木薯MeSAD基因的電子克隆與生物信息學分析
2021-11-22 11:03:38杜尚廣涂序堂曹文艷徐雅楠羅火林
南昌大學學報(理科版) 2021年4期
關鍵詞:分析
杜尚廣,涂序堂,熊 敏,余 波,曹文艷,徐雅楠,羅火林
(1.南昌師范學院生命科學學院,南昌 330000;2.南昌大學生命科學學院,南昌 330000)
木薯(ManihotesculentaCrantz),又名樹葛,是大戟科木薯屬多年生灌木,起源于南美洲亞馬遜流域,現已廣泛分布于熱帶及亞熱帶區域[1-2]。木薯光合效率高、塊莖中含有大量的淀粉,是7億人賴以生存的主糧之一[3-4],木薯在我國已有200余年的種植歷史,是廣東、廣西和海南等地區的一種重要糧食作物。隨著木薯產業的不斷發展,木薯廣泛應用于飼料、乙醇等生產,現已成為一種重要的工業原料[5-6]。木薯喜高溫、耐干旱貧瘠,但耐寒能力差;春潮早臨及倒春寒等低溫氣候將導致木薯減產、嚴重時甚至導致絕產,低溫現已成為木薯北移的主要限制因素[7-8]。目前,培育低溫耐受品種及其遺傳改良已成為世界性難題,因此克隆并研究木薯抗寒基因具有一定的理論意義和現實意義。
Δ9-硬脂酰-ACP脫氫酶(SAD蛋白)是植物抵抗逆境脅迫的重要酶,并在響應低溫過程中具有至關重要的作用[9]。低溫使植物細胞膜流動性下降,引起膜脂相變、降解,甚至導致植物死亡[10]。不飽和脂肪酸(unsaturated fatty acids,FAs)能夠提高生物膜的穩定性,防止變為凝膠相,維持細胞結構和功能的穩定[10]。SAD是一種可溶性蛋白,定位于質體基質,可將飽和的硬脂酰-ACP脂肪酸鏈還原成不飽和的油酰-ACP,提高植物中FAs的水平[3]。當前研究表明,植物的耐寒能力和細胞中FAs的水平具有一定的聯系[11]。將擬南芥SAD6基因轉入突變體中,能夠顯著提高FAs的水平,并減小低溫、干旱和缺氧等逆境的傷害[12]。研究發現,與野生型相比,SAD轉基因煙草(Nicotianatabacum)植株FAs的含量增高,并且其抗寒能力明顯增強[13]。對擬南芥[14]、花生(Arachishypogaea)[15]和小麥(Triticumaestivum)[16]等研究表明,SAD基因在抵抗低溫脅迫過程中發揮十分關鍵的作用,但尚未有關木薯SAD基因的報道。
本研究利用NCBI數據庫,以擬南芥SAD同源蛋白為探針,通過電子克隆技術獲得木薯SAD基因的cDNA序列。使用生物信息學方法,對表達蛋白進行一級至高級結構、理化性質和亞細胞定位等進行預測,并構建系統進化樹,分析木薯SAD基因的進化特征。因此,克隆木薯SAD基因對其耐寒基因的功能鑒定和遺傳改良提供參考,也為抗性品種的培育和篩選奠定基礎。
1 材料與方法
1.1 木薯MeSAD基因的cDNA獲取
以擬南芥的SAD氨基酸序列(登錄號:2281099)為探針,利用tblast在線工具搜索木薯的同源EST序列,選取一條Query cover最高、E值最低的同源序列,并以之為誘餌,利用blast在線搜索能覆蓋誘餌的EST序列,利用DNAMAN拼接,繼續blast直至無新的EST序列可拼接,得到SAD基因的全cDNA序列。本實驗所用的EST序列包括:DV455612.1、FF381161.1、FF380418.1、DB941776.1、DB938249.1、DB938249.1、DV456201.1和DB937741.1。
1.2 理化性質分析、疏水性/親水性分析
分別利用DNAMAN和ORF finder軟件完成cDNA和開放閱讀框(open reading frame,ORF)的翻譯和讀取;利用ProtParam tool在線工具完成蛋白質理化性質的預測;利用ProtScale軟件完成蛋白質的親水性/疏水性分析。
1.3 亞細胞定位和跨膜區分析、磷酸化位點與信號肽分析
分別利用ProtComp 9.0和TMHMM等在線工具進行表達蛋白亞細胞定位和跨膜區的分析;分別利用NetPhos 3.1、SignalP 4.1完成磷酸化位點與信號肽的預測和分析。
1.4 二級及高級結構的預測
利用NCBI的CDD在線工具進行結構域的預測和分析;利用SOPMA、SWISS-MODEL和VMD 1.9.3進行二級及高級結構的分析。
1.5 基因的系統進化樹構建
利用ClustalX 2.0軟件完成木薯和其他15種植物SAD蛋白序列的多重比對分析[6-7];利用MEGA X進行系統發育分析,進化樹遺傳距離、檢驗方法、循環次數分別設置為Kimura雙參數模型、Bootstrap method、1 000。
2 結果
2.1 基因全cDNA獲取
在NCBI數據庫中,利用擬南芥SAD氨基酸序列作為探針進行比對,選取相似度最高的序列,以之為探針繼續比對,選擇7條親緣關系較近的EST進行拼接和衍生;得到木薯SAD基因的全cDNA序列(圖1A)。cDNA序列總長為1 685 bp,ORF為176~1 366 bp(圖1B);翻譯蛋白含396個氨基酸,起始和終止密碼子分別是ATG和TAA。新基因命名及表達蛋白名分別為MeSAD、MeSAD。
2.2 理化性質分析
使用ProtParam tool工具預測MeSAD蛋白的理化性質可知。表達蛋白包含396個氨基酸,由20種氨基酸構成,含量最高和最低的氨基酸分別為亮氨酸(9.8%)、半胱氨酸(0.8%)。分子式是C2031H3181N545O597S15、分子量是45.3 kDa;脂肪系數是81.82;負電荷殘基是58、正電荷殘基是52;等電點(Theoretical pI)是5.93,呈酸性;不穩定系數是37.55;平均總親水性是-0.443。

圖1 (A)木薯SAD基因的cDNA拼接圖、(B)cDNA序列及推導的氨基酸序列
2.3 疏水性/親水性分析
分析MeSAD蛋白的疏水性/親水性可知(圖2A),第132位纈氨酸(Val)以及第265位的組氨酸(His)分別具有最高得分(2.522)和最低得分(-2.478),平均疏水指數為-0.443。即該蛋白質屬于親水性蛋白。
2.4 亞細胞定位和跨膜區分析
分析MeSAD蛋白的亞細胞定位可知,葉綠體的積分值最大(9.28)(表1);分析圖2B可知,該蛋白無跨膜區域。
2.5 磷酸化位點與信號肽分析
分析MeSAD蛋白潛在的磷酸化位點可知(圖3A),絲氨酸、蘇氨酸和酪氨酸磷酸化位點數目分別是19、13、5。信號肽分析可知(圖3B),該蛋白質無信號肽。
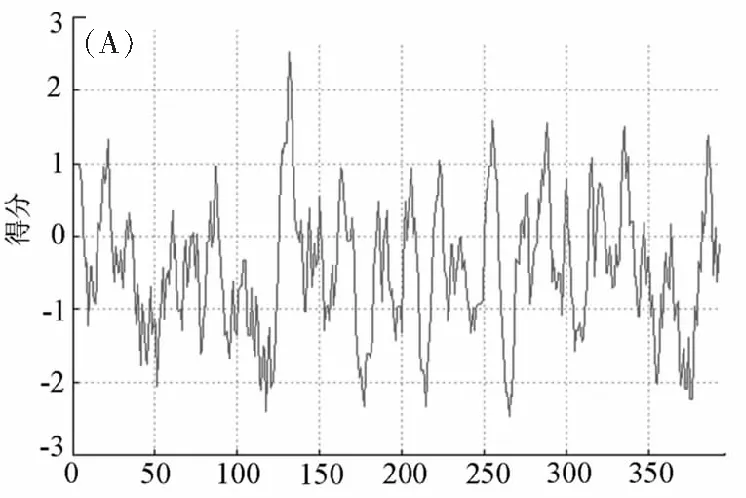
氨基酸位點
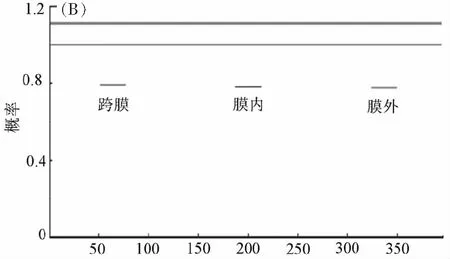
氨基酸位點圖2 (A) MeSAD蛋白疏水性/親水性預測、(B)跨膜區域的預測
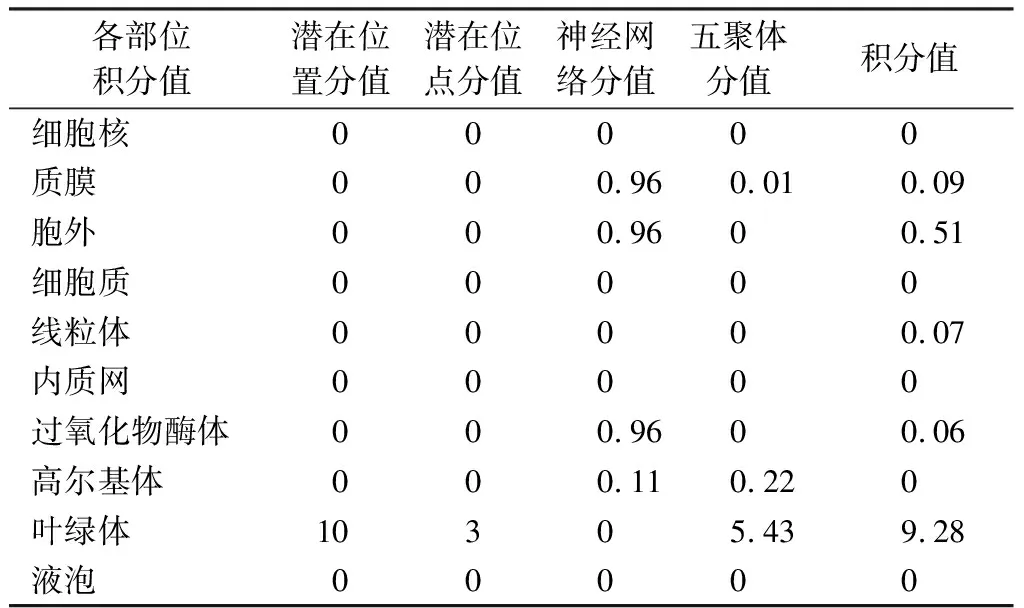
表1 MeSAD蛋白亞細胞定位預測

氨基酸位點
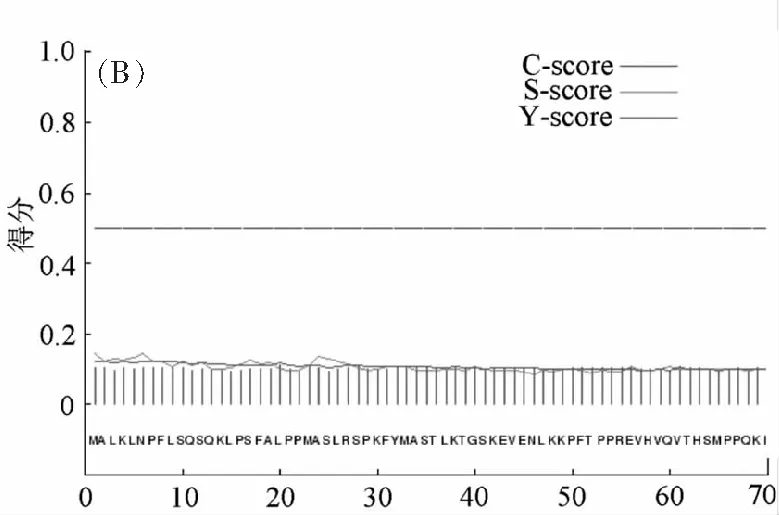
氨基酸位點圖3 (A) MeSAD蛋白的磷酸化位點預測、(B) 信號肽預測
2.6 二級及高級結構的預測
分析MeSAD蛋白二級結構可知,多肽鏈結構中含有α螺旋(53.28%)、無規則卷曲序列(34.09%)、延伸鏈(7.58%)和β轉角(5.05%)等(圖4A)。選擇“2j2f.pdb”模板,采用SWISS-MODEL在線軟件進行高級結構分析,其氨基酸序列一致性為60%,覆蓋度為92%,SAD主要由α螺旋以及無規則卷曲序列構成;使用VMD 1.9.3軟件繪制高級結構(圖4B)。
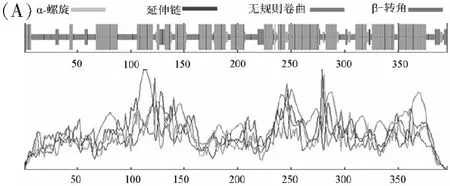
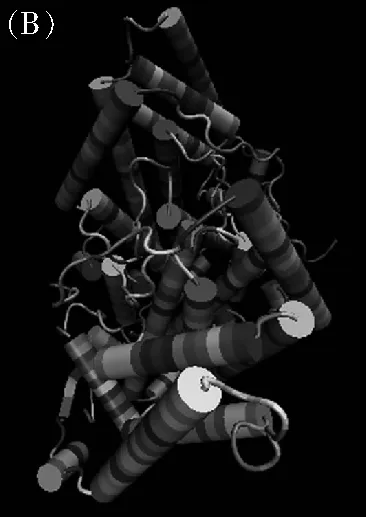
圖4 (A) MeSAD蛋白的二級結構預測、(B)三級結構預測
2.7 氨基酸同源性分析及進化樹構建
在NCBI數據庫中進行MeSAD和其他植物的SAD氨基酸同源序列比對,木薯與蓖麻(RicinuscommunisNP_001310659.1)、巴西橡膠樹(HeveabrasiliensisXP_021688869.1)、麻瘋樹(JatrophacurcasKDP44234.1)、可可(TheobromacacaoEOY04657.1)、臍橙(CitrussinensisXP_006494043.1)、溫州蜜柑(CitrusunshiuGAY44796.1)、克萊門柚(CitrusclementinaXP_006442769.1)序列一致性分別為95.71%,95.96%,95.20%,92.68%,90.66%,90.66%,90.66%。多重比對發現,以上物種的SAD具有很高的同源性,并且1~40序列部分同源,40~396序列高度同源(圖5A)。利用NCBI的CDD在線工具分析可知(圖5B),SAD屬于類鐵蛋白家族,具有酰基-ACP脫氫酶結構域(3~396),主要包括6個二鐵結合位點、33個底物結合位點以及14個底物結合口袋位點。
為探究SAD基因的進化特征,本研究基于SAD蛋白一級序列進行系統進化樹的構建,結果表明,SAD基因的進化和所選物種的進化是一致的(圖5C)。木薯與蓖麻、巴西橡膠樹處于同一分支,這3種植物同屬于大戟科;樹棉(Gossypiumarboreum)、海島棉(Gossypiumbarbadense)和陸地棉(Gossypiumhirsutum)處于同一分支,這3種植物同屬于錦葵科;甜瓜(Cucumismelo)、南瓜(Cucurbitamoschata)和筍瓜(Cucurbitamaxima)同屬于葫蘆科,聚在另一分支;大豆(Glycinemax)、木豆(Cajanuscajan)和水黃皮(Pongamiapinnata)同屬于豆科,亦聚在另一分支。這與它們在植物進化系統所處位置完全吻合。
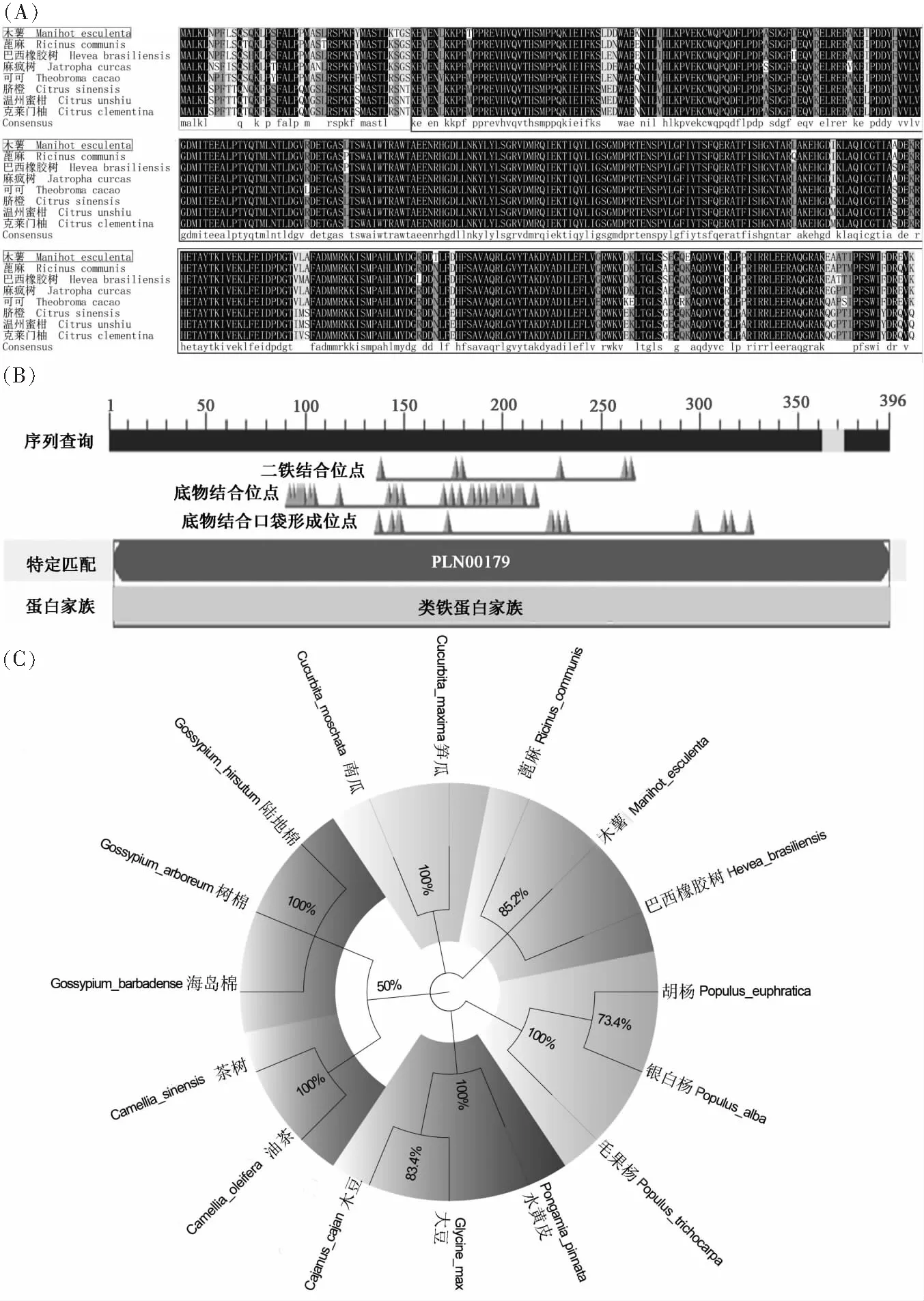
圖5 (A)不同植物中SAD氨基酸序列多重比對、保守結構域預測以及系統進化樹
3 討論
SAD基因在植物響應低溫脅迫中起到至關重要的作用[17]。迄今為止,SAD蛋白是植物中唯一發現的脂質去飽和酶,SAD催化硬脂肪酸脫氫生成油酸,油酸被運輸至類囊體或細胞質進一步去飽和,以此提高其抗寒水平[17]。關于SAD基因的脂質去飽和化研究現已成為生物化學及分子生物學的科研熱點,克隆SAD基因具有十分重要的意義[18]。
以往研究表明,SAD蛋白是植物中較為保守的一類蛋白,具有典型的同型二聚體結構以及4個α螺旋與Fe2+結合構成的活性中心[19-21],本研究發現木薯的預測蛋白中具有這類典型的結構。SAD蛋白主要是靠三級結構的Fe2+活性中心來行使功能[22-24],這說明木薯預測蛋白具有SAD的功能。
在GENEBANK數據庫中,SAD蛋白的預測結果有兩個,登錄號分別為XP_021610569和XP_021628521。這兩條序列是通過二代基因組測序預測而來的,成本高昂,并耗費大量的人力和物力。本研究所預測的蛋白質序列是基于公共數據庫中的EST序列拼接的,零成本地預測SAD蛋白序列。經過與GENEBANK數據庫中已有序列相比,和其一致性分別為98%和95%,這表明電子克隆可經濟有效地預測基因序列。
本研究利用電子克隆技術克隆木薯SAD基因并進行生物信息學分析,具有一定的科學意義。本研究的結果擬為木薯基因克隆、表達及干擾載體構建提供參考,并為木薯SAD轉基因研究、抗寒機理闡述以及品種改良奠定基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
