漢防己甲素衍生物HL-49對Bloom DNA解旋酶生物學特性的影響
2022-02-24 06:47:18劉金河張望明晏文濤楊凱琳徐澤權劉杰麟
中國藥理學通報 2022年2期
關鍵詞:實驗
劉金河,張望明,晏文濤,向 天,羅 維,楊凱琳,徐澤權,劉杰麟
(1.貴州醫科大學細胞工程生物醫藥技術國家地方聯合工程實驗室,組織工程與干細胞實驗中心,貴州省再生醫學重點實驗室,貴州 貴陽 550004;2.貴州醫科大學基礎醫學院免疫教研室,貴州 貴陽 550025;3.貴州中醫藥大學第一附屬醫院檢驗科,貴州 貴陽 550001;4.貴州醫科大學檢驗學院,貴州 貴陽 550004)
Bloom DNA解旋酶(bloom DNA helicase,BLM)是RecQ解旋酶家族中的一員,能利用三磷酸腺苷(ATP)和其他三磷酸核苷水解釋放的能量解開雙鏈DNA或G四鏈體,是核酸復制、重組、修復、轉錄、端粒穩定等代謝過程的重要組成部分[1-2]。
研究發現,BLM基因突變能引起皮膚鱗狀細胞癌的發生[3]和前列腺癌轉移有關[4]。Wang等[5]發現喹唑啉酮為母核合成的衍生物中,9 h能顯著抑制BLM 解旋酶與DNA的結合活力,也促進端粒DNA的瓦解,誘導細胞凋亡。因此,研究小分子對BLM DNA解旋酶功能的影響及以BLM DNA解旋酶為靶向的腫瘤治療具有重要意義。
漢防己甲素(tetrandrine,TET)是一種雙芐基異喹啉類生物堿,具有廣泛的抗腫瘤活性,例如肺癌、結腸癌、乳腺癌等[6]。臨床研究發現,該藥物毒性小、副作用低,后期成藥性較好。TET能通過上調IGFBP-5蛋白的表達增強p53蛋白表達進而抑制乳腺癌MCF-7細胞增殖[7],Li等[8]研究發現,TET通過調節CCND1/CDK4化合物及其下游蛋白磷酸化Rb(p-Rb)抑制結腸癌 HT-29 細胞的生長。Bhagya等[9]證實了TET激活活性氧(reactive oxygen species,ROS)進而通過caspase 途徑誘導乳腺癌和胰腺癌細胞凋亡,Sun等[10]發現TET檸檬酸鹽上調 ROS 水平,下調Bcl-2以及上調cleaved caspase-3、Fas、p-p38和 p-JNK 的表達水平來介導抗腫瘤活性。因此TET抗癌潛在分子機制可能與誘導癌細胞凋亡、抑制細胞增殖、遷移、侵襲并抑制腫瘤細胞生長有關[6]。
目前研究漢防己甲素衍生物對腫瘤抑制作用的文章不多,以BLM DNA解旋酶作為靶點,分析其與BLM DNA解旋酶相互作用的分子機制的文章鮮見報道。本文擬使用熒光偏振檢測、紫外光譜掃描、孔雀綠-磷鉬酸銨比色等技術和方法,通過研究漢防己甲素衍生物HL-49對BLM DNA解旋酶結構和生物學活性的影響,揭示漢防己甲素衍生物抑制腫瘤細胞生長的可能機制。
1 材料與方法
1.1 藥物與試劑漢防己甲素衍生物HL-49:(貴州省中國科學院天然產物化學重點實驗室潘衛東教授課題組)提供;ssDNA底物委托北京鼎國昌盛生物技術有限責任公司合成。序列如下:A1(45bp):5′-AATCCGTCGAGCAGAGTTAGGTTAGGTTAGGTT-AGTTTTTTTTTT-3′;A2(21bp):3′-FAM-TTAGGCA-GCTCGTCTCAATCC-5′。(注:FAM為羧基熒光素)。
DNA底物雜交緩沖液(20 mmol·L-1Tris-HCl,100 mmol·L-1NaCl,pH 7.9);熒光偏振實驗反應緩沖液(20 mmol·L-1Tris-HCl,25 mmol·L-1NaCl,3 mmol·L-1MgCl2和0.1 mmol·L-1DTT,pH 7.9);紫外光譜實驗緩沖液buffer C (20 mmol·L-1Tris,500 mmol·L-1NaCl,500 mmol·L-1咪唑,10%甘油,pH 7.9);ATPase活性檢測實驗所需三乙醇胺、孔雀綠、鉬酸銨均用雙蒸水進行配置。其他化學試劑均為國產分析純。
1.2 儀器NGCTM蛋白質分離純化系統(美國BIO-RAD公司);高壓細胞破碎儀(英國Constant Systems公司);Synergy 4多功能酶標儀(美國BIO-TEK公司);UV-2100紫外可見分光光度計(日本SHIMADZU公司);Milli-Q超純水系統(美國Millipore Corp公司)。
1.3 方法
1.3.1BLM DNA解旋酶的制備、純化 將表達 BLM DNA 解旋酶的重組大腸桿菌置于含有 50 mg·L-1氨芐青霉素+30 mg·L-1氯霉素的LB培養基中,在37 ℃、160 r·min-1的條件下擴大培養至OD值達到0.5。然后轉到18 ℃、180 r·min-1的條件,加入終濃度0.4 mmol·L-1IPTG 誘導Bloom DNA解旋酶表達20 h,隨后在4 ℃、4 000 r·min-1的條件下離心15 min收集菌體。使用高壓細胞破碎儀在21 kpsi壓力下進行一次性破碎,收集流出液,4 ℃、13 000 r·min-1的條件下離心45 min收集上清液,通過鎳親和層析和分子篩層析進行分離純化,獲得能用于進行酶生物學特性研究的重組BLM DNA解旋酶[11]。
1.3.2HL-49對 BLM DNA解旋酶紫外光譜的影響 Buffer C 緩沖液(pH 7.9)中加入終濃度為2.7 μmol·L-1的BLM DNA解旋酶與不同終濃度(0~20 μmol·L-1)的HL-49,保持整個反應體系總體積為200 μL。將混合液置于紫外分光光度計測量室內,設定掃描速度為中等速度,掃描波長間隔為1 nm,在230~300 nm波段下進行掃描,以每5 min掃描1次的頻率掃描至平衡,取3次平衡值的圖譜進行擬合。根據光譜峰值和峰形的變化即可判定蛋白質的構象是否發生變化[11]。再用相同的方法掃描其余不同終濃度(0~20 μmol·L-1)的 HL-49 在buferC緩沖液中的紫外吸收光譜。
1.3.3HL-49對BLM DNA解旋酶的DNA結合活性的影響 本實驗采用熒光偏振法測定 HL-49對 DNA與BLM DNA解旋酶結合活性的影響。采用2種方式研究 HL-49對BLM DNA解旋酶 DNA結合活性的影響[12]。第1種,25 ℃下,用不同濃度的HL-49(0~25.33 μmol·L-1)滴定BLM DNA解旋酶與2 nmol·L-1的DNA(A2或A1A2)相互孵育形成的復合物;第2種是25 ℃下,不同濃度的HL-49先與BLM DNA解旋酶反應5 min,再加入到含2 nmol·L-1DNA(ssDNA/dsDNA)的反應緩沖液中進行檢測。實驗重復3次,數據為3~5個穩定偏振值平均之后的結果。通過調節雙蒸水的體積,使實驗的總體積保持在150 μL。
1.3.4HL-49 對 BLM DNA 解旋酶 DNA 解鏈活性的影響 本實驗采用熒光偏振法檢測HL-49對BLM DNA解旋酶DNA解鏈活性的影響。取終濃度為75 nmol·L-1的BLM DNA解旋酶與不同終濃度(0~20 μmol·L-1)的HL-49混勻,在室溫下孵育5 min,加入到含2 nmol·L-1的dsDNA的反應緩沖液中,測定熒光偏振值至穩定,再加入終濃度為0.2 mmol·L-1的ATP,測定熒光偏振值至穩定。取各個反應穩定時5~7個的熒光偏振值的平均值,實驗重復3次。通過調節雙蒸水的體積,使本次實驗的總體積保持在150 μL。DNA解鏈的速率常數可根據方程(1)獲得:
At=A1exp(-kobst)
(1)
式中At是解鏈時間為t時的熒光偏振值,A1是解旋酶與dsDNA完全結合時的熒光偏振值。
1.3.5HL-49 對 BLM DNA 解旋酶 ATP 酶活性的影響 本實驗采取孔雀綠-磷鉬酸銨比色法來測定 HL-49 對 BLM DNA解旋酶 ATPase 活性的影響。首先,反應緩沖液(50 mmol·L-1三乙醇胺、50 mmol·L-1KCl、20 mmol·L-1MgCl2,pH 7.5)中加入終濃度為100 nmol·L-1的ssDNA,終濃度為75 nmol·L-1的 BLM DNA 解旋酶和不同終濃度(0~100 μmol·L-1)的HL-49于室溫下孵育10 min。隨后加入終濃度為2 mmol·L-1的ATP,在室溫下分別孵育(0、5、10、15、20、25、30)min,通過調節雙蒸水的體積,使本次反應的總體積保持在75 μL。然后取 50 μL上述反應液迅速加入850 μL染液(36 mL 0.045%孔雀綠溶液,12 mL 4.2%鉬酸銨溶液,1 mL 1%的Triton X-100)中以終 ATP的水解反應,在3 min時,往染液中加入100 μL 34% 的檸檬酸溶液來終止顯色反應。終止顯色反應后,各不同濃度藥物組吸取100 μL 加至96孔板中,于660 nm波長處讀值,實驗重復3次。使用國際單位定義表示酶量的多少,即每分鐘催化1 μmol底物水解所需的酶量為1個國際單位(unit)。酶活力Activity(kU·L-1·min-1)可通過以下公式(2)計算
(2)
式中,A是根據標準曲線得出的磷酸鹽濃度(μmol·L-1),B是反應時間(min)。
2 結果
2.1 HL-49對BLM DNA解旋酶的構象無影響HL-49對BLM DNA解旋酶紫外光譜影響的實驗結果見Fig 2。對于蛋白質來說,肽鍵的強吸收峰一般出現在210~240 nm處,而芳香族氨基酸殘基中共軛雙鍵的吸收峰出現在280 nm附近。由 Fig 2A可以看出,HL-49在238 nm和280 nm 處具有兩個峰。隨著HL-49濃度的增加,BLM DNA 解旋酶在236 nm和280 nm處的峰位置和峰型未發生明顯變化(Fig 2B),再結合HL-49對BLM DNA解旋酶在236 nm和280 nm處的紫外吸收值約等于HL-49和BLM之和(Fig 2C)可知兩者之間并無相互作用,由此推斷HL-49不能改變BLM DNA解旋酶的三維構象。

Fig 1 Determination of BLM DNA helicase purity after molecular sieve chromatography
2.2 HL-49對BLM DNA解旋酶的DNA結合活性無影響HL-49對BLM解旋酶的DNA結合活性的影響見Fig 3。由圖Fig 3A和Fig 3B可知,無論是dsDNA(Fig 3A)還是ssDNA(Fig 3B)作為底物,HL-49對BLM DNA解旋酶因結合而增加的熒光偏振值影響非常小,也不具濃度依賴性,熒光偏振值的增量(ΔA1,ΔA2)無明顯變化(P>0.05),說明HL-49并不影響BLM DNA解旋酶與DNA的結合;然而,HL-49 單獨滴定dsDNA或者ssDNA 時,熒光偏振值的增量(ΔA3)隨著HL-49濃度的增加而明顯增大(P<0.01),說明HL-49可與DNA(dsDNA or ssDNA)相結合(Fig 3C),且結合能力與HL-49的濃度呈正相關。
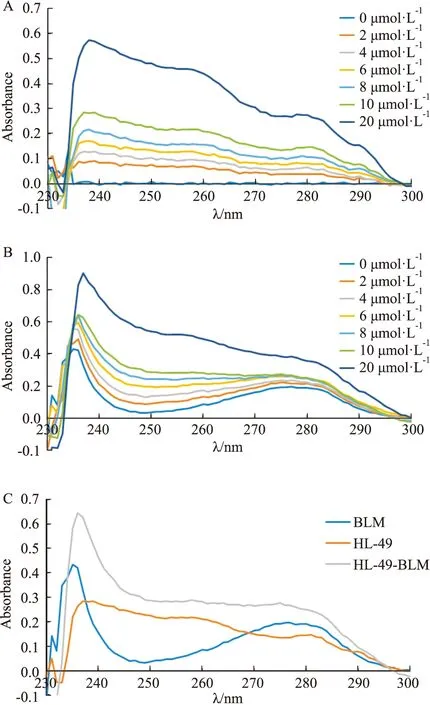
Fig 2 The Ultraviolet (UV)absorption spectra of HL-49 interacted with BLM DNA helicaseA:The changes of UV absorption spectra by different concentrations of HL-49;B:The changes of UV absorption spectra by different concentrations of HL-49 interacted with BLM helicase treatment;C:The UV absorption spectra of BLM (2.7 μmol·L-1),BLM (2.7 μmol·L-1)and HL-49(10 μmol·L-1),HL-49(10 μmol·L-1 );D:The titration curve of BLM helicase for the addition of HL-49 at the 236 nm.
2.3 HL-49能抑制 BLM DNA 解旋酶的DNA解鏈活性利用熒光偏振方法研究HL-49對BLM DNA解旋酶的DNA解鏈活性的影響。由Fig 4A可知,隨著反應體系中HL-49終濃度的逐漸增加(0~25.33 μmol·L-1),因解鏈而降低的熒光偏振值(A1-A2)逐漸減小,表明BLM DNA 解旋酶的解鏈活性與HL-49的濃度呈負相關。根據公式(1)進一步分析HL-49存在下BLM解旋酶的Kobs值的變化(Fig 4B),可知隨著HL-49濃度的增加,BLM解旋酶的Kobs值逐漸減小,表明HL-49能夠明顯抑制BLM解旋酶的DNA解鏈活性。

Fig 3 Effects of HL-49 on DNA-binding activity of BLM DNA Fluorescence anisotropy values were determined as a function of the helicase concentration both A and B.C:The effects of HL-49 on dsDNA and ssDNA were assayed by fluorescence polarized technology.2 nmol·L-1 DNA substrate was pre-incubated in the DNA binding buffer for 5 min at 25 ℃.ΔA1 and ΔA2 represent the increased value of the total fluorescence polarization value after the addition of HL-49 minus the fluorescence polarization value of DNA.ΔA3 represents the total fluorescence polarization value of HL-49 after addition minus the value of DNA.
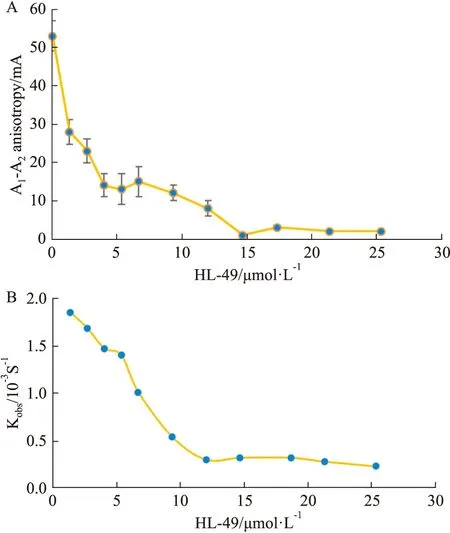
Fig 4 Effects of HL-49 on DNA unwinding activity of BLM DNA A:Effects of different concentrations of HL-49 on the unwinding activity of BLM.B:The change of Kobs values of the BLM helicase to dsDNA in the presence of different concentrations of HL-49.The Kobs values were calculated from equation (1).A1 is the fluorescence polarization value of HL-49 interacting with BLM helicase and DNA at different concentrations.A2 is the fluorescence anisotropy of the reaction system after the addition of 2 nmol·L-1 ATP.
2.4 HL-49 能抑制BLM DNA 解旋酶的ATPase活性由Fig 5A所示可知,隨著HL-49濃度的逐漸增加,BLM DNA解旋酶的ATPase活性逐漸降低,揭示出BLM DNA解旋酶的酶活力與HL-49的濃度呈負相關(P<0.01)。由Fig 5B可知,在相同HL-49濃度下,隨著時間的延長(0~30 min),BLM DNA解旋酶的酶活力逐漸下降。通過米氏方程雙倒數作圖法進一步分析實驗數據,計算出酶反應的Vmax、Km等常數,結果見Tab 1,可知隨著HL-49濃度的增加,Vmax保持不變,而Km逐漸增大,且Kcat減小,說明HL-49對BLM DNA解旋酶的作用為可逆性競爭性抑制作用。
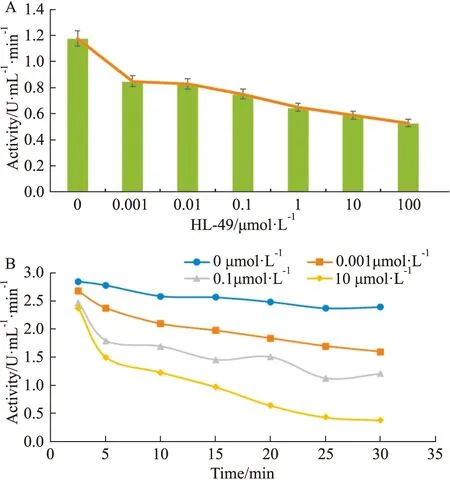
Fig 5 Effects of HL-49 on ATPase activity A:Curves of activity that based on the equation (2)of recombinant BLM helicase in the presence of different concentrations of HL-49.In each reaction,DNA was present at a concentration of 100 nmol·L-1 and ATP was 2 mmol·L-1.B:Time course of ATP hydrolysis by 2.7 μmol·L-1 recombinant BLM helicase in the presence of different concentrations of HL-49 at 30 min.Experiments were performed at 25 ℃ in ATPase activity assay buffer containing 50 mmol /L Tris-HCl (pH 7.9),2.5 mmol /L MgCl2.
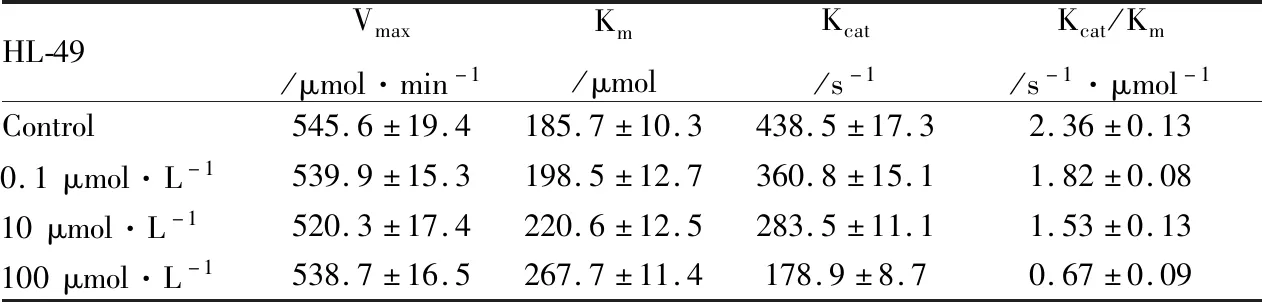
Tab 1 The ATPase activity constants of HL-49 interacted with BLM DNA helicase
3 討論
由紫外光譜掃描和DNA結合實驗結果可知,HL-49不會改變BLM DNA解旋酶的構象,也不會與其相互結合,且能與DNA(dsDNA/ssDNA)相互結合并具濃度依賴性。DNA解鏈活性的結果顯示:隨著HL-49濃度增大,BLM DNA解旋酶的解鏈活性逐漸受到抑制,抑制作用明顯(P<0.01)。ATPase活性隨著時間的延長而明顯降低,且隨著HL-49濃度的增加,ATPase活性逐漸降低。由此可知,BLM DNA解旋酶的ATPase活性具時間依賴性和濃度依賴性,進一步分析酶的相關動力學參數Km逐漸增大、Vmax不變且Kcat值減小,可知HL-49抑制BLM DNA解旋酶生物學活性的類型為DNA的可逆性競爭。由以上實驗結果可知,HL-49抑制BLM DNA解旋酶的生物學活性的可能機制為HL-49可逆性結合于DNA(ssDNA/dsDNA)鏈上,阻礙BLM DNA解旋酶與DNA的結合,進而影響BLM DNA解旋酶的DNA解鏈活性和ATPase活性。
BLM DNA解旋酶作為人體細胞當中存在的一種具解開雙鏈DNA活性的蛋白,在DNA的復制、重組、轉錄和端粒維持的功能中發揮重要作用。BLM基因的表達異常造成BS綜合征,其臨床表現為高水平的姐妹染色單體互換、身材矮小、免疫系統缺陷、不孕不育、癌癥發生率高[12]。
BLM DNA解旋酶在大多數腫瘤細胞中普遍高表達,但表達水平因腫瘤細胞系而異[13],故以BLM DNA解旋酶為抗癌研究靶標,利用小分子化合物抑制癌癥細胞中BLM DNA解旋酶的表達及活性,抑制細胞的增殖,最終抑制癌癥的發生發展,在理論上是可行的。因此,尋找一種療效好,副作用低,成本低廉且靶向抑制BLM DNA解旋酶活性的小分子化合物具有顯著的臨床意義。而漢防己甲素目前作為臨床常用藥物,普遍用來治療各種炎癥反應,毒副作用很小,且提煉成本較低,具有先天優勢,在此基礎上改造而來的一系列漢防己甲素衍生物,比如HJNO,HL-49[14-15],都表現出了很好的抗腫瘤活性,而本文對HL-49抑制腫瘤的分子機制的研究印證了其抗腫瘤活性。該研究的結果能為后續的動物實驗、臨床實驗提供一定的參考。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
