過(guò)表達(dá)DcR3對(duì)肝癌細(xì)胞自噬功能的影響*
2021-11-26 09:21:14張玉達(dá)趙靜靜
檢驗(yàn)醫(yī)學(xué)與臨床 2021年22期
張玉達(dá),彭 亮,趙靜靜
上海市松江區(qū)中心醫(yī)院檢驗(yàn)科,上海 201600
肝癌是世界上最常見(jiàn)的惡性腫瘤之一。據(jù)統(tǒng)計(jì),2018年肝癌的發(fā)病率在所有癌癥中排第6位,是全球第4大癌癥死亡原因,每年約有78.2萬(wàn)人死于肝癌[1]。肝癌的臨床治療包括肝切除、肝移植、放化療等,治療效果差且易復(fù)發(fā)[2-3],晚期肝癌患者5年生存率低于5%,預(yù)后極差。以中國(guó)為首的亞洲發(fā)展中國(guó)家是肝癌的高發(fā)地區(qū)[4]。肝癌嚴(yán)重威脅著人們的健康,明確其發(fā)病機(jī)制,有針對(duì)性地采取早期干預(yù)措施是當(dāng)前研究的熱點(diǎn)。
前期研究發(fā)現(xiàn),肝癌誘騙受體3(DcR3)的表達(dá)量越高,其對(duì)腫瘤細(xì)胞凋亡的抑制作用越顯著[5]。有研究發(fā)現(xiàn),下調(diào)DcR3可以抑制肝癌細(xì)胞的侵襲性,增強(qiáng)腫瘤壞死因子相關(guān)凋亡誘導(dǎo)配體(TRAIL)介導(dǎo)的細(xì)胞凋亡[6]。在先前的研究中發(fā)現(xiàn),DcR3促進(jìn)了細(xì)胞的增殖和遷移,抑制了細(xì)胞凋亡指標(biāo)半胱氨酸蛋白酶3(caspase 3)和Bcl-2相關(guān)X(Bax)蛋白的表達(dá)[5]。自噬在真核細(xì)胞中廣泛存在,是一種基本生命現(xiàn)象,微管相關(guān)蛋白輕鏈3B(LC3B)和程序性死亡受體1(Beclin1)是自噬途徑的關(guān)鍵物質(zhì)[7]。LC3B以LC3BⅠ和LC3BⅡ兩種形式存在并參與自噬,LC3BⅠ在正常情況下定位于細(xì)胞質(zhì),當(dāng)自噬被誘導(dǎo)時(shí),LC3B的細(xì)胞質(zhì)形式LC3BⅠ通過(guò)與脂質(zhì)分子磷脂酰乙醇胺(PE)結(jié)合轉(zhuǎn)化為L(zhǎng)C3BⅡ,靶向定位于自噬體膜,介導(dǎo)自噬溶酶體的形成[8]。在細(xì)胞的生理過(guò)程中,自噬、凋亡、程序性壞死都是細(xì)胞的死亡形式,它們相互影響、相互調(diào)節(jié),自噬可能在細(xì)胞死亡過(guò)程中扮演重要角色。本研究通過(guò)在肝癌細(xì)胞中過(guò)表達(dá)DcR3來(lái)觀察其對(duì)自噬功能的影響,為肝癌的早期診斷和預(yù)后判斷提供理論基礎(chǔ)。
1 材料與方法
1.1一般材料 人肝癌細(xì)胞株HepG2(北京中科質(zhì)檢生物技術(shù)有限公司)置于37 ℃、5%CO2培養(yǎng)箱中常規(guī)培養(yǎng)。
1.2儀器與試劑 含各種氨基酸和葡萄糖的培養(yǎng)液(DMEM),胎牛血清,胰蛋白酶(美國(guó)Hyclone公司);DcR3過(guò)表達(dá)慢病毒載體和空載慢病毒均購(gòu)于上海吉?jiǎng)P基因化學(xué)技術(shù)有限公司;DcR3抗體、Beclin1抗體(Abcam 公司,美國(guó)); LC3B抗體(美國(guó)Cell signaing公司);辣根過(guò)氧化物酶(HRP)標(biāo)記親和純化山羊抗小鼠IgG二抗(美國(guó)sigma公司);反轉(zhuǎn)錄聚合酶鏈反應(yīng)(RT-PCR)試劑盒。RT-PCR引物序列合成(上海生工生物工程有限公司)。
1.3方法
1.3.1細(xì)胞轉(zhuǎn)染 肝癌細(xì)胞株HepG2以每孔3×105個(gè)分布于六孔板,當(dāng)細(xì)胞融合達(dá)到70%時(shí),分別感染DcR3過(guò)表達(dá)慢病毒載體和空載慢病毒。對(duì)照組為L(zhǎng)V-NC組(感染空載慢病毒),實(shí)驗(yàn)組為L(zhǎng)V-DcR3組(感染DcR3過(guò)表達(dá)慢病毒載體)。以病毒滴度最小致死量(MIO)50 IU/mL加入病毒上清液(參照預(yù)實(shí)驗(yàn)結(jié)果)。72 h 后收集LV-NC組和LV-DcR3 組細(xì)胞,然后進(jìn)行相關(guān)實(shí)驗(yàn)。
1.3.2免疫印跡試驗(yàn)(Western blot) 用預(yù)冷卻的磷酸鹽緩沖液(PBS)沖洗收集的細(xì)胞3次,然后用細(xì)胞裂解液(RIPA)裂解細(xì)胞,測(cè)定蛋白量,加入5×上樣緩沖液進(jìn)行混勻,在100 ℃變性10 min,取30 μg蛋白進(jìn)行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE),然后轉(zhuǎn)膜,將硝酸纖維素膜置于含5%脫脂奶粉的緩沖液中封閉2 h,吸取干凈封閉液后加入一抗(均為1∶1 000),4 ℃搖勻過(guò)夜。洗滌緩沖液(TBST)洗膜3遍,每次10 min,分別加入稀釋后的二抗IgG(1∶1 000),室溫避光孵育2 h。孵育后TBST洗膜,電化學(xué)發(fā)光法(ECL)顯色蛋白條帶,然后進(jìn)行凝膠成像分析,共進(jìn)行平行實(shí)驗(yàn)3次。
1.3.3RT-PCR Trizol法提取細(xì)胞總RNA,根據(jù)反轉(zhuǎn)錄試劑盒說(shuō)明書(shū)將總RNA的1 μg mRNA反轉(zhuǎn)錄為cDNA,反轉(zhuǎn)錄條件為42 ℃ 60 min,70 ℃ 5 min。PCR擴(kuò)增:95 ℃預(yù)變性30 s;94 ℃變性15 s,56 ℃退火45 s,72 ℃延伸45 s,擴(kuò)增40個(gè)循環(huán)。引物序列見(jiàn)表1。基于ABI7500平臺(tái)收集RT-PCR數(shù)據(jù),以甘油醛-3-磷酸脫氫酶(GAPDH)為對(duì)照,采用2-ΔΔCt計(jì)算相對(duì)表達(dá)水平。

表1 引物序列

2 結(jié) 果
2.1DcR3蛋白表達(dá)情況 本研究選擇DcR3表達(dá)較低且實(shí)驗(yàn)常用的肝癌細(xì)胞株HepG2作為研究對(duì)象。DcR3過(guò)表達(dá)慢病毒載體轉(zhuǎn)染72 h后,用Western Blot驗(yàn)證轉(zhuǎn)染情況。結(jié)果顯示,在相同印跡上以肌動(dòng)蛋白B(ACTB)信號(hào)標(biāo)準(zhǔn)化成像條帶為標(biāo)準(zhǔn),測(cè)定免疫印跡條帶的灰度值,LV-DcR3組DcR3蛋白的表達(dá)水平(1.88±0.15)明顯高于LV-NC組(0.74±0.05),差異有統(tǒng)計(jì)學(xué)意義(P<0.05),說(shuō)明DcR3慢病毒轉(zhuǎn)染效率達(dá)到實(shí)驗(yàn)要求。見(jiàn)圖1。
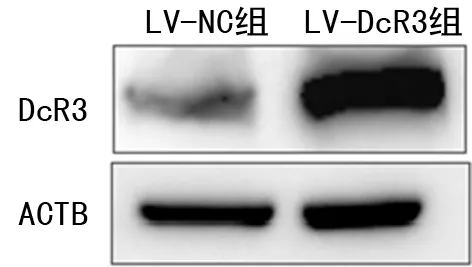
圖1 不同病毒感染細(xì)胞組的蛋白表達(dá)水平
2.2過(guò)表達(dá)DcR3對(duì)肝癌細(xì)胞自噬相關(guān)基因mRNA的影響 肝癌細(xì)胞自噬相關(guān)蛋白LC3B和Beclin1的mRNA相對(duì)表達(dá)水平結(jié)果顯示,LV-DcR3組LC3B和Beclin1 mRNA相對(duì)表達(dá)水平與LV-NC組相比,明顯下降,差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。見(jiàn)圖2。
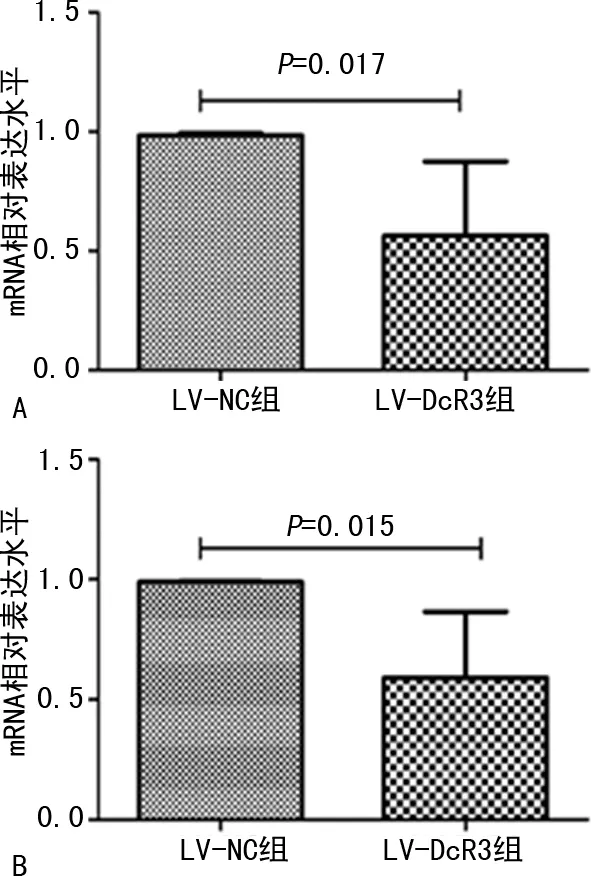
注:A為Beclin1;B為L(zhǎng)C3B。
2.3從蛋白相對(duì)表達(dá)水平觀察過(guò)表達(dá)DcR3對(duì)肝癌細(xì)胞自噬功能的影響 Western Blot結(jié)果顯示,在相同印跡上以ACTB信號(hào)標(biāo)準(zhǔn)化成像條帶為標(biāo)準(zhǔn),與LV-NC組比較,LV-DcR3組LC3BⅡ/LC3BⅠ比例(1.65±0.15)、Beclin1蛋白相對(duì)表達(dá)水平(0.89±0.05)顯著低于LV-NC組(LC3BⅡ/LC3BⅠ比例為3.48±0.31、Beclin1蛋白相對(duì)表達(dá)水平為1.41±0.05),差異均有統(tǒng)計(jì)學(xué)意義(P<0.05)。見(jiàn)圖3。
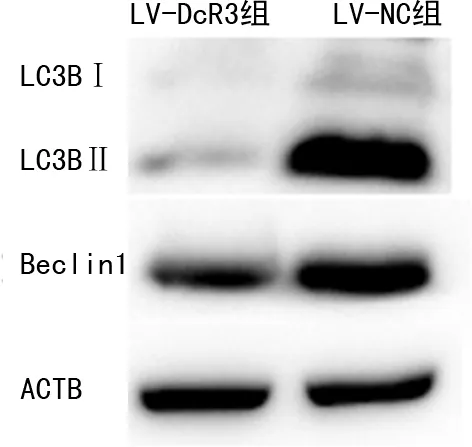
圖3 過(guò)表達(dá)DcR3的肝癌細(xì)胞自噬蛋白的相對(duì)表達(dá)水平
3 討 論
DcR3是腫瘤壞死因子受體超家族(TNFR)的成員之一,可與Fas配體(FasL)、腫瘤壞死因子配體相關(guān)分子1A(TLlA)競(jìng)爭(zhēng)結(jié)合,從而阻斷以上配體誘導(dǎo)的凋亡[6]。有研究認(rèn)為肝癌組織中DcR3的表達(dá)水平高于癌旁組織及正常組織,且與腫瘤分化、漿膜浸潤(rùn)、肝外轉(zhuǎn)移相關(guān)[7]。然而,DcR3在肝癌進(jìn)展和轉(zhuǎn)移中的確切機(jī)制仍不清楚。
近年來(lái),自噬作為細(xì)胞死亡方式的一種,成為腫瘤研究的熱點(diǎn)。已有研究報(bào)道,自噬在促進(jìn)腫瘤進(jìn)展或抑制腫瘤轉(zhuǎn)化中發(fā)揮雙重作用,提示自噬缺陷誘發(fā)腫瘤是一種促癌機(jī)制[8]。有研究表明,自噬在肝臟疾病從慢性肝病發(fā)展為肝硬化,最終發(fā)展為肝癌的病理機(jī)制中起著重要的促死亡(促癌)作用[9]。Beclin-1和微管相關(guān)蛋白1輕鏈3(LC3)是自噬途徑的關(guān)鍵物質(zhì)[10]。筆者推測(cè)DcR3可能影響自噬的過(guò)程。本實(shí)驗(yàn)通過(guò)在肝癌細(xì)胞中過(guò)表達(dá)DcR3以驗(yàn)證此推測(cè)。
LC3B被認(rèn)為是參與自噬的關(guān)鍵因素,是監(jiān)測(cè)自噬的特殊指標(biāo)。本研究結(jié)果顯示,過(guò)表達(dá)DcR3時(shí),LC3B蛋白和mRNA相對(duì)表達(dá)水平均降低。有研究表明,LC3B在腫瘤區(qū)較正常鄰近組織表達(dá)水平更低[11],與本研究結(jié)果一致。此外,一篇Meta分析顯示,LC3B的表達(dá)水平與性別、年齡、腫瘤數(shù)量、乙型肝炎表面抗原(HBsAg)、肝硬化、TNM分期、甲胎蛋白、血管侵犯、組織學(xué)分級(jí)和肝功能無(wú)關(guān)(P>0.05),而與總生存期有關(guān)(P<0.05)[12]。
Beclin1是首個(gè)被發(fā)現(xiàn)的哺乳動(dòng)物自噬蛋白,在自噬活性中起著重要的調(diào)節(jié)作用,在小鼠肝癌模型中發(fā)現(xiàn)Beclin1發(fā)揮抑癌作用[13]。肝癌組織中Beclin1表達(dá)水平明顯低于癌旁組織,Beclin1表達(dá)水平與復(fù)發(fā)及生存率相關(guān)[14]。本研究結(jié)果顯示,DcR3過(guò)表達(dá)會(huì)抑制Beclin-1的表達(dá)。因此,這些結(jié)果表明,DcR3抑制了肝癌細(xì)胞中Beclin1和LC3B的表達(dá),提示自噬可能受到DcR3的調(diào)控。
綜上所述,本研究通過(guò)體外實(shí)驗(yàn)初步驗(yàn)證了DcR3可以抑制自噬的發(fā)生,但由于是初步研究結(jié)果,LC3B和Beclin-1是否受DcR3調(diào)控,有待進(jìn)一步研究證實(shí)。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
天津醫(yī)科大學(xué)學(xué)報(bào)(2019年3期)2019-08-13 06:53:08
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
人大建設(shè)(2019年12期)2019-05-21 02:55:32
腫瘤預(yù)防與治療(2015年1期)2015-09-26 07:26:20
中國(guó)當(dāng)代醫(yī)藥(2015年16期)2015-03-01 02:03:11
癌變·畸變·突變(2015年4期)2015-02-27 06:15:25

- 檢驗(yàn)醫(yī)學(xué)與臨床的其它文章
- G6PC基因復(fù)合雜合突變致糖原累積病Ⅰa型1例
- 血清正五聚蛋白3在類(lèi)風(fēng)濕關(guān)節(jié)炎診斷中的價(jià)值
- 復(fù)發(fā)高級(jí)別膠質(zhì)瘤應(yīng)用IMRT再程放療的臨床評(píng)價(jià)*
- AZF區(qū)微缺失檢測(cè)方法分析Y-STR基因座分型缺失現(xiàn)象*
- 實(shí)驗(yàn)診斷學(xué)的教學(xué)改革與實(shí)踐
- PBL聯(lián)合CBL在臨床基礎(chǔ)檢驗(yàn)形態(tài)學(xué)實(shí)習(xí)教學(xué)中的應(yīng)用*