尾葉香茶菜丙素對肝癌耐藥株HepG2/ADM細胞體外活性的影響
2021-10-30 05:52:24王欣玉裴曉東何志龍林佳雯黎永卓陳文卿劉小玲蔣利和
安徽醫藥 2021年11期
王欣玉,裴曉東,何志龍,林佳雯,黎永卓,陳文卿,劉小玲,蔣利和,3
惡性腫瘤是當今極難治愈的疾病之一,嚴重地威脅著人類的生命健康,是致死率第二高的疾病。其中肝癌作為第二大致死癌癥,僅次于肺癌。目前,我國已成了全球肝癌發病率和死亡率最多的國家,每年肝癌發病約34.7 萬人,占到全球的55%,死亡32.3 萬人,占全球的45%,晚期肝癌病人只有大約15% 的人適合手術切除,多數病人選擇放化療治療的方法。目前,化療藥物因其高毒性和耐藥性使病人預后較差,祖國傳統中藥材具有多靶點低毒性的優點,因此人們期待在傳統中草藥中有應用前景的天然產物。
香茶菜屬(Isodon
)植物隸屬唇形(Lamiaceae)科羅勒亞科(Ocimoideae),約有30 余種在我國和日本民間被廣泛使用,它們大多具有清熱解毒、抗炎抗菌及抗腫瘤等作用。其中尾葉香茶菜產于黑龍江、吉林及遼寧等地區,活性成分包括二萜類化合物尾葉香茶菜丙素(Kamebakaurin),尾葉香茶菜丙素對慢性骨髓性白血病(CML)有治療活性,且對心、肝、脾、肺、腎5 個臟器無副作用,還可減少撲熱息痛引起的肝損傷;在抑制腫瘤方面,可通過抑制HIF-1α 蛋白表達抑制結腸癌細胞HCT116 和SNu638 生長。但是在對耐藥腫瘤方面還未見文獻報道,因此本研究自2018 年11 月至2019 年11 月通過尾葉香茶菜丙素對HepG2/ADM 細胞的生長活力、遷移能力、細胞凋亡和周期的影響和可能機制,初步探明尾葉香茶菜丙素對耐藥肝癌的應用潛力。1 材料與方法
1.1 材料與試劑
尾葉香茶菜丙素購買于成都艾法公司,純度99%;阿霉素購自上海生工,純度99%。尾葉香茶菜丙素先用DMSO 溶解成200 μmol/L 濃度的母液,使用時用DMEM 稀釋。胎牛血清購自美國GIBCO 公司,HepG2/ADM 細胞購自上海中科院細胞庫,周期試劑盒購于聯科公司,CCK-8 購于MCE 公司,凋亡試劑盒購于BD 生物公司,BCA 蛋白濃度測定試劑盒和蛋白上樣緩沖液、RIPA 裂解液、蛋白酶抑制劑均購自索萊寶公司,PCR 引物由上海生工合成。抗體β-Tublin 多克隆抗體(貨號10068-1-AP),p-AKT 多克隆抗體(貨號66444-1-Ig),MDR1 多克隆抗體(貨號22336-1-AP),Bax 多克隆抗體(貨號50599-2-Ig),Bcl-2 多克隆抗體(貨號12789-1-AP),AKT 多克隆抗體(貨號10176-2-AP)GAPDH 多克隆抗體(貨號60004-1-Ig),PTEN 多克隆抗體(貨號22034-1-AP)均在Proteintech 購買;p-PTEN 單克隆抗體(貨號AP0930)購自AbClon生物公司。1.2 儀器與設備
DMI4000B 倒置熒光顯微鏡購于日本奧林巴斯公司;CO恒溫培養箱購自Eppendorf AG;低溫離心機購于廣州吉迪儀器有限公司;BGPower 300電泳儀產自貝基恩生物科技有限公司。1.3 細胞培養
HepG2/ADM 細胞用含10% 的胎牛血清、100 IU/mL 青霉素、100 mg/L 鏈霉素的高糖DMEM 完全培養基加入1 μg/mL 的阿霉素培養以保持細胞耐藥性,于37 ℃,5% 二氧化碳細胞培養箱內培養。1.4 CCK-8 試劑盒檢測HepG2/ADM 細胞增殖活力
取分裂期細胞以5×10/mL接種在96孔板中,每孔100 μL,每個濃度梯度平行設4 個復孔。37 ℃,5% 濃度二氧化碳培養箱中培養24 h,更換用新鮮DMEM 稀釋的含尾葉香茶菜丙素培養基或含阿霉素培養基,濃度梯度為培養24 h,后吸出培養基,加入濃度為10% 的含CCK-8 的新鮮的無血清DMEM,培養箱中孵育1 h后用酶標儀在450 nm 波長處測定吸光值。按下列公式計算各組藥物對細胞的抑制率(inhibition rate,IR)=(1-藥物組OD 值/細胞對照組OD值)×100%。實驗重復3次結果取平均值。1.5 細胞周期實驗
將HepG2/ADM 細胞接種于六孔板中,密度為5×10個/毫升,生長密度達到70% 時加入混有尾葉香茶菜丙素的新鮮培養基,分為四組:空白組,40 μmol/L 組、60 μmol/L 組、80 μmol/L組。培養24 h后收集細胞加入按1∶100濃度混合的PI 染色緩沖液,每個樣本加入500 μL 充分重懸細胞,37 ℃培養箱孵育25 min,上機檢測。實驗重復3次結果取平均值。1.6 流式細胞儀檢測HepG2/ADM 細胞凋亡
分裂期HepG2/ADM 細胞接種于6 孔板中,培養至密度70%后更換新鮮培養基并加入K,濃度分別為0、40、60、80 μmol/L。24 h 后收集細胞并用PBS 洗滌兩次,1 200 r/min,3 min。吸出PBS 后每孔加入Bind?ing Buffer 500 μL 重懸細胞,再加入5 μL Annexin VFITC 混合均勻,最后加入5 μL Propidium IAide。將細胞充分吹打散開后放入培養箱中37 ℃孵育15~30 min,取出后置于冰上,上機測定其凋亡率,重復3次取平均值。1.7 Transwell 小室實驗
消化收集分裂期的細胞,通過細胞計數吸取5 000 個細胞加入到小室中,其中上室200 μL 混有細胞的無血清培養基,下室500 μL 含有10% 血清的培養基。24 h 后停止實驗,無水乙醇固定10 min,1% 濃度的結晶紫染色10 min,PBS 清洗兩次每次8 min,之后進行常光拍照。實驗重復3次結果取平均值。1.8 細胞平板劃痕實驗
細胞接種于6 孔板,密度為5×10個/mL,待生長到密度90% 時用10 μL 白色槍頭劃線,每組畫三條,用PBS清洗兩次洗去浮起的細胞,加入混有尾葉香茶菜丙素的無血清培養基,設置四組為空白con 組、尾葉香茶菜丙素40 μmol/L組、60 μmol/L 組、80 μmol/L 組。在0、24、48 h 分別進行拍照記錄。1.9 實時熒光定量PCR
尾葉香茶菜丙素處理HepG2/ADM 細胞后24 h 提取RNA,之后RNA 反轉為DNA,預變性95 ℃3 min,95 ℃變性10 s,60 ℃退火30 s,95 ℃延伸30 s,總共40 個循環;之后69 ℃終延伸1 min。1.10 蛋白質印跡法(Western blotting)
六孔板中細胞生長在70% 時將培養液換為混有尾葉香茶菜丙素的新鮮培養基,尾葉香茶菜丙素濃度為40 μmol/L、60 μmol/L、80 μmol/L。37 ℃繼續培養24 h后吸出舊培養基,PBS 沖洗2 次加入RIPA 裂解液(RIPA∶蛋白酶抑制劑=1∶100),每孔加入200 μL。用細胞刮將細胞刮下后轉移至EP 管中4 ℃冰上裂解40 min,每隔10 min 用手指彈管壁使細胞充分裂解。用BCA蛋白濃度試劑盒測定蛋白濃度,加入4X蛋白上樣緩沖液沸水煮10 min,上樣跑膠,轉膜,5%脫脂奶粉或5%BSA 封閉,一抗4 ℃孵育過夜,1%TBST 洗膜3 次,二抗常溫孵育70 min,洗膜3 次,上機顯色拍照。1.11 統計學方法
采用SPSS 22.0 軟件,計量資料采用x ± s,
組間比較采用LSD-t
檢驗,多組間比較采用單因素方差分析,P
<0.05為差異有統計學意義。2 結果
2.1 尾葉香茶菜丙素可抑制HepG2/ADM 細胞的生長活力
為測定尾葉香茶菜丙素對HepG2/ADM細胞生長活力的影響,使用CCK-8 試劑盒測定,實驗顯示加入阿霉素濃度達到1.0 mg/mL 時對HepG2/ADM 細胞的抑制率只有26.5%(n
=3),而加入尾葉香茶菜丙素后肝癌細胞HepG2/ADM 細胞生長活力下降明顯(圖1),說明尾葉香茶菜丙素可抑制HepG2/ADM細胞生長活性且呈濃度依賴性,IC50為(62.97 ±2.81)μmol/L(n
=3)。同時,也表明尾葉香茶菜丙素對耐藥ADM 的HepG2 細胞具有抑制作用。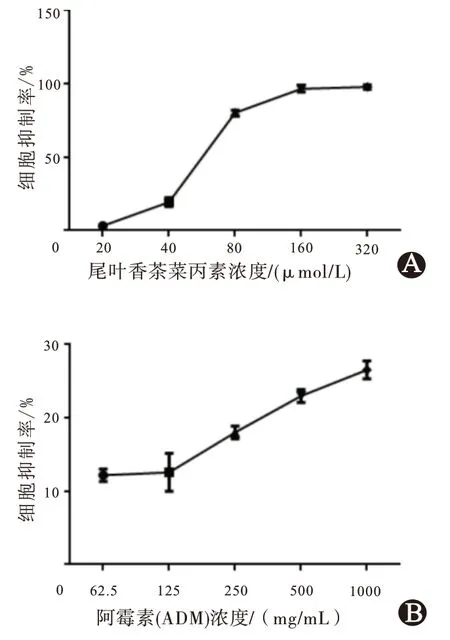
圖1 尾葉香茶菜丙素對HepG2/ADM細胞活力的影響(24 h):A為尾葉香茶菜丙素對HepG2/ADM細胞抑制率;B為阿霉素對HepG2/ADM細胞的抑制率
2.2 尾葉香茶菜丙素將HepG2/ADM 細胞周期阻滯在G2 期
通過流式細胞儀測定HepG2/ADM 細胞在增殖不同時期的數目比例分布,結果表明隨著尾葉香茶菜丙素濃度,HepG2/ADM 細胞被阻滯到G2 期,G2 期比例在空白組、40 μmol/L、60 μmol/L、80 μmol/L 分別增加(11.883±2.551)% 、(21.756±1.737)% 、(30.152±1.964)% 、(37.024±2.727)%(n
=3),呈現濃度依賴性的趨勢(F
=1.821,2.380,2.516,t
=6.189,5.221,4.287,P
=0.041,0.033,0.0017)。2.3 尾葉香茶菜丙素促進HepG2/ADM 細胞凋亡
利用流式細胞術分析不同濃度尾葉香茶菜丙素對細胞凋亡的影響,實驗結果顯示,在尾葉香茶菜丙素濃度分別為0、40、60、80 μmol/L 時,HepG2/ADM 細胞凋亡率分別為(6.267±1.317)%、(15.10±1.265)%、(19.77±1.963)%、(25.614±2.301)%(n
=3),凋亡率呈遞增趨勢,說明尾葉香茶菜丙素能誘導HepG2/ADM 細胞凋亡,且呈濃度依賴性。Western blotting 結果顯示Bcl-2 家族中促凋亡蛋白Bax 蛋白灰度值在尾葉香茶菜丙素濃度分別為0、40、60、80 μmol/L 時 分 別 為1、(1.24±0.072)、(1.63±0.128)、(2.19±0.183),說明Bax 蛋白表達上升,抑凋亡蛋白Bcl-2 在尾葉香茶菜丙素濃度分別為0、40、60、80 μmol/L 時 別 為1、(0.62±0.049)、(0.36±0.036)、(0.23±0.028),表達下降,顯示尾葉香茶菜丙素通過調節Bcl-2家族蛋白表達發揮促凋亡作用(F
=2.057,1.553,5.785,t
=4.757,12.58,10.07,P
=0.0089,0.002,0.005)。見圖2。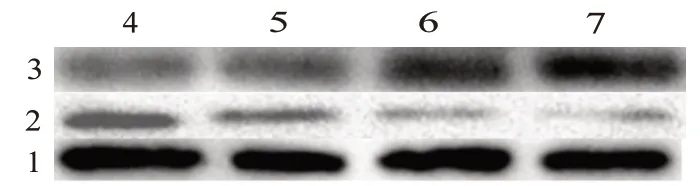
圖2 尾葉香茶菜丙素對HepG2/ADM細胞凋亡的促進作用
2.4 尾葉香茶菜丙素抑制HepG2/ADM 細胞遷移
為了研究尾葉香茶菜丙素對HepG2/ADM 細胞遷移的影響,研究進行了Transwell 小室實驗和平板劃痕愈合實驗,發現在K 濃度為0、40、60、80 μmol/L時,穿過膜的細胞數量分別為[(832.56±38.91)、(617.85±26.71)、(436.23±31.52)、(361.69±27.13)]個,在平板劃痕愈合實驗中發現,24 h時劃痕面積在K 濃度為0、40、60、80 μmol/L 分別是0 h 時的(0.68±0.012)、(0.83±0.021)、(0.90±0.019)、(0.95±0.010)倍;48 h 時劃痕面積在K 濃度為0、40、60、80 μmol/L分別是0 h 時的(0.41±0.022)、(0.56±0.024)、(0.72±0.031)、(0.88±0.016)倍,說明隨著尾葉香茶菜丙素濃度的升高,小室中穿過膜的細胞數量越來越少,劃痕愈合的速率也呈濃度依賴性的變慢,說明尾葉香茶菜丙素可以抑制HepG2/ADM 細胞的遷移(t=12.61,9.82,P=0.003,0.005)。見圖3。
圖3 尾葉香茶菜丙素對HepG2/ADM細胞遷移的影響:A為Tran?swell小室實驗;B為平板劃痕愈合實驗
2.5 尾葉香茶菜丙素能抑制MDR1 在轉錄和翻譯水平的表達
為了檢測尾葉香茶菜丙素是否調控耐藥基因的表達,我們首先進行了實時熒光定量PCR,表1 為引物合成表,結果顯示,BCRP、MRP1、MRP2 都未受到顯著調節,而在尾葉香茶菜丙素濃度為60 μmol/L 時MDR1 基因表達下調為空白對照的(0.255±0.013)倍(t
=20.41,P
=0.017)(n
=3)。West?ern blotting 結果顯示MDR1 蛋白的表達量也受到了抑制,在尾葉香茶菜丙素處理濃度在80 μmol/L 時MDR1蛋白的濃度為空白對照組的(0.352±0.089)倍(t
=19.87,P
=0.008)(n
=3),說明尾葉香茶菜丙素在轉錄和翻譯兩個水平都抑制了MDR1 的表達。見圖4。
表1 引物設計和合成
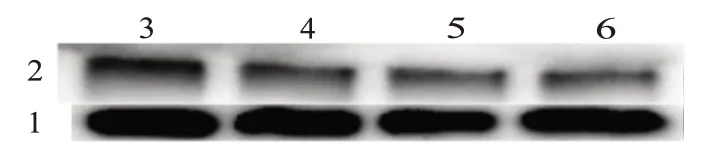
圖4 尾葉香茶菜丙素對ABC超家族轉運蛋白mRNA和蛋白質表達的影響
2.6 尾葉香茶菜丙素抑制能p-PTEN 和p-AKT 的表達
另外,Western blotting 結果顯示(圖5),尾葉香茶菜丙素處理HepG2/ADM 細胞后p-AKT 蛋白表達量下降在80 μmol/L 時p-AKT 蛋白表達量降至空白組的(0.231±0.036)倍(t
=15.49,P
=0.002),p-PTEN表達量上升,在60 μmol/L時上升為空白組蛋白表達量的(1.523±0.126)倍(t
=7.189,P
=0.002)且這些蛋白全部以濃度依賴性的方式受到調節。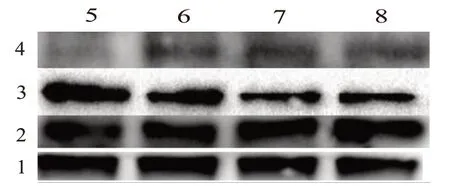
圖5 尾葉香茶菜丙素對PENT-AKT通路蛋白的調控
3 討論
肝癌耐藥性一直是其化療效果不佳和預后較差的重要原因,本研究發現阿霉素在1 mg/mL(184 μmol/L)時對HepG2/ADM 細胞耐藥株的生長抑制率只有26.5%,而尾葉香茶菜丙素(Kamebakurin)對肝癌細胞HepG2/ADM細胞生長活力有強抑制作用,其IC50 值為62.97 μmol/L,尾葉香茶菜丙素對HepG2/ADM細胞的毒性要高于一線藥物ADM。
在細胞凋亡主動中Bcl2 家族發揮重要作用,尾葉香茶菜丙素能誘導HepG2/ADM 細胞凋亡,蛋白免疫印跡結果顯示尾葉香茶菜丙素通過上調促凋亡蛋白Bax 的表達并抑制抑凋亡蛋白Bcl-2 表達;尾葉香茶菜丙素可將HepG2/ADM 細胞周期阻滯在G2期,說明尾葉香茶菜丙素使細胞DNA 損傷,細胞G2檢查點將受損細胞阻滯在G2 期無法進行后續有絲分裂,是HepG2/ADM 細胞發生凋亡和生長活力下降的主要原因之一;癌細胞的高遷移能力,是癌癥難以治愈的重要原因之一,我們的研究表明尾葉香茶菜丙素具有很強的抑制HepG2/ADM 細胞遷移的能力。MDR1在癌細胞中過表達是腫瘤化療成功的主要障礙之一,在腫瘤病人中,MDR1 將化療藥物排出細胞外,使藥物療效下降。在本研究中,尾葉香茶菜丙素處理細胞后,MDR1 在mRNA 和蛋白水平上顯著降低,逆轉了HepG2/ADM 細胞耐藥株將藥物泵出細胞的能力,說明尾葉香茶菜丙素具有逆轉HepG2/ADM 細胞耐藥的潛力,其部分功能通過抑制MDR1 基因和蛋白的表達發生,可能這是HepG2/ADM 細胞對尾葉香茶菜丙素要比阿霉素更敏感的原因之一。AKT 通路與很多腫瘤的發生相關,其異常激活會促進腫瘤的生長增殖以及遷移侵襲等,PTEN 是一種新發現的抑癌基因,作為AKT 通路的負調控因子其磷酸化水平的升高會抑制p-AKT 的激活從而抑制細胞增殖和遷移。通過Western blotting 實驗顯示尾葉香茶菜丙素處理細胞后,p-AKT含量下降且AKT通路上游蛋白p-PTEN表達上升最終抑制AKT通路的激活。
綜上所述,尾葉香茶菜丙素可能通過抑制MDR1 蛋白表達從而逆轉HepG2/ADM 細胞對藥物的敏感性,主要通過上調p-PTEN抑制p-AKT的激活從而促進HepG2/ADM 細胞的凋亡并抑制其遷移能力,說明尾葉香茶菜丙素具有很好的抑制肝癌耐藥的功效和潛力。
