附睪精子抗氧化保護機制研究進展
2022-02-15 10:46:20許嬌霞張家新
中國畜牧獸醫 2022年1期
關鍵詞:氧化應激
許嬌霞,張家新
(內蒙古農業大學動物科學學院,內蒙古自治區動物遺傳育種與繁殖重點實驗室,呼和浩特 010018)
精子在睪丸形成后功能還未成熟,在附睪轉運過程中才逐漸獲得受精能力和運動能力[1],因此,附睪是精子最后成熟的場所[2]。精子在附睪轉運的同時,附睪將含有與精子成熟相關蛋白的附睪小體分泌到附睪液中并作用于精子。精子在經歷一系列的表觀修飾后,染色質、質膜都將發生重塑,使得精子逐漸成熟[3]。且附睪上皮分泌的附睪小體中含有多種抗氧化酶可傳遞給精子,保護其免受氧化損傷[4-5]。
1 附睪精子面臨的氧化應激
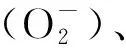
精子氧化損傷主要表現為脂質過氧化增加和DNA損傷[7]。由圖1可知,精子代謝異常時線粒體產生過量的ROS進入細胞核后,可將鳥苷氧化成8-羥基脫氧鳥苷(8-OHdG),從而引發DNA突變;而當細胞外ROS過量時脂質過氧化增加,其產物羥基壬烯醛(4-HNE)相應上升后對DNA的突變也具有誘導作用[8-11]。Wu等[2]為檢測氧化應激對附睪精子的影響作用,對成年大鼠進行腹腔注射叔丁基過氧化氫(t-BHP)處理,并收集其第3、6和9周附睪尾部的精子,發現3個時期的精子都呈低運動性和高DNA氧化水平,而且t-BHP造成的氧化應激會對附睪的功能造成長期影響,在大鼠經t-BHP處理后第9周,其附睪的脂質過氧化依舊處于一個較高的水平,從而影響到精子成熟,導致精子質量較低。據報道,人的精子在受到氧化應激后,其功能和活力受損,進而對精子的受精過程造成影響,嚴重時則會導致男性不育[3-14]。而適量的ROS不僅可以保護精子免受氧化損傷,還對精子的獲能具有促進作用[10-11],可見附睪中正常生理水平的ROS對雄性生殖能力至關重要。
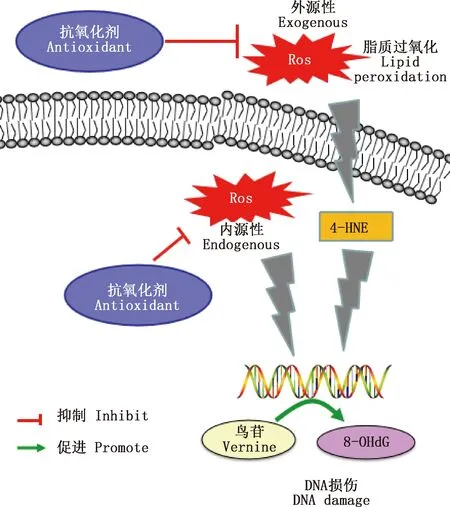
圖1 精子中活性氧的作用通路圖[10-11]Fig.1 Pathway diagram of reactive oxygen species in sperm[10-11]
隨著精子在附睪內的運行與成熟,其代謝率、形態、運動模式都會發生改變,使附睪不同部位的精子產生了不同水平的ROS,而此時精子對于抗氧化劑的需求也會隨之發生改變,以適應不同產量的ROS[15]。Rana等[16]報道,在山羊附睪不同節段的精子和附睪液中可以檢測到多種抗氧化酶,附睪液中抗氧化酶的活性從附睪頭到附睪尾逐漸增加,而精子中的趨勢卻與之相反,這說明山羊附睪液為精子提供的抗氧化環境對精子具有相應的補償作用。因此,附睪中生理水平ROS的維持需要各種抗氧化酶的相互協調,從而起到保護精子使之正常成熟的作用[5]。
2 附睪中的抗氧化酶
在附睪中,將ROS維持在生理水平的抗氧化酶主要有超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、谷胱甘肽過氧化物酶(GPXs)以及過氧化物酶(PRDXs)等,各種抗氧化酶清除的ROS類型以及產物見圖2。以下將對抗氧化物酶在附睪中的作用進行闡述。
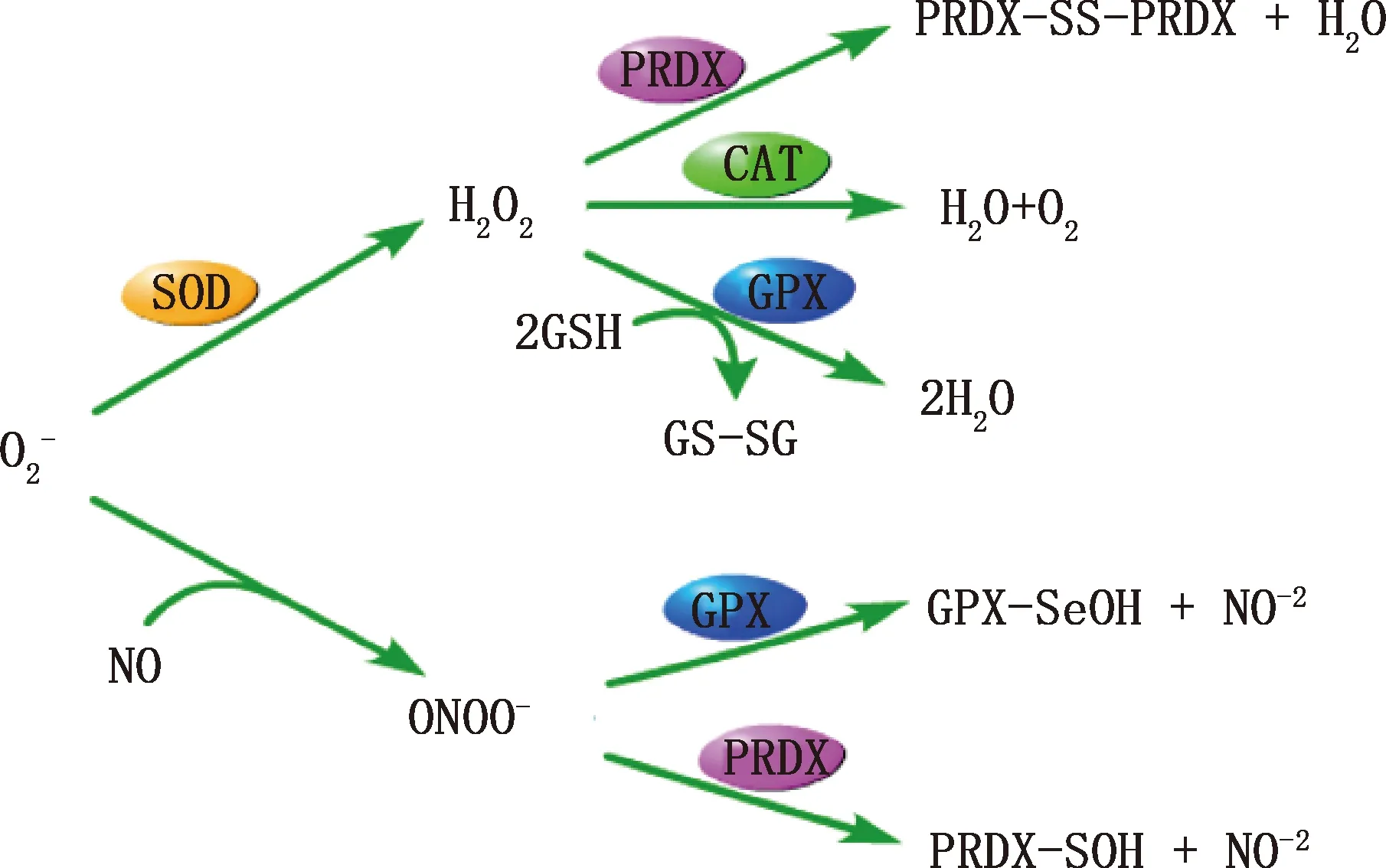
圖2 重要抗氧化劑的作用通路[17]Fig.2 Pathway diagram of important antioxidants[17]
2.1 超氧化物歧化酶

有報道表明,SOD在維持精子活力以及功能方面具有重要作用,Aitken等[19]對高活力精子與異常精子SOD活性檢測后發現,SOD活性與氧化損傷程度呈正相關,而且在大鼠附睪中SOD的活性還會隨年齡的增長而降低[20]。另外,一些精子DNA完整性較差的患者可通過口服SOD抗氧化劑使其癥狀得以改善[21]。由此可見,SOD對精子DNA完整性具有重要的保護作用。
2.2 過氧化氫酶
CAT是一種含血紅素的酶,通過使用鐵(Fe)或錳(Mn)輔助因子,將H2O2轉化為H2O和O2。在大多數真核細胞中,CAT在過氧化物酶體中的濃度最高,在線粒體和胞漿中濃度較低。Jemec等[22]認為,CAT在某種程度上起輔助GPXs的作用。通過對人紅細胞使用放射性標記和氨基三唑抑制后發現,CAT可清除50%的H2O2,而GPXs和谷胱甘肽還原酶(GSH)則負責清除另一半[23]。
然而,CAT在兔[24]、小鼠[25]、牛[26]、水牛[27]、豬[28]的精子中并不存在,但在人[29]、大鼠[30]精子中有較低水平表達,Jean-fran?ois等[26]和Kar等[27]還發現CAT在牛精液中含量很高。Kumar等[31]在山羊附睪不同部位的附睪液和精子中也未檢測到CAT的活性。表明不同物種的精子與附睪中CAT的作用機制存在較大差異。此外,Wu等[12]通過使用t-BHP處理大鼠附睪以驗證CAT對附睪中ROS的清除能力發現,大鼠附睪任何部位中的CAT表達水平均不會隨t-BHP的處理而改變。說明CAT似乎并不是大鼠附睪中抵御氧化應激的關鍵酶。但對于其他動物是否也如此仍需進一步的研究。
2.3 谷胱甘肽過氧化物酶
GPXs可將脂質氫過氧化物(ROOH)還原為相應的醇,并將H2O2轉化為H2O。到目前為止,在哺乳動物中已經報道了8種GPXs(GPX1-8)的基因亞型,這些基因在哺乳動物不同器官中廣泛分布[32-33]。有報道表明,當給雄性大鼠服用谷胱甘肽合成酶抑制劑(BSO)后,附睪尾精子抗氧化能力下降,繼而使染色質質量受損[34]。這些研究表明GPXs在精子成熟過程中的保護作用至關重要[2,5]。
在GPXs家族中,GPX1-4的活性依賴于硒,其中GPX4是一種重要的結構蛋白[35],在精子線粒體中含量非常豐富(約占精子中段蛋白的50%),而當其缺失時則會導致男性不育[36]。GPX4的核形式(nGPX4)有助于維持精子染色質的穩定性,但敲除nGPX4基因的雄性小鼠仍具備生育能力[37]。GPXs家族的另一成員GPX5,其活性不依賴于硒[32],由附睪上皮的主細胞分泌,與位于細胞質中GPX1、GPX3和GPX4的低表達量相反[38],GPX5在附睪頭中高度表達,而且受雄激素的調控[39],可以保護精子DNA免受氧化損傷[33]。敲除GPX5的雄性成年小鼠(3個月~1歲),其精子的DNA氧化水平高于野生型對照,敲除GPX5后雖然具備生育能力并可以產生正常的后代,但隨著這些雄性小鼠年齡的增長,其幼崽中流產、圍產期死亡及具有不同程度發育缺陷的異常生殖現象都有所增加[32-33]。這表明附睪中GPX5對于精子的抗氧化保護作用具有重要意義。Vilagran等[40]發現,豬新鮮精液中的精子質量較低時,機體會通過升高精漿中GPX5的濃度發起補償機制來保護精子,且精漿中GPX5的含量對人工授精母豬的產仔數有積極影響[41]。Williams等[42]發現,GPX5能夠防止精子過早地發生頂體反應。作者所在研究團隊在對綿羊附睪抗氧化功能的研究中發現,在綿羊正常生理條件下,不論是在附睪不同部位的組織、管腔液還是精子中,從附睪頭至附睪尾,GPX5的表達量均呈下降趨勢,但隨著精子在附睪的轉運,精子上GPX5的分布卻越來越廣泛,儲存至附睪尾部的成熟精子在核后帽、頸部、線粒體鞘、以及精子尾部均有GPX5的分布(數據未發表)。此外,大鼠精子的免疫熒光結果顯示,精漿中GPX5在附睪轉運和射精過程中結合到精子頭部[42]。總之,在哺乳動物各階段,精子抵御氧化損傷時,GPXs都起著重要的核心作用[5,15,33]。
2.4 過氧化物酶
PRDXs是一個抗氧化酶家族,從酵母到人類都高度表達。其活性位點含有1或2個半胱氨酸(Cys)殘基,在氧化還原過程中參與分子二硫鍵交聯,這對它們的抗氧化功能至關重要[43]。根據活性位點上Cys殘基的數量,PRDXs亞型分為2-Cys PRDXs (PRDX1-5)和1-Cys PRDX (PRDX6)[44],它們的過氧化物酶活性受其活性部位Cys殘基的氧化狀態調節。PRDXs在附睪頭和附睪尾的主細胞、基細胞以及窄細胞中高表達,但在透明細胞中不表達[17]。這些酶是精子中重要的抗氧化劑,可以降低過氧化物(H2O2和有機氫過氧化物)和ONOO-等ROS的水平,以避免細胞受到氧化損傷[45-46]。當活性中心Cys被氧化(硫醇氧化)變得無活性時[44],如果ROS水平進一步增加,PRDXs就會被過度氧化,并形成含有PRDX-SO2的高分子復合物[47]。因此,PRDXs須還原Cys殘基才能繼續清除ROS。 其中,PRDX1-5的氧化形式可以通過硫氧還蛋白(TRX)/硫氧還蛋白還原酶(TRD)/NADPH系統恢復到其活性狀態,PRDX1-5的過氧化形式則可以通過硫氧還原蛋白(SRX)或sestrins用ATP和GSH還原(圖3A)。而在PRDX6被氧化為PRDX6-SOH的情況下,谷胱甘肽巰基轉移酶(GSTpi)和GSH可以通過還原其活性位點的氧化硫醇來恢復其活性。然而,PRDX6的過氧化形式PRDX6-SO2卻不能被再次還原,將永久失活不可逆[48](圖3B)。
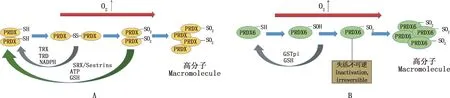
圖3 PRDX1-5(A)和PRDX6(B)的活化機制[17]Fig.3 Activation mechanism of PRDX1-5 (A) and PRDX6(B)[17]
在PRDXs家族中,PRDX6是被研究較多的一個酶,它是一種具有過氧化物酶和鈣非依賴性磷脂酶A2(iPLA2)活性的雙重酶,可防止脂質過氧化和修復氧化膜[49]。基于PRDX6基因敲除動物模型的研究表明,在附睪中缺乏PRDX6的精子出現異常成熟的跡象[48],且與野生型對照相比,產仔數和窩產仔數均降低[50]。這樣的低生育能力與缺乏PRDX6所導致的精子活力受損、DNA氧化、精蛋白水平降低以及DNA致密化有關[5,47],而且這種現象會隨著年齡的增長而加劇[51]。Liu等[52]通過給大鼠腹腔注射t-BHP來模擬氧化應激,2周后收集附睪組織和精子發現,經處理后的精子PRDX1和PRDX6水平升高,由于精子不能合成蛋白質,因此推測精子中PRDXs含量增加是由附睪上皮分泌含有PRDXs的附睪小體轉移到了精子上引起的。另外,有研究發現,附睪上皮在對抗氧化應激時,除了大量表達PRDX1和PRDX6外,同時還會通過直接分泌的方式進入附睪液中保護精子抵抗氧化應激[48]。這些研究表明附睪中PRDXs的抗氧化作用對于附睪成熟過程中的精子具有重要的保護作用。
3 小 結
精子在附睪轉運過程中發生的一系列變化,增加了其對高水平ROS的敏感性,而當發生氧化應激時,則會導致精子脂質過氧化增加和DNA的損傷,因此附睪中各抗氧化酶家族對成熟過程中的精子具有重要意義。目前,附睪中各類抗氧化酶家族如何協同發揮作用以將ROS維持在生理水平并不清楚,這仍需進一步研究,以闡明附睪精子的抗氧化保護機制。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
