速為0.6 mL/min,色譜柱為Aminex HPX-87H,柱溫60 ℃,使用示差和紫外檢測器進(jìn)行檢測,進(jìn)樣量為10 μL。OD600 nm利用分光光度計(jì)測定發(fā)酵液在600 nm下的吸光度。
2 結(jié)果與分析
2.1 弱化乙酸途徑對底盤細(xì)胞生長的影響
如圖1所示,敲除poxB基因,OD600 nm值達(dá)到13.8,提高了26.6%。其原因是弱化了丙酮酸流向乙酸的碳流,促進(jìn)TCA循環(huán)[16],為細(xì)胞生長提供更多能量。相反,敲除pta菌株的生物量有明顯降低,由于該途徑與ATP合成相關(guān)[17],最終導(dǎo)致細(xì)胞內(nèi)能量失衡,不利于菌體生長。在此基礎(chǔ)上利用改造后的宿主研究對BT合成的影響。
2.2 以poxB基因缺失株為宿主的1,2,4-丁三醇合成
丙酮酸通過poxB基因編碼的丙酮酸氧化酶催化合成乙酸[16]。與對照菌株E.coliKXW3009相比,敲除poxB可以弱化乙酸,減少丙酮酸消耗增加流向TCA循環(huán)的碳流,提高胞內(nèi)ATP和還原力[18]。如圖2-a和圖2-b所示,poxB的缺失使乙酸最高積累量減少了37.9%,生物量提高了13.3%,且葡萄糖完全消耗,說明敲除poxB可以增強(qiáng)葡萄糖消耗,對生物量有明顯的正調(diào)控。其原因可能是增加了流向TCA循環(huán)的碳流,促進(jìn)了細(xì)胞代謝,減少了乙酸對細(xì)胞的毒害作用。另外,單敲poxB與對照菌E.coliKXW3009相比,殘留木糖減少了約85%,但木糖摩爾轉(zhuǎn)化率從0.75 mol/mol降低到0.67 mol/mol,木糖酸含量從6.3 g/L增加到6.8 g/L,提高了7.9%,其原因可能是該策略為木糖轉(zhuǎn)運(yùn)提供了更多能量,木糖消耗的增加促進(jìn)了中間代謝產(chǎn)物的積累。單敲poxB菌株BT產(chǎn)量由9.5 g/L提高到13.1 g/L,單位菌體產(chǎn)量增加了23.3%(圖2-a、圖2-b)。上述結(jié)果表明,敲除poxB減少乙酸合成的同時(shí)促進(jìn)了細(xì)胞生長、葡萄糖和木糖消耗,增強(qiáng)了BT合成能力,但是降低了木糖轉(zhuǎn)摩爾化率。
2.3 以pta基因缺失株為宿主的1,2,4-丁三醇合成
pta基因編碼的磷酸轉(zhuǎn)乙酰酶催化乙酰輔酶A轉(zhuǎn)化為乙酸[16]。與對照菌株E.coliKXW3009相比,敲除pta可以弱化乙酸途徑,減少乙酰輔酶A節(jié)點(diǎn)處碳流流向乙酸[18]。如圖2-a和圖2-c所示,pta缺失使乙酸最高積累量減少了84.5%。生物量降低了21.9%,葡萄糖和木糖殘留增加了72.7%和35.3%,說明pta缺失不利于菌體生長。其原因可能是由于乙酰輔酶A和丙酮酸通量不平衡[18],而反過來又會影響底物的消耗,同時(shí)Pta-AckA途徑是ATP合成的重要方式[14]。BT產(chǎn)量下降了24%,但是單位菌體產(chǎn)量和木糖轉(zhuǎn)化率提高了6.7%,木糖摩爾轉(zhuǎn)化率達(dá)到0.8 mol/mol,木糖酸含量降低了44%,說明菌體量的降低導(dǎo)致BT終產(chǎn)量的降低,但是增強(qiáng)了BT合成途徑,降低了中間代謝產(chǎn)物的積累。
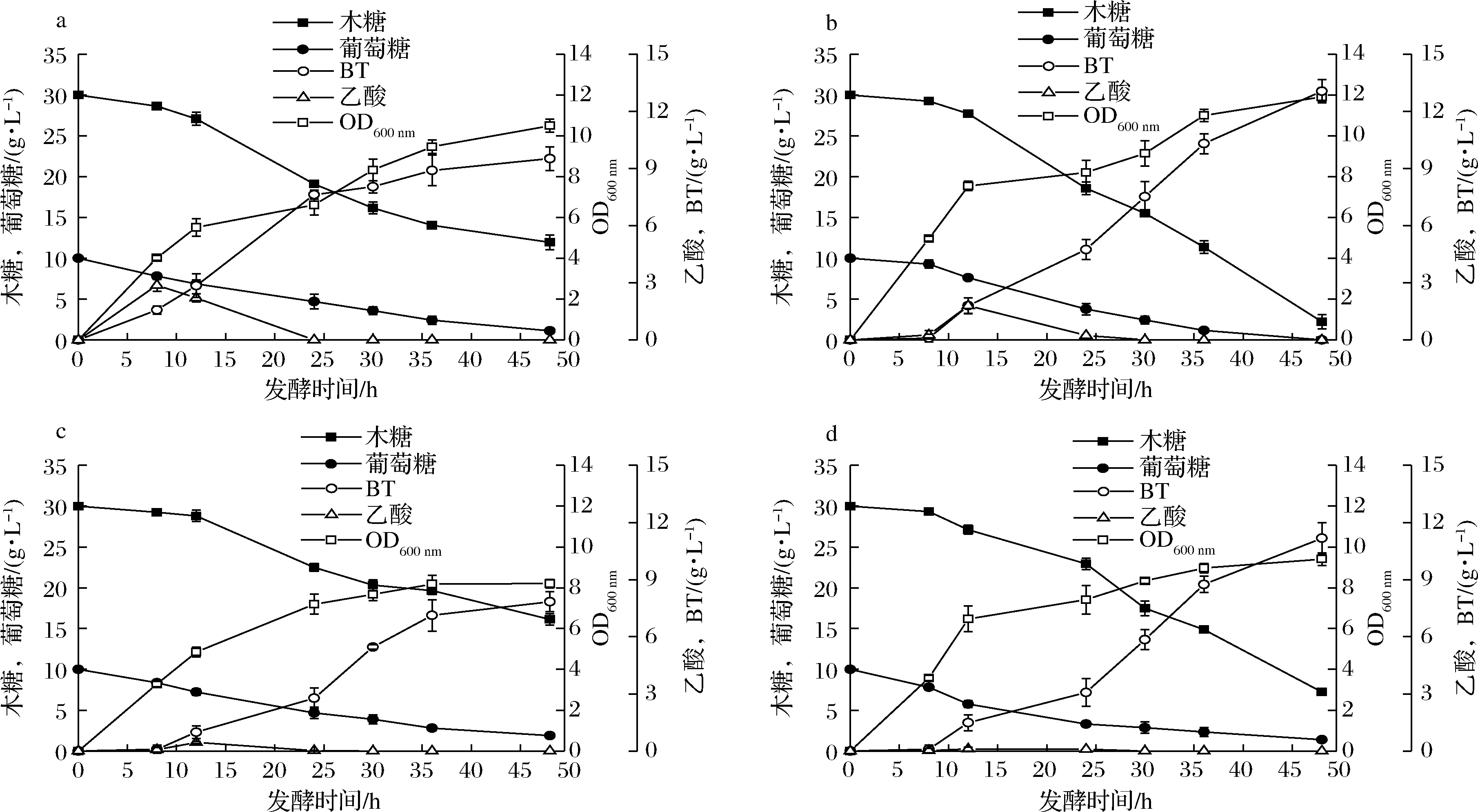
a-E.coli KXW3009;b-E.coli KXW3009B;c-E.coli KXW3009A;d-E.coli KXW3009AB圖2 搖瓶發(fā)酵代謝產(chǎn)物產(chǎn)量Fig.2 Metabolite production during shake flask fermentation
2.4 以共敲除pta和poxB基因缺失株為宿主的1,2,4-丁三醇合成
結(jié)合以上兩種策略,在E.coliW3009中雙敲pta和poxB基因,以期進(jìn)一步弱化乙酸,雙敲菌株積累的乙酸與單敲poxB和單敲pta相比分別降低了93.9%和75.5%,與對照菌株E.coliKXW3009相比降低了96.1%(圖2)。與對照相比,生物量降低了10.5%,說明共敲除pta和poxB對生物量有負(fù)調(diào)控,此結(jié)果與單敲pta類似。但是BT終產(chǎn)量和單位菌體產(chǎn)量分別提高了18.9%和30%,BT達(dá)到11.2 g/L,說明雙敲菌株可以進(jìn)一步提高胞內(nèi)還原力,強(qiáng)化BT合成途徑。
2.5 不同敲除菌株發(fā)酵比較及葡萄糖優(yōu)化
由表3可知,除單敲poxB外,敲除菌株生物量均低于對照菌株,而在pta缺失菌株中敲除poxB可以緩解菌株的生長缺陷,說明敲除poxB對生物量是正向調(diào)控的,可能是減少了丙酮酸的消耗,促進(jìn)了TCA循環(huán)。單敲pta菌株的葡萄糖和木糖利用率以及單位菌體木糖消耗是最低的但是摩爾轉(zhuǎn)化率最高,說明敲除pta雖然會降低生物量和底物攝取,但是強(qiáng)化了BT合成途徑,提高了木糖摩爾轉(zhuǎn)化率。
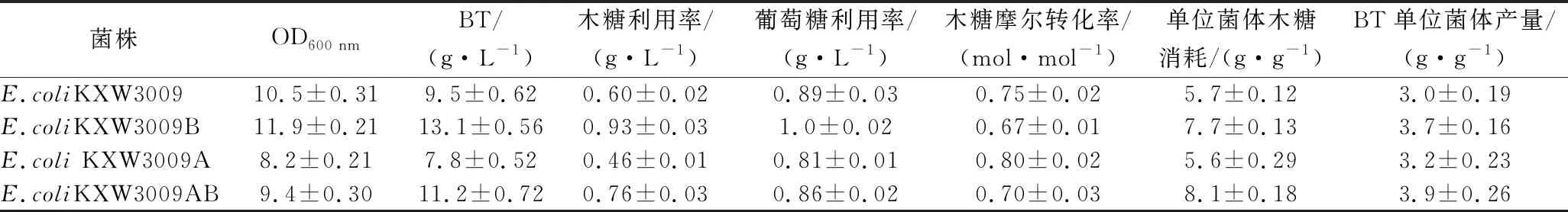
表3 不同菌株發(fā)酵參數(shù)Table 3 Comparison of parameters of strains
有研究表明,共敲除pta和poxB能夠進(jìn)一步提高乙酰輔酶A含量[19],而胞內(nèi)NAD(P)H水平直接影響B(tài)T的合成[20]。如表3所示,雙敲菌株BT單位菌體產(chǎn)量均高于單敲poxB和pta以及對照菌株E.coliKXW3009,分別提高了5.4%、21.8%、30%,單位菌體木糖消耗量分別提高了5.2%、44.6%、42.1%,表明進(jìn)一步弱化乙酸可以增加胞內(nèi)還原力,提高木糖利用能力,強(qiáng)化BT的合成。但過度抑制乙酸途徑反而會影響菌體生長,導(dǎo)致雙敲菌株BT終產(chǎn)量低于單敲poxB菌株,所以選擇E.coliKXW3009B為后續(xù)研究對象。
由于弱化乙酸途徑較大地影響到葡萄糖代謝能力,所以對培養(yǎng)基中葡萄糖添加量進(jìn)行優(yōu)化。如圖3-a和圖3-b所示,隨著葡萄糖初始質(zhì)量濃度的增加,改造菌株葡萄糖消耗也增加,同時(shí)菌株的OD600 nm也隨著增加,但是當(dāng)葡萄糖初始濃度達(dá)到15 g/L時(shí),生物量開始下降。另外,除了葡萄糖初始濃度為10 g/L時(shí),其他葡萄糖優(yōu)化條件下木糖殘留均增加(圖3-c)。
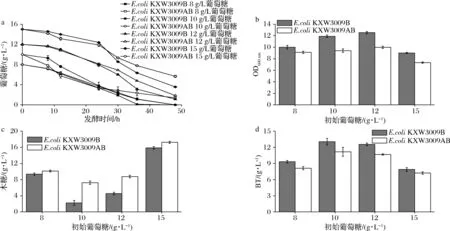
a-葡萄糖消耗;b-OD600 nm;c-木糖殘留;d-BT產(chǎn)量圖3 葡萄糖優(yōu)化搖瓶發(fā)酵代謝產(chǎn)物產(chǎn)量Fig.3 Glucose optimized shake flask fermentation metabolite yield
其原因可能是當(dāng)葡萄糖初始質(zhì)量濃度<10 g/L時(shí)菌株生物量降低,減少了木糖消耗。而當(dāng)葡萄糖初始>10 g/L時(shí),較高濃度的葡萄糖限制了木糖的消耗[3],進(jìn)而導(dǎo)致BT產(chǎn)量的降低。最終單敲poxB和雙敲菌株均在葡萄糖初始濃度達(dá)到10 g/L時(shí)BT產(chǎn)量達(dá)到最高,分別為13.1和11.2 g/L(圖3-d)。
2.6 5 L發(fā)酵罐分批補(bǔ)料發(fā)酵
對菌株E.coliKXW3009B、E.coliKXW3009AB以及對照菌株E.coliKXW3009進(jìn)行分批補(bǔ)料發(fā)酵。與對照菌株相比,E.coliKXW3009B和E.coliKXW3009AB中乙酸積累分別減少了34%和75%(圖4-a、圖4-b和圖4-c)。此外,最終E.coliKXW3009B的生物量提高了14.2%,而E.coliKXW3009AB生物量降低了15%,表明適度弱化乙酸途徑可以增加TCA循環(huán)的碳通量,為細(xì)胞生長提供了更多的碳源和能量,而過度弱化乙酸途徑會影響細(xì)胞生長。
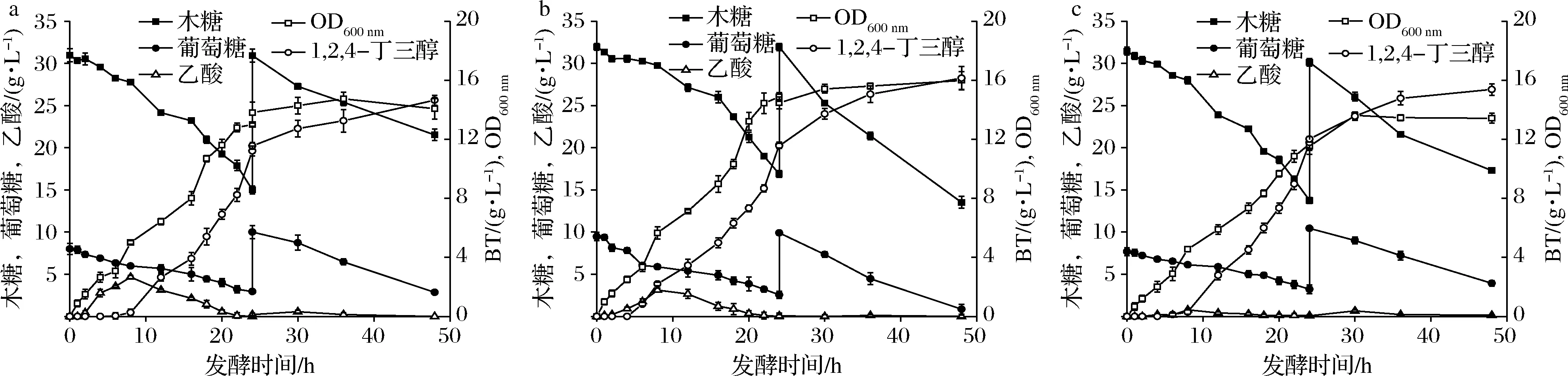
a-E.coli KXW3009;b-E.coli KXW3009B;c-E.coli KXW3009AB圖4 5 L發(fā)酵罐發(fā)酵過程中敲除菌株的代謝產(chǎn)物產(chǎn)量Fig.4 Metabolite yield of knockout strains during fermentation in 5 L fermenters
目前有研究通過對大腸桿菌中PTS系統(tǒng)進(jìn)行改造,發(fā)酵罐產(chǎn)量達(dá)到7.5 g/L[21]。通過在大腸桿菌中敲除基因mtfA,實(shí)現(xiàn)木糖和葡萄糖共利用,最終重組菌中BT產(chǎn)量和摩爾轉(zhuǎn)化率分別為7.2 g/L和0.55 mol/mol[20]。利用PyjhI驅(qū)動的遺傳開關(guān)合成BT,BT產(chǎn)量僅有1.3 g/L[22]。本論文通過弱化乙酸途徑可以減少碳源浪費(fèi)和對細(xì)胞的毒害作用[7],提高木糖消耗,促進(jìn)BT的合成,最終在5 L發(fā)酵罐分批補(bǔ)料發(fā)酵中E.coliKXW3009B和E.coliKXW3009AB的BT產(chǎn)量分別達(dá)到16.1和15.4 g/L,摩爾轉(zhuǎn)化率分別達(dá)到0.69 mol/mol和0.72 mol/mol。所以選擇E.coliKXW3009B為后續(xù)研究對象。
3 結(jié)論
本研究在E.coliW3009中敲除丙酮酸氧化酶poxB和磷酸轉(zhuǎn)乙酰酶pta減少了乙酸的合成,強(qiáng)化了TCA循環(huán),并考察了細(xì)胞生長和BT合成。通過弱化乙酸考察對細(xì)胞生長的影響,只有單敲poxB才能促進(jìn)細(xì)胞生長。在弱化乙酸E.coli基礎(chǔ)上考察對BT合成的影響,單敲或雙敲均可以提高BT單位菌體產(chǎn)量,并且雙敲高于單敲,說明弱化乙酸途徑強(qiáng)化了BT合成。敲除pta降低了80%以上的乙酸,但是影響了細(xì)胞生長,降低了木糖和葡萄糖的消耗以及BT終產(chǎn)量。而對生物量有促進(jìn)作用的單敲poxB菌株,BT最終產(chǎn)量達(dá)到13.1 g/L,摩爾轉(zhuǎn)化率為0.67 mol/mol。對敲除poxB的菌株進(jìn)行5 L發(fā)酵罐分批補(bǔ)料發(fā)酵,產(chǎn)量達(dá)到16.1 g/L,摩爾轉(zhuǎn)化率達(dá)到0.69 mol/mol。綜上,適宜的乙酸途徑弱化策略可以強(qiáng)化TCA循環(huán),葡萄糖消耗增加可以為BT合成提供更多NAD(P)H,但是過度抑制乙酸途徑反而會影響菌體生長,為今后進(jìn)一步代謝改造E.coli提高BT的產(chǎn)量提供了理論基礎(chǔ)和研究思路,并為以E.coli為底盤細(xì)胞的合成類似化合物的合成生物學(xué)改造提供借鑒。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
