銅離子對肺癌細胞增殖的影響及相關基因的臨床意義
2021-12-07 10:00:52王桂珍周光飚
中國癌癥雜志 2021年11期
吳 冉,王桂珍,程 昕,3,周光飚
1.貴州大學醫學院生物醫學系,貴州 貴陽 550025;
2.國家癌癥中心/中國醫學科學院腫瘤醫院分子腫瘤學國家重點實驗室,北京 100021;
3.哈佛醫學院達納-法伯癌癥研究所癌癥生物學系,馬薩諸塞州 波士頓 02215
不同微量元素在生物體內發揮不同的生物學功能,并在一定程度上參與癌癥的發生、發展[1]。銅作為酪氨酸激酶及銅藍蛋白等多種酶的關鍵輔助因子,在生物體的生長發育過程中發揮重要作用。有研究[2]表明,銅是維持機體正常生物學功能的必需微量元素,具有氧化還原性質,能維持生物體內催化、氧化、細胞呼吸及各類生命活動。此外,也有研究[3]指出,銅離子能調節腫瘤的氧化磷酸化和生長過程,可參與致癌信號轉導通路BRAF轉導和腫瘤發生[4],對生物體有一定的潛在毒性[5]。因此,生物體的銅離子含量應該維持在一個相對穩定的狀態,否則將會破壞生物體的內環境穩態,引起機體的應激反應,從而造成不必要的傷害。
研究表明,破壞銅離子在體內的平衡可能會促進癌癥的發生、發展。由于銅離子在許多生物學過程中是重要的輔助因子,在不同的癌癥表型中常會出現銅離子的身影,例如銅可以通過激活參與細胞生長和代謝的酶LOX、SPARC、MEK1和MEMO1來增強癌細胞轉移的能力[6]。飲食中銅和鋅的攝入量與患肺癌風險相關[7-8]。銅是多種金屬酶活性必需的微量元素之一,血清中銅或鐵含量較高的人,會增加死于癌癥的風險[9]。此外,研究發現,肺癌患者血清中銅離子含量比健康對照組高[10-11],且肺癌患者癌組織中的銅、鈣、鎂和鋅含量也顯著高于正常肺組織[12-13]。但這些研究主要報道了患者血清或癌組織中的主要/微量元素含量的異常變化,而在細胞水平上研究銅離子濃度對細胞的增殖及信號轉導通路影響的報道較少。肺癌是全世界最常見的癌癥,每年有210萬新發及180萬死亡病例,在中國,肺癌也是最常見的癌癥,每年新發及死亡病例分別為78.7萬及63.1萬[14-15]。本文以肺癌作為腫瘤模型,系統性研究銅離子濃度對細胞增殖的影響及其銅伴侶蛋白編碼基因在肺癌發展和預后中的意義。
1 材料和方法
1.1 材料
人肺癌細胞系A549和人正常肺上皮細胞系16HBE由國家癌癥中心/中國醫學科學院腫瘤醫院分子腫瘤學國家重點實驗室傳代培養,DMEM培養基、RPMI-1640培養基、0.25%胰酶及胎牛血清均購自美國Gibco公司,A/J小鼠購自上海南方模式生物科技股份有限公司,脫脂奶粉購自美國BD公司,抗細胞外調節蛋白激酶(extracellular regulated protein kinase,ERK)抗體、抗磷酸化ERK(phosphorylated ERK,p-ERK)抗體均購自美國CST公司,抗β-actin抗體購自美國Sigma公司,化學發光液購自大連美侖生物技術有限公司。
1.2 方法
1.2.1 細胞生長曲線實驗
將A549細胞和16HBE細胞接種于6孔板中,每孔0.8×105個細胞。待細胞貼壁完全后,實驗組用濃度為5 μmol/L的Cu2+溶液分別處理A549和16HBE細胞,對照組用等體積溶劑進行處理,分別于加藥后24、48和72 h,用0.25%的胰酶消化細胞,臺盼藍染色后計數,每個實驗重復3次后,用GraphPad Prism 7軟件繪制生長曲線圖。
1.2.2 蛋白樣品的制備及蛋白質印跡法(Western blot)實驗
將16HBE細胞接種于6孔板中,每孔2×105個細胞。待細胞貼壁后,將含不同濃度的銅離子溶液與細胞一起溫育48 h后,棄去培養基,用預冷的磷酸鹽緩沖液(phosphate-buffered saline,PBS)洗滌3次。然后將濃度為1×十二烷基硫酸鈉(sodium dodecyl sulphate,SDS)的上樣緩沖液(每3×105個細胞加入100 μL上樣緩沖液)加入6孔板中,用預冷的細胞刮將細胞刮下,并轉至Eppendorf試管中。將上述Eppendorf試管置于99 ℃水浴鍋煮12 min,每間隔3 min用渦旋儀振蕩混勻1次,得到的蛋白樣品用12 000 r/min離心1 min后取10 μL的蛋白量進行SDS聚丙烯酰胺凝膠電泳(SDS polyacrylamide gel electrophoresis,SDS-PAGE);濕轉至NC膜上,在電壓為110 V條件下轉膜2 h。取出NC膜并放入提前配置好的含5%脫脂奶粉的1×洗膜緩沖液(tris-buffered saline Tween,TBST)中,室溫放置1~2 h,用一抗稀釋液以1∶1 000比例稀釋抗p-ERK、抗ERK和抗β-actin抗體,將上述封閉后的NC膜放入目的抗體溶液中,4 ℃平板搖床溫育過夜,第2天用TBST洗膜3次,每次10 min;再放入稀釋好的帶辣根過氧化物酶(horseradish peroxidase,HRP)標記的二抗中,常溫溫育1~2 h后,用TBST洗膜3次,每次5 min,用化學發光成像儀顯影。
1.2.3 構建動物肺癌模型
4~5周齡雌性A/J小鼠在SPF級別動物房中飼養,對小鼠進行編號,待小鼠充分適應動物房無菌環境1周后開始給予煙草致癌物4-甲基亞硝銨-1-(3-吡啶基)-1-丁酮[4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone,NNK],來誘發小鼠產生肺癌。將A/J小鼠隨機分為對照組和NNK組,每組10只。通過灌胃方式給予NNK組小鼠100 mg/kg的NNK,1周兩次,連續5周。同時對照組小鼠給予相同體積的溶劑。在灌胃結束后,通過頸椎脫臼法處死小鼠,并將小鼠的肺組織取出,放置-80 ℃保存,以備后續實驗。本研究的所有動物實驗均按照中國醫學科學院腫瘤醫院動物管理與使用委員會批準的指南進行。
1.2.4 H-E染色
將40%的甲醛溶液固定、石蠟包埋的小鼠肺組織標本切成5 μm厚的切片,通過二甲苯脫蠟和不同濃度的乙醇對切片進行脫蠟處理。隨后用蘇木精染色5 min,用鹽酸乙醇分化后,用伊紅染液染色1~3 min;經過不同濃度梯度的乙醇與二甲苯徹底脫水透明后,用中性樹膠封蓋,在普通光鏡下觀察組織形態。
1.2.5 小鼠體內銅離子的檢測
對小鼠肺組織樣品進行稱重,取200 mg肺癌組織及對應癌旁正常組織,液氮冷凍狀態下研磨成粉末。將粉末在預冷的混合酸(HNO3∶HClO4=1∶1)中過夜,并轉移至5 mL去離子水中,隨后用電感耦合等離子體質譜儀(inductively-coupled plasma-mass spectrometry,ICP-MS)對銅離子濃度進行測定。
1.3 統計學處理
利用癌癥基因組圖譜(The Cancer Genome Atlas,TCGA)和Oncomine等數據庫分析銅伴侶蛋白超氧化物歧化酶(copper chaperone for superoxide dismutase,CCS)、細胞色素C氧化酶銅伴侶蛋白17(cytochrome C oxidase copper chaperone 17,COX17)和抗氧化劑1銅伴侶蛋白(antioxidant 1 copper chaperone,ATOX1)在患者中的表達情況,利用Kaplan-Meier Plotter網站分析上述基因表達水平與患者預后的關系。在GraphPad Prism 8軟件中采用雙尾Student’st檢驗對實驗結果進行數據分析。P<0.05為差異有統計學意義。
2 結果
2.1 銅離子促進細胞增殖
用濃度為5 μmol/L的銅離子溶液處理人肺癌細胞A549,同時用等體積溶劑處理對照組細胞,并于第1、2和3天檢測細胞的增殖情況。結果顯示,相較于對照組,用含銅離子的溶液處理細胞后,細胞數目明顯增加(P=0.038 6),說明銅離子能促進A549的增殖(圖1A)。在正常人肺上皮細胞來源的16HBE細胞中,濃度為5 μmol/L的銅離子也可促進細胞增殖,與對照組相比,差異有統計學意義(P=0.031 4,圖1B)。上述結果表明銅離子可促進細胞增殖。
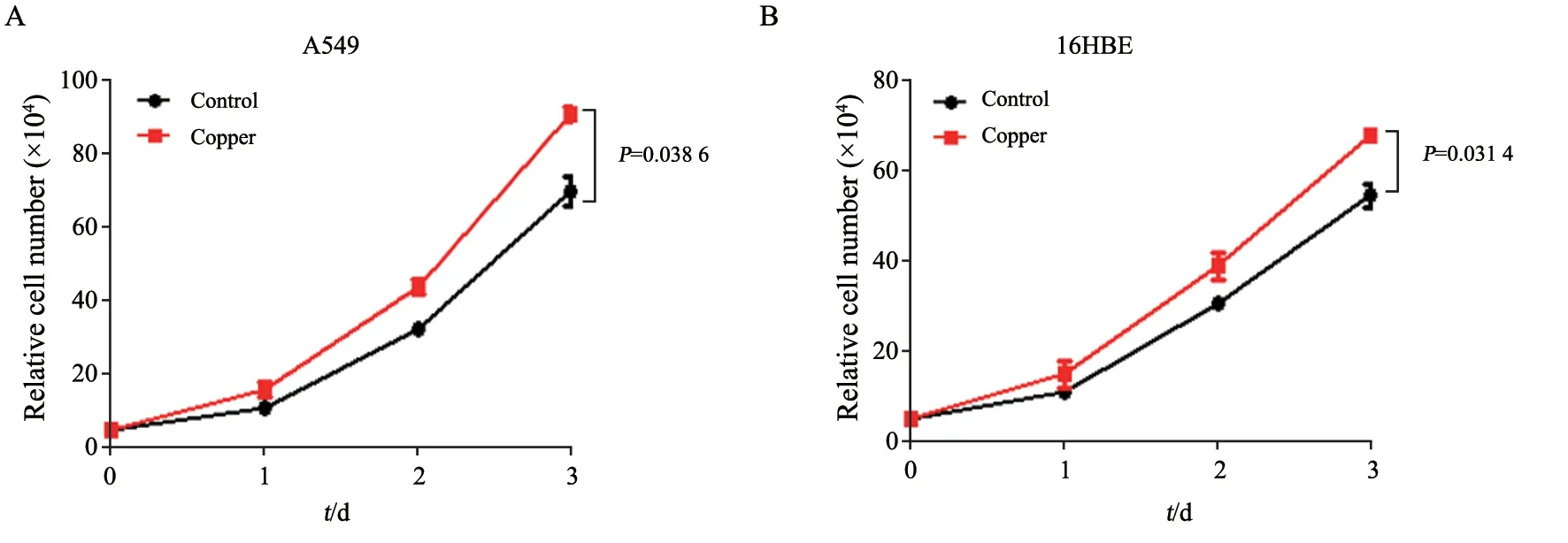
圖1 銅離子促進A549和16HBE細胞增殖Fig.1 Copper ions promote the proliferation of A549 and 16HBE cells
2.2 銅離子能提高細胞內ERK的磷酸化水平
ERK作為一類絲/蘇氨酸蛋白激酶,是傳遞絲裂原信號的信號轉導蛋白,也是多種生長因子的下游蛋白,能通過被磷酸化參與調節細胞的增殖、分化和存活等[16]。本研究分析了銅離子對細胞內ERK信號通路的影響,用濃度為2.5、5.0和10.0 μmol/L的銅離子溶液處理16HBE細胞,并于48 h后提取蛋白進行SDS-PAGE,通過Western blot檢測p-ERK、ERK蛋白的表達情況。結果顯示,隨著銅離子濃度的升高,p-ERK蛋白的表達量也隨之升高(圖2A、2B),說明銅離子能在一定程度上誘導細胞內ERK的磷酸化,從而促進細胞增殖。
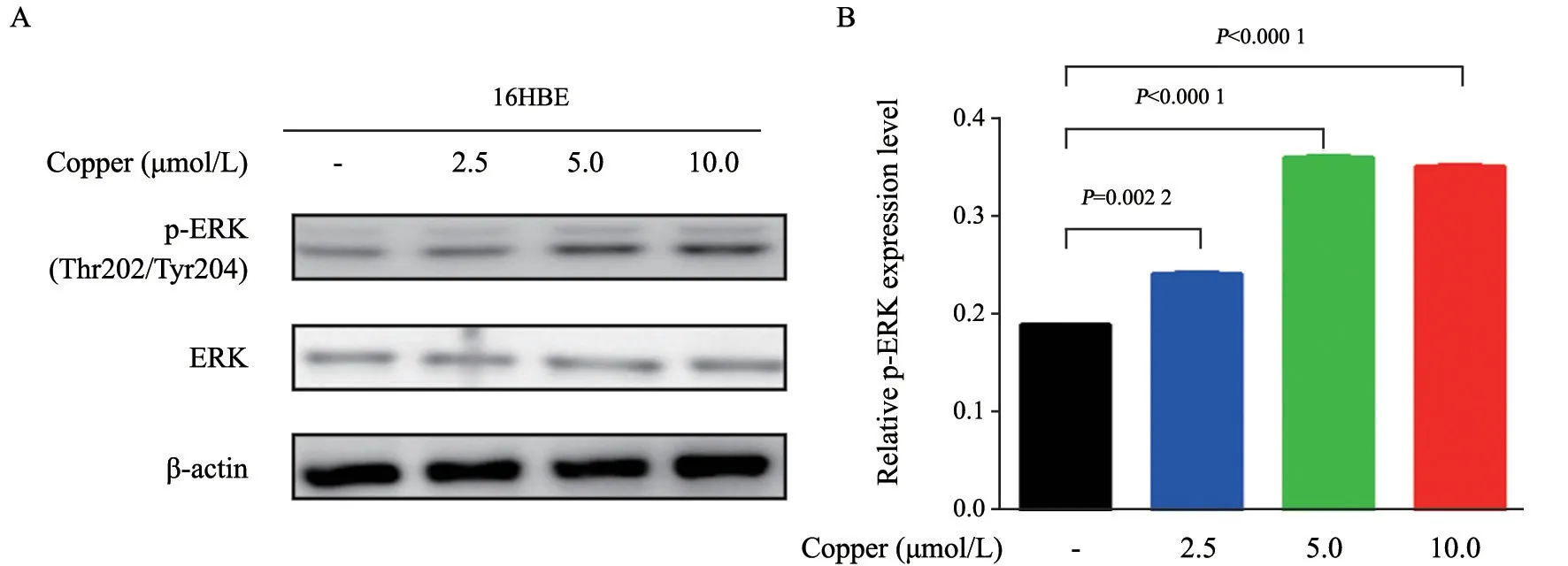
圖2 銅離子對16HBE細胞內ERK的影響Fig.2 The effects of copper ions on ERK in 16HBE cells
2.3 小鼠肺癌組織中銅離子的含量高于正常組織
將A/J小鼠在SPF級別的動物房中飼養并用耳號進行標記,隨機分為兩組。1周后通過對小鼠給予NNK建立小鼠肺癌模型。每周詳細觀察并記錄小鼠的體重和狀態,用小動物計算機體層顯像儀(small animal computed tomography,micro-CT)成像和對小鼠肺組織取材觀測小鼠肺部腫瘤生長情況,并通過H-E染色鑒定腫瘤類型。實驗結果顯示,NNK可以誘發小鼠肺癌的產生(圖3A)。進一步通過ICP-MS檢測小鼠肺癌組織和正常肺組織的銅離子含量,結果表明,小鼠肺癌組織中的銅離子含量高于正常肺組織(圖3B)。
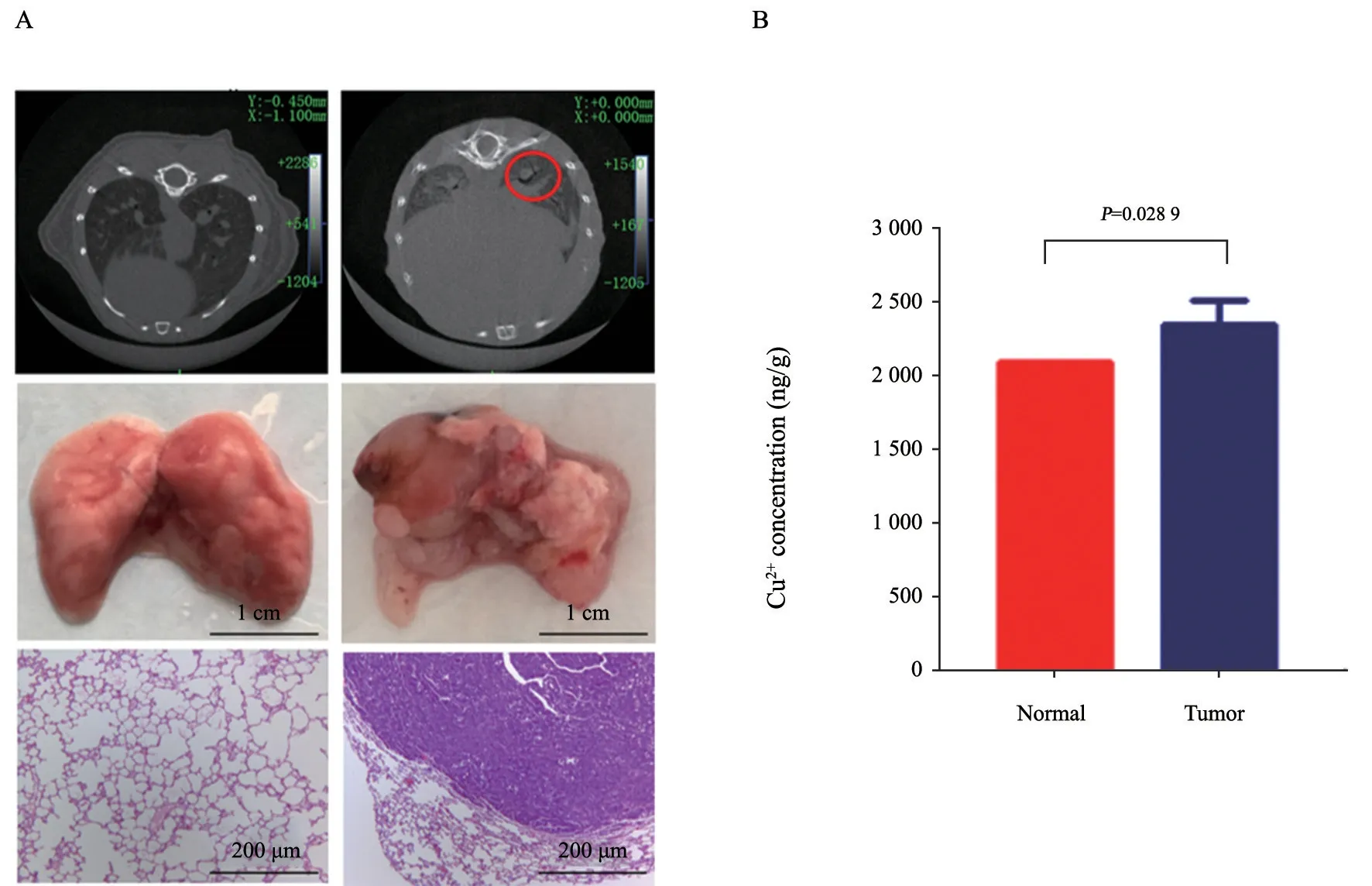
圖3 小鼠肺癌組織中的銅離子含量高于正常肺組織Fig.3 Copper ion concentration was higher in mouse lung cancer tissues than in normal lung tissues
2.4 銅伴侶蛋白編碼基因在肺癌組織中高表達
研究發現,銅離子進入細胞質后,主要通過CCS、COX17和ATOX1等,實現銅離子在不同細胞器中的運輸,進而發揮相應的生物學功能[17]。因此,本研究利用多種數據庫,進一步分析了銅伴侶蛋白編碼基因在肺癌患者癌組織和正常組織中的表達差異。通過對TCGA數據庫中肺癌患者數據進行分析發現,在肺腺癌患者中,肺癌組織中的CCS、COX17和ATOX1基因的mRNA表達水平顯著高于正常肺組織(圖4A~4C)。此外,通過對Oncomine基因芯片數據庫進行分析發現,在Garber、Stearman、Okayama、Su、Hou和Selamat數據中,肺癌組織中的CCS基因的mRNA表達水平高于癌旁組織(圖4D);在Garber、Okayama和Wachi數據中,肺癌組織中的COX17基因的mRNA表達水平也高于癌旁組織(圖4E);在Garber、Okayama、Hou和Landi數據中,肺癌組織中的ATOX1基因的mRNA表達水平也高于癌旁組織(圖4F)。
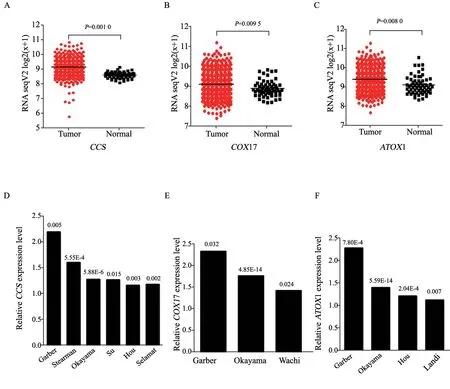
圖4 銅伴侶蛋白編碼基因CCS、COX17和ATOX1在肺癌患者癌組織中的表達水平顯著高于正常肺組織Fig.4 The expression levels of copper chaperone protein-encoding genes CCS,COX17 and ATOX1 were significantly higher in cancer tissues than in normal lung tissues
2.5 銅伴侶蛋白編碼基因與患者預后的關系
為進一步研究銅伴侶蛋白編碼基因CCS、COX17和ATOX1與肺癌患者預后的關系,利用Kaplan-Meier Plotter網站對3個銅伴侶蛋白編碼基因的表達水平與肺癌患者的預后進行了統計學分析。結果表明,在肺腺癌患者中,銅伴侶蛋白編碼基因CCS、COX17和ATOX1的表達水平與患者的預后及生存呈負相關(圖5),說明肺癌患者體內銅伴侶蛋白的表達水平可作為判斷患者臨床療效和評估預后的依據。
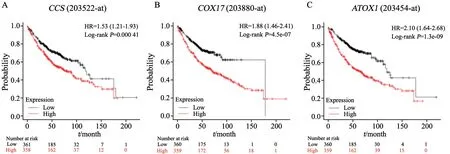
圖5 銅伴侶蛋白編碼基因的表達水平與肺腺癌患者的預后呈負相關Fig.5 The expression levels of copper chaperone protein-encoding genes were negatively correlated with prognosis of patients with lung adenocarcinoma
3 討 論
銅離子是人體生命活動所必需的一種重要微量元素。人體內銅離子的含量異常往往會引起一系列的疾病[18]。本研究發現,銅離子可以誘導細胞內ERK磷酸化,從而促進肺癌細胞和正常細胞的增殖。有研究[19]發現,培養羊的胰腺類器官時,銅離子能以ATOX1依賴的方式直接結合MEK,從而激活MEK-ERK信號轉導通路,而小鼠細胞中銅離子轉運蛋白1(copper transporter 1,CTR1)的缺失和突變都可降低MEK1磷酸化ERK的能力,說明MEK1-ERK相互作用需要銅離子的參與,而銅離子伴侶蛋白ATOX1和CTR1在銅離子與MEK的直接結合中均扮演重要角色。此外,二價銅離子可啟動兩種受體酪氨酸激酶(receptor tyrosine kinase,RTK)介導的下游信號轉導通路,包括AKT和ERK[20],說明銅離子還可能參與RTK的激活,從而導致ERK的磷酸化。
吸煙是引發肺癌的主要外因,吸煙者發生肺癌的風險是非吸煙者的20~30倍,且二手煙也會增加肺癌的發病風險[21],吸煙可引起各種類型的肺癌[22]。煙草燃燒產生的煙霧中有60多種已知的致癌物,包括多環芳烴類(如苯并芘)、苯、丙烯及亞硝胺類(如NNK)[23]。這些致癌物一般先與DNA形成加合物,再直接或間接誘使抑癌基因如TP53突變失活或癌基因如KRAS突變激活[23-25]。對亞硝胺類敏感的CYP2E1一般誘發肺腺癌[26]。在Cu2+和NADPH的存在下,亞硝胺的代謝物NNK和NNN能夠產生顯著水平的DNA氧化,從而增加癌癥發生的風險;同時,香煙煙霧中也含有大量的金屬離子,其中也包括銅離子;電子煙中的銅離子含量甚至可以達到普通香煙的6倍,這進一步解釋了肺癌組織中高濃度銅離子的來源[27]。
在機體正常組織中,銅離子主要通過銅伴侶蛋白來實現在不同細胞器中的運輸,進而發揮相應的生物學功能。目前,銅伴侶蛋白是腫瘤研究的熱點。本研究發現,CCS、COX17和ATOX1在人肺癌組織中的表達水平顯著高于正常組織,且這些蛋白的表達水平與肺癌患者的預后和生存呈負相關,這在一定程度上說明高濃度的銅離子在腫瘤的發生、發展中起著重要作用。銅離子濃度與腫瘤的惡性程度顯著相關,也預示著銅伴侶蛋白可作為判斷患者臨床療效和評估預后的一項依據。ATOX1和CCS7都是銅離子發揮生物學功能不可或缺的蛋白,且二者與銅離子相結合的結構區域有一定的相似性。抑制劑DC_AC50可以通過結合ATOX1和CCS的銅轉運界面,阻斷銅離子在細胞內的轉運,在裸鼠移植瘤模型中表現出顯著的抑瘤效果[28]。COX17與銅離子的作用方式則不同于ATOX1和CCS。COX17主要存在于細胞線粒體膜結構中,COX17與銅離子作用后可能通過調節腫瘤細胞的代謝從而影響腫瘤的發生、發展。也有研究[29-30]發現,在小鼠和人類細胞環境中,MEK1銅離子結合區的突變會降低BRAFV600E突變驅動的信號轉導和腫瘤發生,而銅離子螯合劑對這類突變型的腫瘤有著顯著療效,說明無論是通過螯合治療降低銅離子的濃度,還是通過抑制劑抑制銅離子的轉運,對腫瘤細胞都有著一定的抑制作用,減少銅離子對癌細胞內惡性通路的激活是十分有潛力的治療方向。然而,由于銅離子本身是一種重要的必需微量元素,而目前大部分銅配體藥物均是采取全身給藥方式,不加甄別地實現全身銅耗竭,因此往往會導致一些不良反應,例如影響銅離子與血管生成素的結合及新生血管生成等,最終導致紅斑、視神經炎、嘔吐和白細胞減少等不良反應[31]。因此,如何在開發靶向銅離子藥物及給藥系統的同時避免影響其正常的生理功能,將是未來研究的重點。
本研究發現,高濃度銅離子能誘導ERK磷酸化,并促進腫瘤細胞增殖,并探索了銅離子在腫瘤組織與正常組織中的濃度差異。ERK信號轉導通路的激活除了與細胞的增殖和生存相關外,還可誘導機體產生炎癥反應[32-33]。因此,在今后的研究中,可以結合其可能的對人體免疫調節作用,綜合研究銅離子促進腫瘤發生、發展的機制。此外,本研究表明,銅伴侶蛋白編碼基因表達水平與肺癌患者預后呈負相關,因此可以通過抑制銅伴侶蛋白的作用,減弱腫瘤細胞對銅離子的過量攝取和阻斷銅離子參與的腫瘤發生、發展過程,從而為腫瘤治療提供新的思路。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
癌變·畸變·突變(2016年3期)2016-02-27 06:15:34
醫學研究雜志(2015年12期)2015-06-10 06:57:46
鄭州大學學報(醫學版)(2015年1期)2015-02-27 14:50:26
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
