不同窖齡及位置窖泥微生物群落和代謝組分的差異
2022-02-22 11:35:34畢天然黃鈞張宿義陳曉茹陳蘇祺母雨蔡曉波邱川峰周榮清
食品與發酵工業 2022年2期
關鍵詞:差異
畢天然,黃鈞,張宿義,陳曉茹,陳蘇祺,母雨,蔡曉波,邱川峰,周榮清*
1(四川大學 輕工科學與工程學院,四川 成都,610065)2(瀘州老窖股份有限公司,四川 瀘州,646000)
濃香型白酒是復雜的微生物種屬協同作用的結果。在其漫長的重復生產過程中,環境脅迫作用驅動窖泥微生物群落的定向進化是生產高質量基酒的必要條件[1],尤其是發酵后期,窖泥中棲息的功能菌群對獨特風味的形成具有重要作用[2]。已完成的研究結果表明,窖池中所棲息的Clostridiaceae、Ruminococcaceae、Methanosaeta、Methanobacterium、Methanocorpusculum等功能菌群隨著窖齡的增長而增多,Lactobacillus則降低,小生境中氧、酸及乙醇脅迫作用是驅動其變化的主要因素[3-5]。窖池的不同位置窖泥菌群多樣性差異顯著,DING等[6]在窖壁泥中檢出Acinetobacter,而窖底泥未檢出,WANG等[7]報道Lactobacillus豐度隨窖池深度增加而減小,而Clostridium與之相反。近年來高通量測序技術已成為研究群落的重要工具之一,廣泛應用于白酒的研究[5, 8]。熒光原位雜交(fluorescence in situ hybridization,FISH)技術是細胞水平表征活細菌群落結構輪廓的有效手段之一,曾與PCR-DGGE組合,從分子和細胞水平表征濃香型白酒釀造過程的群落特征[9]。
本文基于Illumina Miseq測序等多相技術,探尋不同窖齡窖池窖壁和窖底泥微生物群落及主要代謝組分的差異,應用生物信息學方法解析了群落中種屬間互作關系及其與主要代謝組分的相關性。旨在多尺度研究窖泥中核心功能菌群的代謝調控規律,為詮釋濃香白酒釀造過程微生態理論奠定基礎,為優化生產過程提供技術支撐。
1 材料與方法
1.1 樣品收集
樣品:窖泥樣取自瀘州老窖股份有限公司(四川瀘州)的30、100、200年窖齡窖池。參考DING等[3]所述方法分別從窖壁和窖底采樣,每樣取200 g,無菌PE袋密封后保存于-20 ℃冰箱。30、100、200年窖池的窖壁(JB)和窖底(JD)泥的編號分別是JB30、JD30、JB100、JD100、JB200和JD200。
1.2 主要理化性質檢測
pH測量:參考HU等[8]所述方法,準確稱取2.00 g窖泥,加6 mL超純水,渦旋振蕩5 min,超聲30 min,離心(12 000 r/min,4 ℃)10 min,上清液用于pH測量。
總酸和總酯測定:準確稱取10.00 g窖泥,加入到100 mL 30%乙醇溶液中,渦旋振蕩5 min,超聲30 min,4 ℃過夜浸提,濾紙過濾,濾清液按照GB/T 10345—2007所述的方法測定總酸和總酯。
1.3 代謝組分分析
有機酸測定:準確稱取5.00 g窖泥, 加20 mL 9 mmol/L H2SO4溶液,超聲60 min,其間每隔15 min渦旋振蕩5 min,離心(12 000 r/min,4 ℃)10 min收集上清液,上清液經活化后的C18SPE小柱純化,再經0.22 μm濾膜過濾,濾液參考DUBER等[10]所述HPLC法檢測有機酸含量。以標準品乳酸、乙酸、丙酸、丁酸和己酸的保留時間確定樣品中有機酸種類,并通過外標法計算含量。
色譜條件:色譜柱為Alltech OA-1000(300 mm×7.8 mm)有機酸柱,流動相為9 mmol/L H2SO4溶液,流速0.3 mL/min,紫外檢測器,檢測波長210 nm,柱溫75 ℃,進樣量10 μL,Agilent 1260高效液相色譜儀(Agilent Technology,美國)。
揮發性組分測定:參考DING等[11]所述的頂空固相微萃取-氣相色譜-質譜聯用法(headspace solid phase microextraction combined with gas chromatography-mass spectrometry,HS-SPME-GC-MS)測定樣品的揮發性組分。準確稱取0.50 g窖泥于頂空瓶中,加入10 μL 5.1 mg/L辛酸甲酯內標物質,60 ℃水浴中平衡15 min、50/30 μm DVB/CAR/PDMS纖維萃取頭吸附50 min,取出后插入進樣口,250 ℃解離5 min進行GC-MS分析。樣品質譜數據與標準譜庫(NIST2005)比對,匹配度>800(最大為 1 000)的物質予以報道,采用內標法對樣品中各檢出物質進行半定量。
色譜-質譜條件:色譜柱為HP-INNOWAX毛細管色譜柱(30.0 m×0.32 mm×0.25 um, Agilent, USA);升溫程序:40 ℃保持5 min,然后以 4 ℃/min 升至 100 ℃,再以6 ℃/min升至 230 ℃,保持10 min,進樣口溫度250 ℃;高純氦作為載氣,流速 1.0 mL/min;離子源溫度和連接線溫度分別為230、250 ℃;EI 電子能量為70 eV;質譜掃描范圍為 35~400 amu,Trace GC Ultra-DSQ II氣相色譜-單四級桿質譜聯用儀(Thermo Fisher Electron,美國)。
1.4 高通量測序檢測微生物群落組成
按照HE等[12]所述步驟,采用Fast DNA SPIN試劑盒(MP Biomedicals,Santa Ana,CA,USA)提取樣品DNA,NanoDrop ND-1000光度計(Thermo Fisher Scientific,Waltham,MA,USA)和1%瓊脂糖電泳檢測提取的DNA含量、純度和完整性。338F/806R和A RC787F/ARC1059R分別用于細菌和古菌16S rDNA 基因V3~V4高變區和ARC區的擴增,而ITS5/ITS1擴增真菌ITS1區。PCR流程參考LI等[13],PCR產物切膠獲得目的片段并純化回收,分光光度計測定后,等分子質量混合。通過試劑盒MiSeq ReagentKit v3構建文庫后,送上海派森諾生物科技有限公司Illumina MiSeq平臺測序。
測序數據刪除堿基平均質量≤Q20及模糊序列,FLASH 軟件(V1.2.7,http://ccb.jbu.edu/software/FLASH)拼接初步篩選的雙端序列(堿基重疊長度>10 bp,且無堿基錯配),將拼接后的序列分配到相應的樣本中,獲得有效序列。QIIME 軟件(Quantitative Insights Into Microbial Ecology,v1.8.0,http://qiime.org/)刪除低質量序列(長度≤150 bp,5′端堿基錯配>1和相同堿基>8的序列)獲得高質量序列。使用UCLUST把高質量序列按照97%的序列相似度聚成不同的可操作分類單元(operational taxonomic units,OTU)。分別采用Greengenes數據庫(Releasese 13.8,http://greengenes secondgenome.com/)和 UNIT 數據庫(Release 5.0,http://unite.ut.ee/)作為OTU分類鑒定的模板序列,獲得每個OTU的微生物分類信息。基于每個 OTU在各樣本中所含序列數構建OTU矩陣,確定微生物在各樣品中的相對豐度。
1.5 熒光原位雜交檢測群落組成輪廓
準確稱取1.00 g窖泥,加25 mL 10 mmol/L PBS(pH 7.2)緩沖液,渦旋振蕩5 min,離心(800 r/min,4 ℃)10 min收集上清液,上清液離心(12 000 r/min,4 ℃)10 min收集沉淀,相同緩沖液重復3次,獲得的菌體沉淀為FISH檢測樣品,參照何翠容等[14]所述條件與步驟進行FISH檢測,靶向探針見表1。
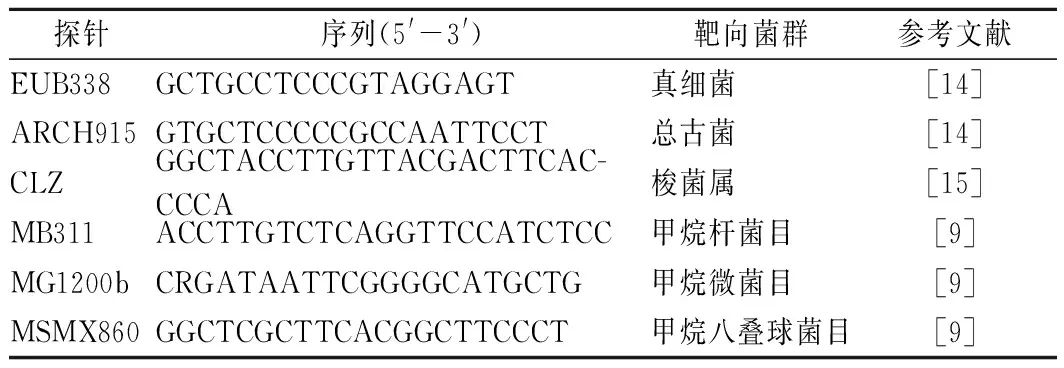
表1 熒光原位雜交靶向探針Table 1 Target probes of fluorescence in situ hybridization
1.6 數據處理
熱圖分析、非加權組平均聚類分析(unweighted pair-group method with arithmetic means, UPMGA)和冗余分析(redundancy analysis,RDA)使用R語言軟件(R x64 3.6.1,https://r-project.org/)完成。SPSS 19.0軟件(SPSS Inc.Chicago,IL,USA)用于Spearman′s相關系數分析和單因素顯著性分析,并根據Spearman′s相關系數繪制群落關聯網絡圖。每組檢測重復3次,所得數值結果以平均值±標準偏差表征。
2 結果與分析
2.1 窖齡及位置對窖泥理化和主要代謝組分的影響
如表2所示,不同窖齡窖底泥pH和總酸差異不大,總酯的含量則是JD200>JD30>JD100。以不銹鋼蓋代替泥封窖的100年窖池,窖壁泥pH高于其余2種窖齡,總酸和總酯含量相反。窖底泥的總酸和總酯含量顯著高于窖壁泥,JB100的pH高于JD100,其余2種窖齡的窖底和窖壁泥pH略有差異,類似曾報道的結果[16]。
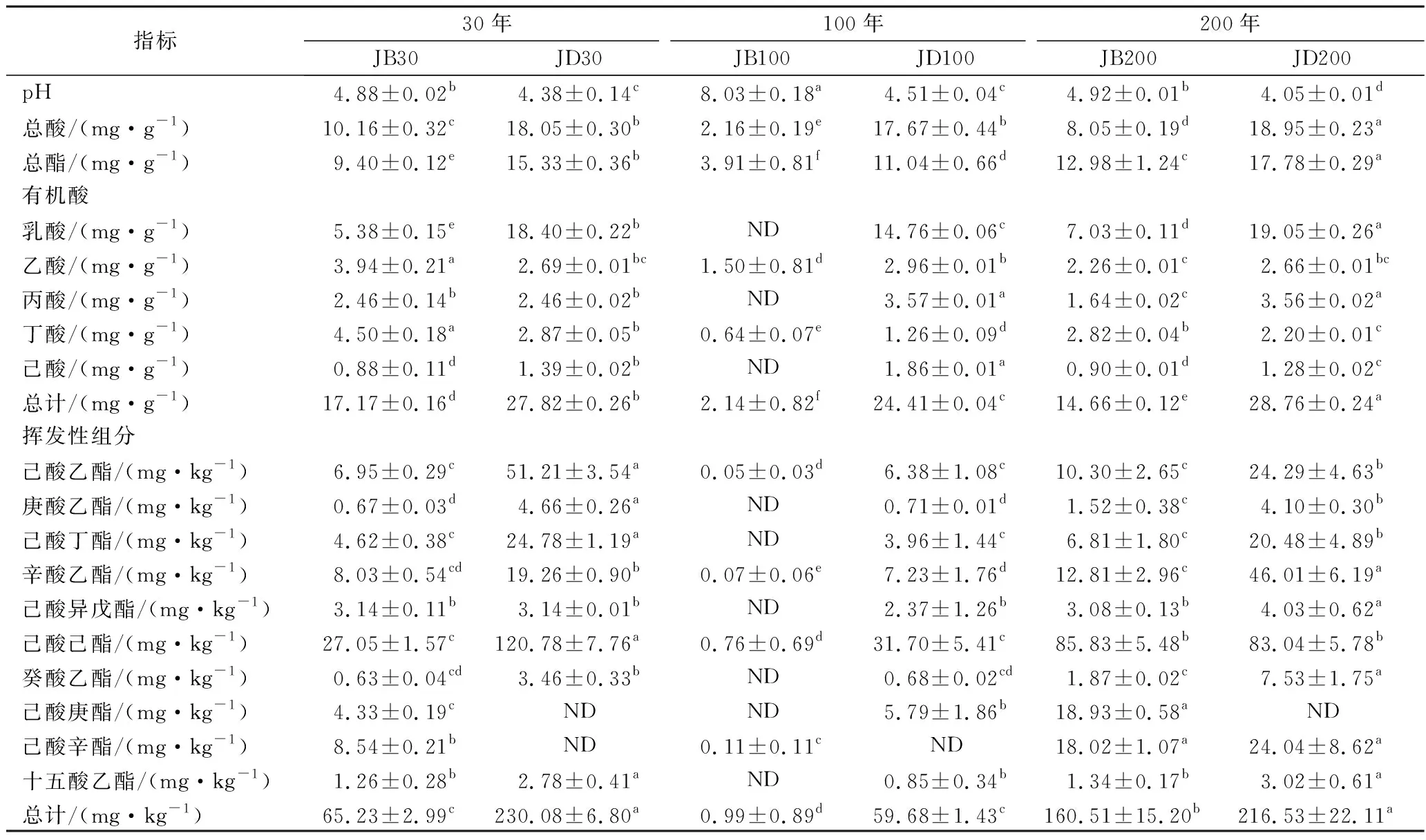
表2 理化及主要代謝組分的差異Table 2 Differences of physiochemical properties and main metabolic components
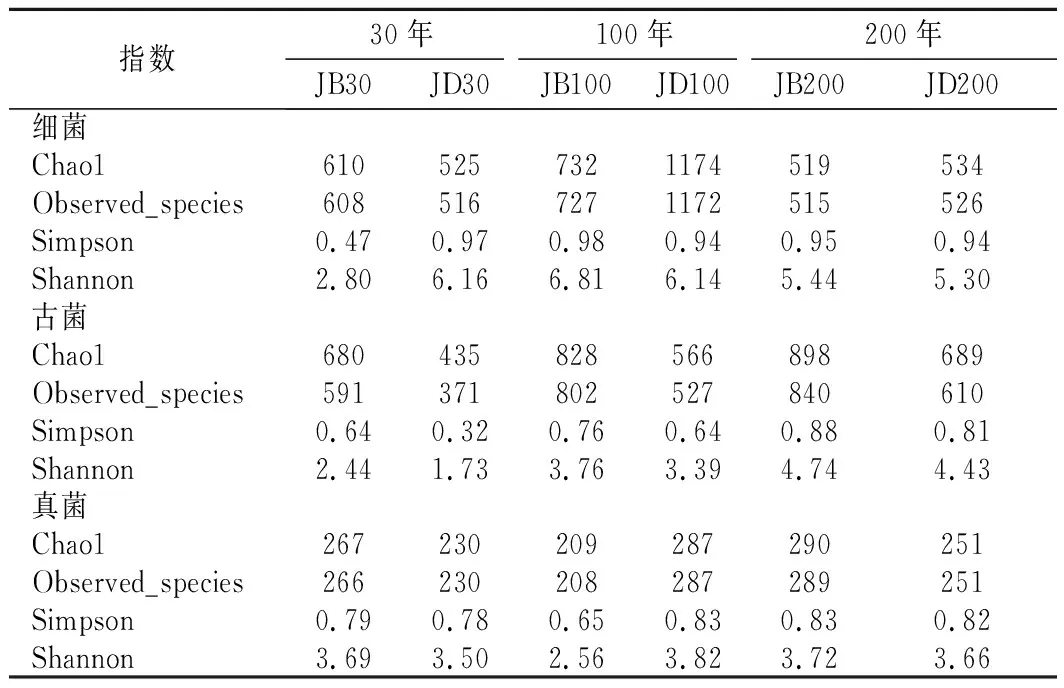
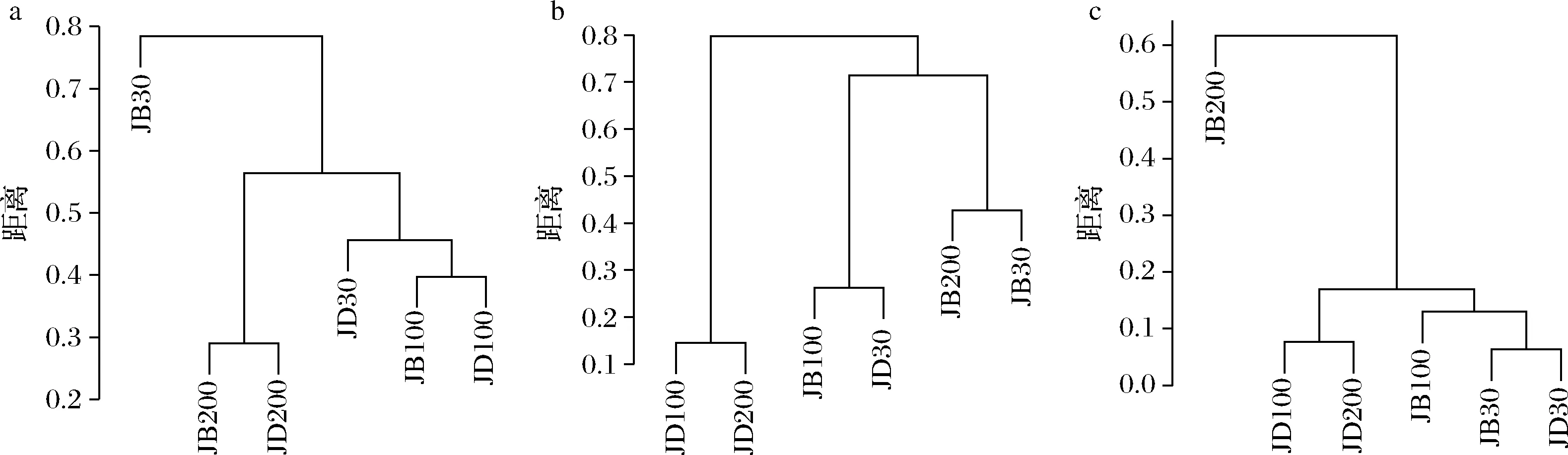
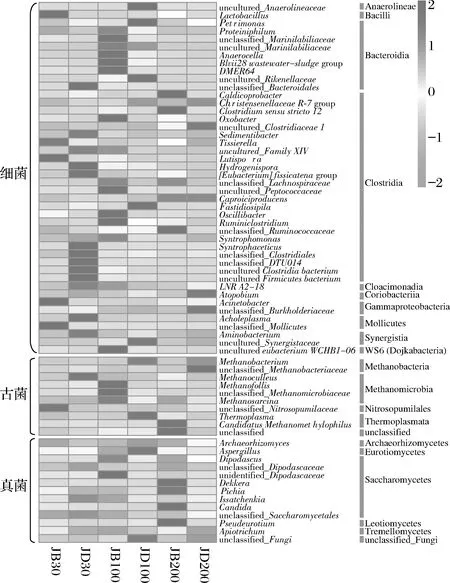
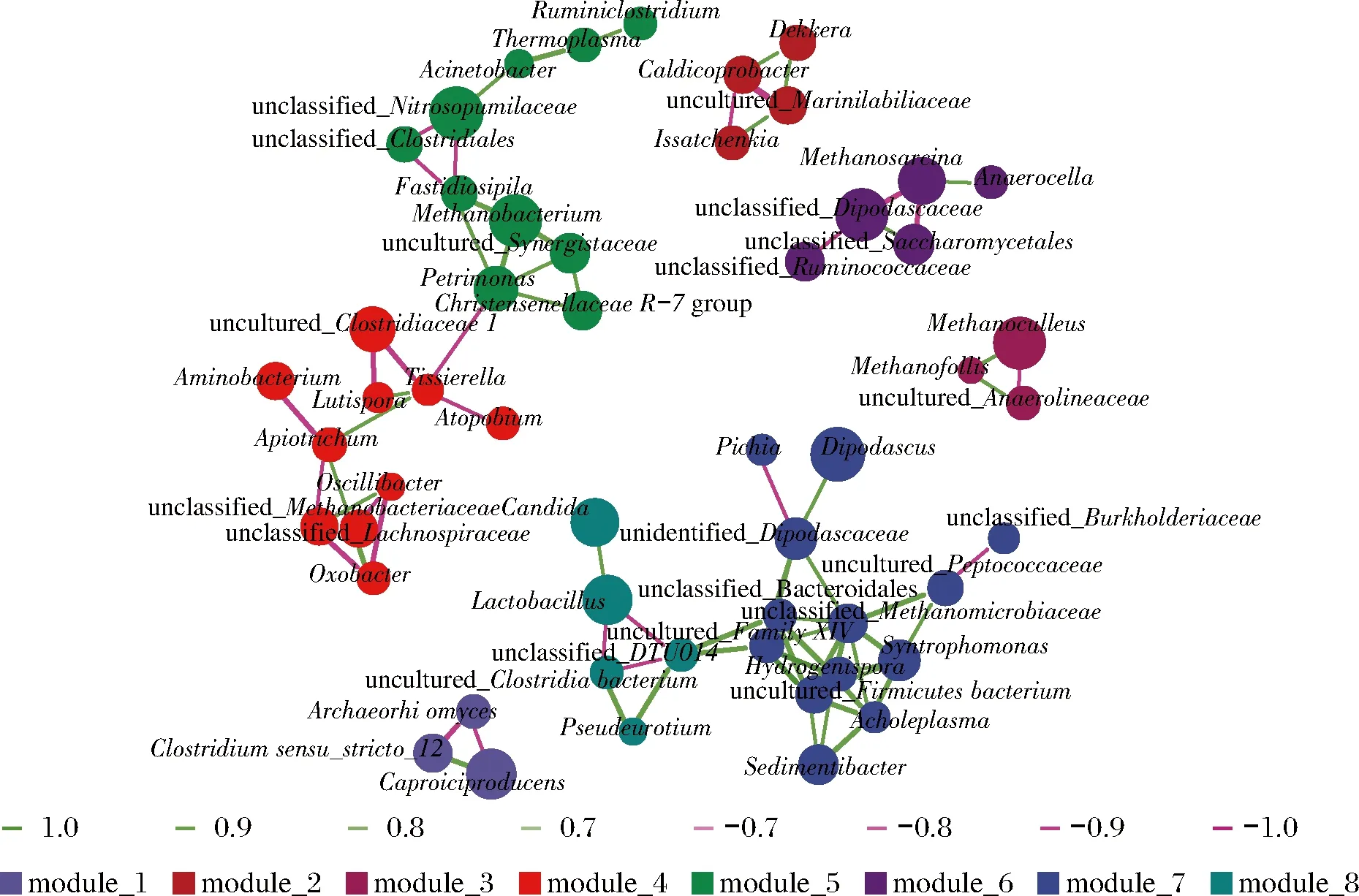
各樣品共檢出了5種有機酸和31種酯類成分,有機酸及10種優勢酯類組分(豐度排名前10),含量如表2所示。窖底泥有機酸及這些酯類組分總含量顯著高于窖壁泥,窖底泥間有機酸含量略有差異,JD100酯類組分最低。JB100有機酸和酯類組分含量顯著低于JB30和JB200,后兩者間也略有差異。JB30中乳酸、乙酸和丁酸是優勢有機酸,JB200乳酸是優勢有機酸,乙酸和丁酸次之。JB100僅檢出己酸乙酯、辛酸乙酯、己酸己酯和己酸辛酯,其中己酸己酯的豐度是76.77%,JD100己酸己酯豐度是53.13%。30年和200年窖池泥中的己酸乙酯、辛酸乙酯、己酸已酯和己酸辛酯是優勢組分,JB30中己酸乙酯和己酸己酯的豐度 如表3所示,100年窖池的細菌α-多樣性指數中的Chao1及Observed species高于30年和200年窖池。可能因封窖方式不同,增大了細菌的物種數,且窖壁和窖底物種數量差異顯著。200年窖池中細菌物種數略低于30年窖池,無論是稀有還是豐富物種,30年和100年窖池細菌菌群易被外源物種擾動,尤其是100年窖池。30年和100年窖池泥中的古菌群的物種數量低于200年的,且隨著窖齡的增長,窖壁和窖底泥差異減小。真菌的物種數量僅略有差異。 表3 不同窖泥微生物群落α-多樣性的差異Table 3 Differences of microbial communities′ α-diversity among pit muds 基于Bray-Curtis距離的微生物群落UPMGA結果如圖1所示。JB200和JD200的細菌群落結構類似,100年窖池的也僅略有差異,JB30和JD30差異顯著,類似DENG等[17]的結果。每個窖池古菌群落均未聚到一簇,與α-多樣性所呈現的規律一致。JB200真菌結構與其余窖泥差異顯著。 a-細菌;b-古菌;c-真菌圖1 不同窖泥微生物群落聚類分析Fig.1 Cluster analysis of microbial communities in pit muds 如圖2所示,群落中的優勢種屬(相對豐度>1%)分別由44種細菌、10種古菌及13種真菌組成,相對豐度總和分別為88.4%~92.8%、98.8%~99.7%和92.1%~97.4%。優勢細菌主要是ChristensenellaceaeR-7 group等12個種屬。熱圖分析表明Clostridiumsensustricto12、uncultured_Clostridiaceae1、Caproiciproducen、unclassified_Ruminococcaceae和ChristensenellaceaeR-7 group等5個種屬的豐度與窖齡呈正相關,類似WANG等[4]報道的結果。Clostridium和Caproiciproducens分別能將乙醇和乳酸轉化為己酸[18-19]等濃香型白酒呈香組分的前體物質。窖底泥中ChristensenellaceaeR-7 、groupAminobacterium、Petrimonas和uncultured_Clostridiaceae1的豐度高于窖壁泥,可能與窖底厭氧程度有關[20-22]。7個隸屬于梭菌綱的屬豐度為36.4%,與窖齡有關,4個隸屬于梭菌綱、擬桿菌綱和互養菌綱的屬豐度為16.6%,與位置有關。30年和200年窖池中Lactobacillus的平均豐度(36.9%和8.8%)顯著高于100年窖池(0.9%)。 圖2 不同窖泥屬水平優勢微生物群落組成熱圖Fig.2 Heat map of dominant microbial communities in pit muds at genus level Methanobacterium、Methanoculleus、Methanosarcina和unclassified_Nitrosopumilaceae是主要優勢古菌,與位置有關。窖底泥甲烷菌及Methanobacterium的豐度較窖壁泥高,而Methanosarcina相反。unclassified_Nitrosopumilaceae是好氧菌[24],在窖壁泥中的豐度高于窖底泥,此外在100年窖池中豐度最低。 Dipodascus、Candida、Apiotrichum、unclassified_Dipodascacea、unidentified_Dipodascaceae和unclassified_Saccharomycetales是主要優勢真菌。隸屬于Dipodascaceae的3種屬在所有窖泥中均被檢出,豐度在24.9%~90.2%,Dipodascus在Dipodascaceae中的比例>52.2%。Candida在JB200中的豐度是53.6%,在其余5個窖泥中則< 5.4%。Candida能代謝乙醇和乳酸轉化為風味組分[24]。 應用FISH檢測3種不同窖齡窖泥中的細菌群落結構結果如表4所示。JB100真細菌的數量高于JB30的,JD100真細菌數量高于JD30和JD200的。梭菌屬的數量在窖底高于窖壁,且隨著窖齡延長而增高,100年窖池梭菌屬的比例略高于其余2種窖齡。產甲烷古菌的總量隨窖齡增長而增高,窖壁氫營養型古菌的比例隨窖齡增加而增高,窖底泥中則是乙酸營養型古菌的比例增高。值得注意的是FISH檢測的結果與高通量測序的結果顯著不同,前者檢出的甲烷桿菌目的比例>36%,而后者的結果<3%,主要是因為FISH是活細胞水平檢出,而后者在OTU水平,樣品既包括活細胞又有死亡細胞,引物和擴增速率誤差等干擾也難避免。2種方法的趨勢相同則從細胞和分子水平反映窖齡及位置的影響規律。 表4 窖泥微生物群落的熒光原位雜交檢測結果 單位:109cell/g 綜合以上分析,窖池的環境,尤其是酸度/pH顯著影響群落結構及代謝,群落及代謝又影響窖泥小生境,這些因素互作是窖泥群落定向進化的關鍵環境脅迫作用[25],封窖方式則可能是影響定向進化另一個關鍵的環境脅迫因素。 基于斯皮爾曼相關性 (|ρ|>0.6,P<0.05)分析豐度> 1.0%屬水平的微生物的結果如圖3所示。這些微生物包含54對共現、26對排斥關系。模塊化分析結果表明關聯網絡主要由模塊內和模塊間互作等2種關聯模式構成。在模塊內關聯的3個模塊中,module_1的Caproiciproducens和Clostridiumsensustricto12呈正相關,這2個屬隨窖齡的變化趨勢相同,它們是窖泥中主要的己酸菌,能夠通過逆向β-氧化合成乙酸、丁酸和己酸,其中乙酸和丁酸是己酸合成中的電子受體[26],兩者的一些產物互為彼此的二次發酵底物。module_6則是Methanosarcina與Anaerocella呈正相關,豐度與位置的關系相同。module_3中Methanoculleus和Methanofollis呈正相關,它們同為甲烷微菌科古菌,生理特性和應激能力相似,在窖池微環境變化的過程中具有相同的變化趨勢。這類網絡結構簡單,節點數較少,窖池微環境的變化如乳酸、含氧量等可能是引起微生物群落發生變化的主要因素。模塊間關聯共有4個模塊,又分為多對一和多對多的2類互作關系。在多對一的互作關系中,如module_7和module_8,Sedimentibacter和Syntrophomonas等為中心的共現群落與Lactobacillus和Candida共現對間呈負相關,而與Dipodascus呈正相關。在多對多的互作關系中,如module_4和module_5,Petrimonas與Methanobacterium等組成的共現群落與由unclassified_Nitrosopumilaceae等組成的共現群落以及由uncultured_Clostridiaceae1等組成的互作群落之間呈負相關。這類互作關系,除自身復雜外,群落豐度還與位置有關,可能是小生境脅迫與群落互作共同致其改變。Lactobacillus的產物乳酸的積累,降低了窖泥pH,可能改變群落結構,Lactobacillus與Candida呈正相關可能是因為二者之間存在協同共生關系[27]。為了維持群落的動態平衡,種間氫轉移,與甲烷菌互養,提高碳利用率及促進碳循環效率[21-22,28]是濃香型白酒窖池發酵重要調控機制。 圖3 窖泥微生物群落關聯網絡分析Fig.3 Network analysis of microbial communities in pit mud 基于高通量測序所得窖泥核心菌群與主要風味組分間的RDA結果見圖4。Lactobacillus與乙酸和丁酸呈正相關,但與乳酸和己酸呈負相關。發酵過程中乳酸隨黃水傳遞至窖底,窖底泥乳酸的含量高于窖壁,而乳酸菌豐度隨窖池的深度增加而降低[7]。Lactobacillus與己酸乙酯、己酸己酯和辛酸乙酯呈負相關。乳酸是Caproiciproducens和Clostridiumsensustricto12的底物[19],與之正相關,乙酸、丁酸作為合成己酸的電子受體被轉化,所以呈負相關。己酸與醇縮合為己酸酯,雖然Caproiciproducens和Clostridiumsensustricto12與己酸僅是弱相關性,但與己酸酯正相關。Petrimonas等與乳酸、己酸和己酸乙酯呈正相關。Methanobacterium與己酸呈強烈的正相關,unclassified_Nitrosopumilaceae與己酸呈負相關,Methanoculleus與己酸酯呈正相關。Methanobacterium和Methanoculleus是氫營養型古菌[29],能夠降低生物合成己酸過程中的生物氫氣[18],促進己酸和己酸酯的合成。自養型古菌Nitrosopumilaceae[23]和Clostridium[30]都以CO2作為底物,二者底物的競爭降低了己酸的合成速率。Dipodascus和Candida僅與丁酸和己酸己酯等相關。 a-細菌;b-古菌;c-真菌圖4 核心微生物與風味之間的冗余分析Fig.4 Redundancy analysis between core microorganisms and flavor 應用高通量測序及FISH研究窖齡及位置對微生物群落結構的影響,百年老窖泥的細菌群落中Caproiciproducens和Clostridiumsensustricto12的豐度高,窖壁和窖底的古菌群落結構差異顯著,前者Nitrosopumilaceae顯著高于后者的,產甲烷古菌的豐度則在后者中更高。除了窖齡和位置以外,封窖材料可能也對微生物群落結構產生影響,不銹鋼蓋窖池中Lactobacillus和Nitrosopumilaceae的豐度顯著低于泥窖,其具體的影響機制還需要進一步研究。群落結構的差異導致理化性質及酯代謝組分組成及含量差異顯著。RDA的結果表明Caproiciproducens和Clostridiumsensustricto12與乳酸和己酸酯呈正相關,產甲烷古菌與己酸和己酸酯呈正相關。FISH檢出的菌群輪廓差異與高通量測序檢測的結果是相似的。2.2 窖齡及位置對微生物群落α-多樣性的影響
2.3 窖齡及位置對微生物群落組成的影響

2.4 優勢菌的相關性分析
2.5 核心微生物與窖泥風味之間的RDA

3 結論
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
