次生林不同植被類型下各個徑階競爭強度探析
2022-01-17 14:08:16張遠榮胡明星
黑龍江生態工程職業學院學報 2022年1期
關鍵詞:研究
張遠榮 胡明星
(云南融創林業規劃設計有限公司,云南 昆明 650000)
隨著國家儲備林培育的開展,林木生長變得尤為重要。在林木生產的過程中,林分中的林木競爭關系是直接制約林木生長的一個重要因素[1]。在生態學研究中,優勢樹種在林分中的競爭強度極強,具有比較明顯的地位,優勢樹種的變化表現為一個種群的變化,因而對一個種群變化的研究也就變得較為重要。種群的研究主要包括種群的分布格局、種群的年齡結構及種群的增長模型。種群空間分布格局分為均勻型空間分布、隨機型空間分布和集群型空間分布3種類型。
林分中的種群特性決定了種群在林分空間中的分布結構。以單株林木作為一個具體的坐標點向周邊輻射,從而研究林分的空間結構特征,這方面的研究取得了較多的研究成果[2-5]。而對于競爭關系的研究也有研究人員以相鄰的五株林木作為一個單元進行研究,以中間的作為目標樹,周邊四株作為競爭木[6]。通過這樣的研究可以直觀地得出一株樹的基本空間研究要素:角尺度、混交度及大小比數[7-9],從而可以推出林木在空間中的競爭關系。
在林分空間競爭指數的研究中,研究林木的通常有CI1指數至CI6指數。其中CI1針對胸徑的變化研究,CI2針對樹高的變化研究,CI3針對平均冠幅的變化研究,而CI4、CI5和CI6則是分別以CI1、CI2、CI3的平方形式強調競爭木與對象木的不對稱性。各競爭指數的計算公式如下:
(1)
(2)
(3)
(4)
(5)
(6)
以上公式(1)至(6)中:DBH為胸徑(或直徑)(cm);H為樹高(m);cpa為平均冠幅(m);t為第t株對象木;j為第j株競爭木;distj為林分樣地中的對象木與競爭木之間的直線距離;n為第n株對象木的競爭木株數。
1 空間結構的研究方法
林木生長直接受限于空間結構的分布情況,要想有限的林地資源獲取最快的生長速度,獲得最大的生物量,必然要減少林木間的競爭,減少林分間的競爭。之前比較注重群落的研究,較少關注同一林分之間不同胸徑上的差異化。
1.1 確定空間結構單元
根據競爭的基本原理,空間單元確定為一目標樹四競爭木的法則作為單元開展研究。但即便是這樣,在實際操作中也比較難以量化地表示其中的相對位置關系,并且這最多也只能得到3個或4個方向上的空間關系,其他方位的空間信息是不完整的。判斷方向用指北針等輔助工具,在野外調查時,受地理位置的影響,本身就有困難,而當超過4個方位時則會更困難一些;并且要嚴格地定位樣地中每一株林木的具體坐標,減少相對誤差,這在很大程度上直接增加了測算的難度。

表1 結構參數取值及含義
除此方法之外,還有一種比較常用的方法是以參照樹為圓心,以一定的距離作為競爭半徑。但是由于立地質量、環境、海拔等都會導致競爭半徑發生較大的變化,不穩定因素比較多,所以很多時候在采用此種方法時都要先進行競爭距離確定,從而也導致調查任務的工作量加大。
1.2 林分空間結構參數的計算
本研究則將調查中心區域中每一株林木分別作為對象木,以其為中心,周圍臨近的四株林木為競爭木開展計算。中心調查樣地設置為30m×50m,競爭木的范圍在30m×50m的基礎上,分別在四周拓寬10m,形成50m×70m的環形樣方。最終得到的數據利用Arcgis的泰森多邊形等技術手段得出空間結構參數。從而計算并確定各個林分中的角尺度、混交度和大小比數等空間競爭數據。
2 研究方法
2.1 樣地的選取
研究選取6種典型林分建立16塊樣地,分別是:熱帶季雨林群落、熱帶雨林群落、山地雨林群落、熱性竹林群落、季風常綠闊葉林群落和落葉闊葉林群落。
2.2 標準地因子測定
(1)區劃方格:以10m作為一個標尺,分出內部核心15塊方格,外部競爭20塊方格,逐地塊逐株調查。
(2)每木檢尺。
2.3 競爭強度計算
CI1競爭指數的計算公式:
式中:DBH為林木胸徑(cm);t為第t株對象木;j為第j株競爭木;distj為林分樣地中的對象木與競爭木之間的直線距離;n為第n棵株對象木的競爭木株數。
3 研究結果
林分中林木的生長競爭受限于很多因素,宏觀上說,有氣候、海拔、季節等等;微觀上說,有樹種類別、植株胸徑、樹高、林木所處的生長階段、個體生產力的多少、相鄰的植物(年齡、大小、生態位、生產力等)對其的影響,等等。從圖1中可以看出,在林木的生長過程中,競爭指數隨著對象木的徑階變化而發生變化,并呈現出一定的規律性。一般而言,林分中的徑階為4cm的對象木所受的競爭最大。整體而言,競爭指數隨著徑級的增大而逐漸減小。結果如圖1所示,圖中小圓圈表示在同一樣地中不同的樹木其競爭強度的大小。
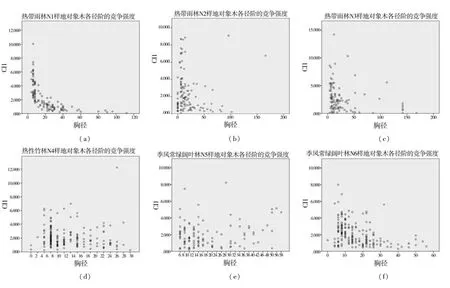
注:(a)(b)(o)表示的是熱帶雨林對象木各徑階的競爭強度;(c)(m)(n)表示的是熱帶季雨林對象木各徑階的競爭強度;(e)(f)(j)表示的是季風常綠闊葉林對象木各徑階的競爭強度;(d)表示的是熱性竹林對象木各徑階的競爭強度;(g)(k)表示的是落葉闊葉林對象木各徑階的競爭強度;(h)(l)表示的是山地雨林對象木各徑階的競爭強度。
由圖1 (a)(b)(o)可以看出:在熱帶雨林中,當林木胸徑小于20cm時,隨著徑階的增大競爭強度急劇下降,其原因主要是由于受到上層林冠對光照的阻擋以及四周樹木的擠壓,中小徑階的林木對于空間和資源的爭奪競爭十分激烈;而當對象木的胸徑超過20cm時,競爭強度趨勢變得平緩,各徑階的競爭強度波動較小,且隨著胸徑的變大,各徑階的競爭強度趨于平穩不變。說明當喬木生長到一定徑階,即相比周圍競爭木有著較大胸徑時,其樹冠基本上已經定型,周圍的競爭木對其影響較小,此時對象木受到的遮擋作用較弱,競爭強度較小,故而大徑階喬木在各徑階的競爭強度中呈現平緩、波幅變化小的勢態。
由圖1(c)(m)(n)可以看出:在熱帶季雨林中,當林木胸徑不足16cm時,各徑階的競爭強度急劇下降,而當對象木的胸徑超過16cm時,下降趨勢變得平緩,各徑階的競爭強度波動較小,且隨著胸徑的變大,各徑階的競爭強度趨于平穩不變。
由圖1(e)(f)(j)可以看出:在季風常綠闊葉林中,林木各個胸徑所對應的下降趨勢整體而言都較為平緩,各徑階的競爭強度波動較小,且隨著胸徑的變大,各徑階的競爭強度趨于平穩不變;但是胸徑從小到大的過程中,多會出現有零散的喬木的競爭強度高于整體水平,其原因主要是季風常綠闊葉林中樹木更新相對于其他林分較好,各個徑階的林木處于一種較為穩定的更新,從而各個徑階的競爭強度不會出現急劇下降的現象。
由圖1(d)可以看出:在熱性竹林中,林木各個胸徑所對應的下降趨勢整體而言都較為平緩,各徑階的競爭強度波動較小,且隨著胸徑的變大,各徑階的競爭強度趨于平穩不變;但是胸徑從小到大的過程中,多會出現有零散的喬木的競爭強度高于整體水平,其原因主要是熱性竹林中樹木更新相對于其他林分較好,各個徑階的林木處于一種較為穩定的更新,從而各個徑階的競爭強度不會出現急劇下降的現象。
由圖1(g)(k)可以看出:在落葉闊葉林中,當林木胸徑不足20cm時,各徑階的競爭強度急劇下降,而當對象木的胸徑超過20cm時,下降趨勢變得平緩,各徑階的競爭強度波動較小,且隨著胸徑的變大,各徑階的競爭強度趨于平穩不變。
由圖1(h)(l)可以看出:在山地雨林中,林木各個胸徑所對應的下降趨勢整體而言都較為平緩,各徑階的競爭強度波動較小,且隨著胸徑的變大,各徑階的競爭強度趨于平穩不變;但是胸徑從小到大的過程中,多會出現有零散的喬木的競爭強度高于整體水平,其原因主要是山地雨林中樹木更新相對于其他林分較好,各個徑階的林木處于一種較為穩定的更新,從而各個徑階的競爭強度不會出現急劇下降的現象。
次生林主要林分中的優勢樹種競爭指數不一,這主要是由于各個林分的海拔、地理位置以及各個林分的樹種組成、年齡結構、林分密度的不同等因素而導致。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
