乳酸菌對干酪風味形成作用的研究進展
2022-03-09 01:48:58司闊林閆清泉李玲玉任敏趙中華宗學醒
食品工業 2022年2期
關鍵詞:途徑
司闊林,閆清泉,李玲玉,任敏,趙中華,宗學醒
內蒙古蒙牛乳業(集團)股份有限公司(呼和浩特 011500)
干酪(cheese)是一種營養豐富的發酵乳制品,世界上有上千種干酪,每種干酪都具有本身所獨有的風味和質構。干酪風味的形成是一個動態且復雜的生物化學過程(圖1)[1],干酪風味物質主要為微生物群對乳脂、蛋白質和碳水化合物的綜合代謝并伴隨著干酪基質中酶的進一步生物化學轉化而產生的揮發性風味物質和芳香性風味物質等混合物,其中乳酸菌(lactic acid bacteria,LAB)在成熟干酪微生物群中占主導地位。因此,通過高效利用乳酸菌,能夠促進干酪成熟,改善或者開發新風味。本文將重點綜述干酪在成熟過程中如何通過一系列生化反應獲得獨特風味及乳酸菌在其中作用兩個方面。

圖1 干酪風味形成的動態過程
1 干酪風味物質形成的關鍵代謝途徑
風味是任何發酵乳制品最重要的屬性之一。消費者愿意嘗試不同口味的乳制品,因此,許多發酵乳制品公司都將發酵劑作為使產品風味多樣化的工具。如圖2所示,干酪風味形成是一個復雜的過程,需要微生物、生物化學和技術等方面的組合,其中乳酸菌主要的代謝途徑為糖(乳糖)酵解、檸檬酸代謝、蛋白質水解和隨后的氨基酸分解代謝,游離脂肪酸的釋放(脂解)及其代謝也參與其中,但分解程度較輕[2]。例如在切達干酪的特征香氣中,一半強烈風味來自于乳糖發酵和檸檬酸鹽代謝(少數來于脂肪降解),另外一半源于亮氨酸和蛋氨酸的分解,其中甲硫氨酸和甲硫醇就是蛋氨酸產生的主要芳香化合物,分別有助于切達干酪的香氣和令人愉悅的大蒜風味[3]。表1列出了4種常見干酪的關鍵風味化合物[4]。
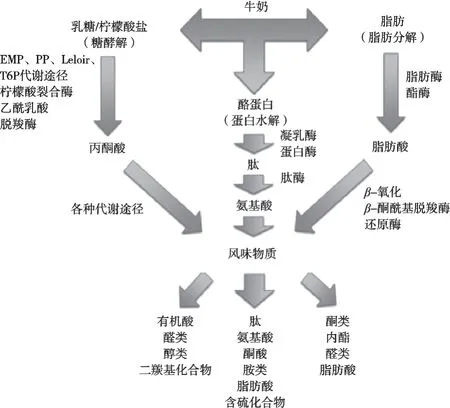
圖2 發酵乳制品風味形成的主要代謝途徑
1.1 蛋白質降解
酪蛋白降解無疑是硬質和半硬質干酪形成風味物質最重要的生化反應。蛋白質降解合成風味物質途徑如圖3所示[5]。在干酪成熟過程中,蛋白質降解產生風味物質的第一步是酪蛋白被乳酸菌細胞壁蛋白酶水解成小肽和氨基酸,其中乳酸乳球菌PrtP是目前最具特色的乳酸菌蛋白酶,且具有相當廣泛的特異性,能夠將酪蛋白分解為100多種不同的寡肽[6],寡肽被寡肽轉運系統(OPP系統)和二/三肽轉運蛋白通過細胞膜攝入細胞內[圖3(A~D)],肽在細胞內被各種肽酶降解為氨基酸。值得一提的是,盡管肽和氨基酸具有特殊的風味特征,如甜味、苦味和麥芽味等,但通常認為它們僅能增加干酪的基本口味,例如刺激或過度表達幾種蛋白水解酶或向干酪中添加氨基酸僅觀察到輕微的風味差異[7]。盡管如此,蛋白質在降解過度及水解不利的情況下會產生過多的苦味肽,超過感官閾值后會嚴重影響干酪的風味[8]。總而言之,盡管蛋白質水解和肽水解對干酪風味形成很重要,但是卻不是蛋白質轉化風味物質過程的關鍵控制點。
如表1所示,3-甲基丁醛、異戊酸和苯甲醛等都是干酪關鍵風味化合物,而這些風味物質都是氨基酸在細胞內被氨基酸轉氨酶[圖3(1)]轉化為α-酮酸再進一步反應而成,目前在乳酸菌中還未發現其他類型的脫氨酶,氨基酸轉氨酶途徑是氨基酸分解代謝的第一步,同時也是干酪風味形成至關重要的一步。有研究在敲除芳香族氨基酸轉氨酶基因后發現半硬質干酪中缺乏苯丙氨酸衍生的風味化合物,再一次證明轉氨作用對形成苯丙氨酸衍生的風味物質是必不可少的[9]。氨基酸另外一個重要因素,干酪風味形成過程中最感興趣的代謝途徑由裂解酶[圖3(10)]啟動,如在大多數乳酸菌中存在的胱硫醚b-裂解酶和半胱氨酸-硫辛酸g-裂解酶,能將蛋氨酸轉化為甲硫醇,另一類裂解酶蘇氨酸醛縮酶[圖3(9)]能將蘇氨酸直接轉化為乙醛[10]。氨基酸的第三種轉化途徑是脫氨基/脫羧成生物胺[11],關于生物胺的健康風險及干酪不同成熟期生物胺量及與細菌菌落結構之間的關系已經進行了研究[12-13],氨基酸的直接脫羧解釋了大多數胺的存在,但還不能解釋仲胺和叔胺的形成。

表1 四種常見干酪的關鍵及其他重要風味物質
干酪風味形成的關鍵中間體α-酮酸,可以進一步被氫化為羥酸[圖3(2)]、被脫羧為醛[圖3(3)]和被脫氫酶復合物脫氫/氧化脫羧為乙酰輔酶A衍生物[圖3(8)],乙酰輔酶A衍生物還可以進一步轉化為風味化合物異戊酸。這些反應大多是酶促的,但是也發現一些化學轉化途徑,例如由苯丙酮酸生成苯甲醛的轉化過程[14]。α-酮酸被氫化而成羧酸,在一些發酵乳制品中已經發現來自支鏈氨基酸、阿拉伯氨基酸和蛋氨酸的羥酸,它們既不是主要的風味化合物,也不是風味化合物的前體,反而會導致α-酮酸濃度降低,從而對風味化合物含量(如醛)產生負面影響。醛通常可以脫氫[圖3(4)]或氫化[圖3(5)]為相應的醇或羧酸,而這些產物也是酯酶和酰基轉移酶的底物[圖3(7)],從而生成(硫代)酯。酯類物質(如丁酸乙酯)有助于切達干酪和高達干酪的風味,但含量過高可能會導致切達干酪水果味缺陷。在卡門培爾干酪中,苯丙氨酸降解產生的苯乙醛、2-苯乙醇和乙酸苯乙酯均帶有玫瑰般花香的成分,從而使干酪產生令人愉悅的花香[4]。除氨基酸代謝外,糖和脂肪代謝也為酯的生成提供了底物。
支鏈醛被醛脫氫酶氧化后會生成相應的支鏈有機酸[圖3(5)],而支鏈有機酸通常被認為是形成(較長)支鏈脂肪酸的底物,最有可能存在于乳酸菌中的一種酮酸脫氫酶復合物[圖3(8)],也能夠將α-酮酸直接轉化為相應的羧酸,如異戊酸。此外,這些羧酸還是其他芳香化合物如酯、硫代酯、甲酚和3-甲基吲哚的前體。大多數乳酸菌中都鑒定出了醇脫氫酶[圖3(4)][15],醛的風味強度高于其相應醇的風味強度(表1),因此,當干酪需要最大風味強度時,這種向醇的轉化可能不利。
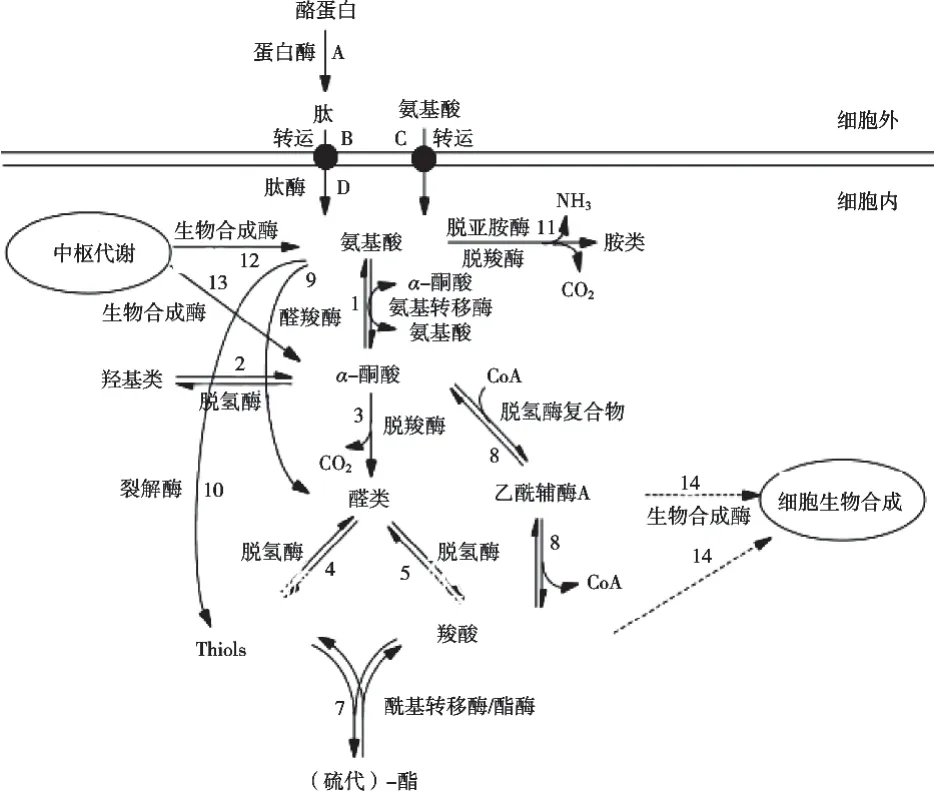
圖3 與干酪風味物質形成相關的蛋白質轉化途徑
1.2 脂肪降解
脂肪降解對干酪的風味和質構也有重要影響。干酪中的脂肪可以提供脂肪-水-蛋白質的接觸面,使香氣形成的反應發生,同時作為溶劑溶解脂溶性香氣化合物,在干酪食用過程時再釋放出來[16]。
脂肪的分解代謝途徑如圖4所示。脂肪分解是由脂肪酶(原生乳脂酶、外源添加的哺乳動物胃前脂酶或微生物脂酶)引起的[17]。干酪脂肪中98%是甘油三酯,甘油三酯被分解后釋放出甘油和游離脂肪酸。長鏈脂肪酸(碳鏈中碳原子數大于12)因為感官閾值較高,在干酪風味中起很小的作用,而短鏈、中鏈脂肪酸及偶數脂肪酸(碳鏈中碳原子數4~12)的感官閾值較低[16],并且每一種脂肪酸都賦予干酪一種特殊的風味,例如丁酸有助于腐敗味和干酪味的形成,己酸有一種辛辣味,是藍紋干酪的風味特征[18]。但是其中的多不飽和脂肪酸容易被氧化產生多種醛類化合物,這些醛類物質風味比較刺激,濃度過高易造成酸敗等不愉悅的風味缺陷,因此要合理控制脂肪分解的程度。而且游離脂肪酸必須與其他風味化合物達到平衡狀態才能產生香氣。相比于游離脂肪酸,大多數衍生自游離脂肪酸(碳鏈中碳原子數為10)的風味活性酯的風味感知閾值要低得多,只是賦予干酪一些水果味。游離脂肪酸同時也是產生一些風味和芳香化合物的分解反應的前體,例如被氧化和脫羧生成甲基酮和仲醇、羥基脂肪酸的酯化生成內酯、與醇基反應形成酯,如丁酸乙酯、己酸乙酯、乙酸乙酯、辛酸乙酯、癸酸乙酯和己酸甲酯等。
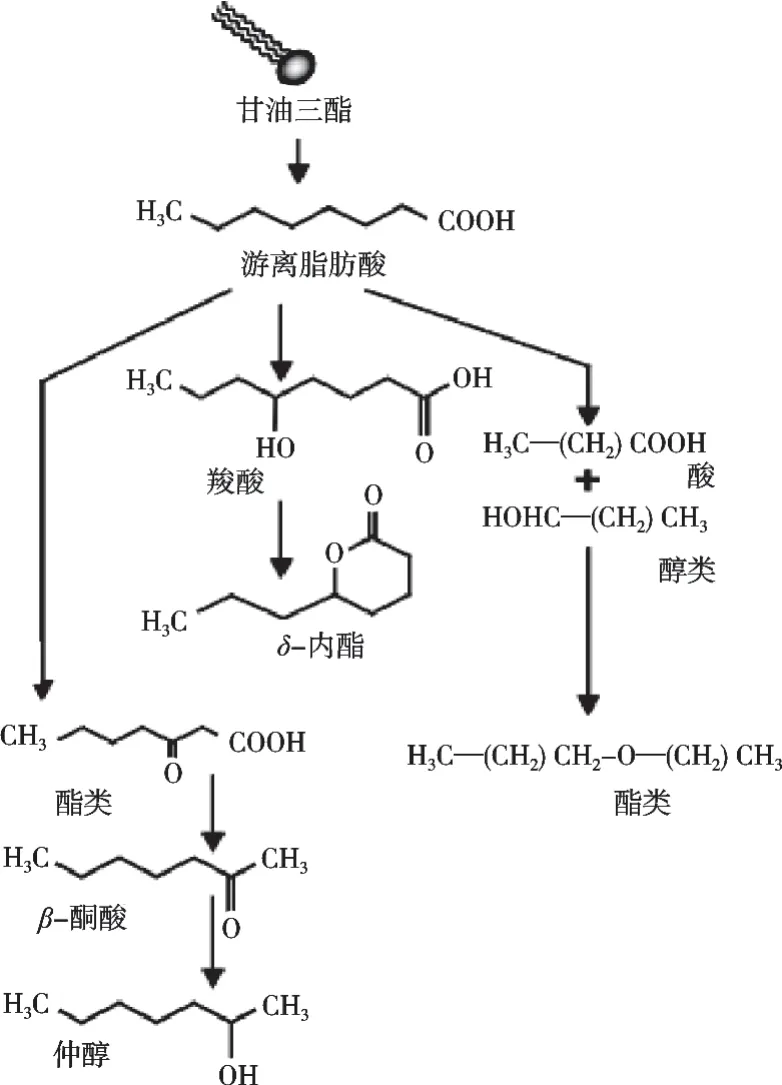
圖4 與干酪風味物質形成相關的脂肪轉化途徑
不同類型的干酪脂肪酸種類、含量不同,所產生的風味也有著較大差異。例如在羅馬諾干酪、帕瑪森干酪和波蘿伏洛干酪這三種意大利品種中,羅馬洛干酪游離脂肪酸是最高的,波蘿伏洛干酪處于中間水平,帕瑪森是最低的[19]。還有研究表明,丁酸是羅馬洛干酪和波羅夫洛干酪中很重要的風味化合物,它也是衡量這兩種干酪風味強度和風味特征的最重要的指標之一[20]。Atasoy等[21]研究表明,風味最好的切達干酪含有4.5~5.0 mg/kg丁酸和2.0~2.5 mg/kg己酸,許多研究已經表明低脂切達干酪缺乏典型風味并且有較低濃度的游離脂肪酸,這證明了游離脂肪酸對切達干酪風味有很重要的作用[22]。
1.3 檸檬酸分解和糖酵解
芳香化合物的形成與檸檬酸鹽和乳糖的代謝也有關,代謝途徑如圖5所示。檸檬酸是牛乳中的微量成分,能被乳酸乳球菌代謝產生乙酰乳酸、二乙酰和3-羥基丁酮。丁二酮乳酸菌和明串珠菌也可將其分解成CO2和丁二酮等碳基化合物,這些化合物都是組成干酪風味的必要成分[23]。然而值得注意的是,嗜熱乳酸菌通常是檸檬酸鹽陰性菌。

圖5 與干酪風味物質形成相關的檸檬酸和乳糖轉化途徑
干酪絕大部分乳糖隨乳清排出,剩余乳糖經乳酸菌代謝生成乳酸,可形成強烈的乳酸風味。這種乳酸有抑菌作用,可以改變干酪的pH,降低干酪中氧化還原電位,為酶促反應的進行提供有利條件[24]。乳糖作為風味物質來源,先被發酵劑細菌(乳酸菌)水解成葡萄糖和半乳糖(發酵劑為乳球菌時為6-磷酸-己酮糖),然后葡萄糖通過糖酵解(EMP)途徑氧化為丙酮酸。丙酮酸是形成短鏈風味化合物的前體,如乙酸、二乙酸、丁酸、丙酸、乙醇、乙醛和3-羥基-丁酮。半乳糖則被半乳糖陽性發酵劑和明串珠菌通過半乳糖降解途徑生成6-磷酸-己酮糖,而乳球菌通過另外一種途徑將半乳糖降解為甘油醛3-磷酸-甘油醛[25]。乳糖和乳酸的代謝在大部分的干酪中屬于基本代謝類型。乳酸異構化作用對干酪風味的影響較小,但乳糖在丙酸菌的作用下會生成丙酸和乙酸,較大程度上影響干酪的整體風味。另外在干酪成熟期間,乳酸會繼續轉化為丙酮,賦予干酪水果味和不愉悅的風味[24]。乳酸也是干酪中許多微生物生長的重要底物,其進一步代謝也是影響干酪風味的主要因素。
2 乳酸菌在干酪風味物質形成過程中的作用
乳酸菌是一種革蘭氏陽性菌,由于其重要的生理功能和生化作用,已被廣泛用于發酵食品尤其是發酵乳制品的生產中。干酪在成熟過程中的微生物群主要由乳酸菌為主,根據其作用可分為兩大類:發酵劑乳酸菌(starer lactic acid bacteria,SLAB,如切達干酪的嗜溫乳球菌)和非發酵劑乳酸菌(non-starer lactic acid bacteria,NSLAB,如乳桿菌),如圖6所示。發酵劑乳酸菌和非發酵劑乳酸菌相互作用主導著干酪風味物質生成的代謝途徑[26]。因此,將分別對不同種類的乳酸菌在干酪風味物質形成過程中的作用進行綜述。

圖6 在制作干酪和成熟過程中發酵劑乳酸菌和非發酵劑乳酸菌的相互作用導致風味形成的生化途徑
2.1 發酵劑乳酸菌在干酪風味物質形成過程中的作用
干酪發酵劑也可根據其作用被分為兩組:主發酵劑和附屬發酵劑。主發酵劑是指能夠代謝原料乳中的乳糖生成乳酸或乳酸鹽的發酵劑,主要目的以酸化為主,由嗜溫型或嗜熱型的乳酸菌組成。例如制作荷蘭艾達姆干酪和高達干酪的發酵劑主要由同型乳酸發酵的乳酸乳球菌以及能夠進行檸檬酸代謝的乳酸乳球菌和明串珠菌構成[27]。附屬發酵劑為選定的干酪相關微生物菌株,將其添加到干酪牛乳中能夠改善干酪感官品質,促進并協調干酪風味物質及色素的形成。總而言之,干酪發酵劑乳酸菌主要的菌種有乳酸球菌屬、嗜熱鏈球菌屬、明串珠菌屬、片球菌屬、腸球菌屬的種、亞種及變種等;另外,某些特殊的干酪為了能形成特有的組織狀態還需要添加丙酸菌。
如圖6所示,發酵劑乳酸菌先利用乳糖作為碳源生成乳酸,檸檬酸是一些乳酸菌的次要能源,已知三種類型的乳制品發酵劑能夠吸收檸檬酸并將其代謝為額外的風味化合物:乳酸乳桿菌、雙乙酰乳酸乳球菌和明串珠菌屬[28]。檸檬酸和乳糖的共同代謝會產生CO2和C4芳香化合物,例如二乙酰(黃油味)[28]。不同種類的發酵劑乳酸菌在干酪中酪蛋白代謝途徑也有很大作用。劉瑛等[29]用甘肅天祝新鮮牦牛乳為原料分別添加嗜溫、嗜熱和混合發酵劑制作硬質干酪,結果表明對蛋白降解程度越大的發酵劑制作的干酪越容易產生苦味,其中,嗜熱發酵劑對干酪蛋白降解程度最大,混合發酵劑次之,嗜溫發酵劑最小。
支鏈酮酸的脫羧作用[圖3(3)]會產生具有麥芽糖味、巧克力味及極低氣味閾值的醛類物質,尤其3-甲基丁醛是許多硬質和半硬質干酪的關鍵風味化合物,這是由于亮氨酸能夠被干酪微生物群有效分解。然而一般認為,只有乳球菌、乳桿菌、鏈球菌、食肉桿菌和腸球菌等乳酸菌涉及干酪中3-甲基丁醛的形成[30]。目前已有研究表明乳酸菌中可能有兩種分解代謝亮氨酸生成3-甲基丁醛的途徑[30]:一是通過α-酮酸脫羧酶(KADC)直接使α-酮酸脫羧而生成,例如德氏乳桿菌亞種(L.delbrueckiisubsp. lactis)CNRZ 207;另外一種是通過α-酮酸脫氫酶(KADH)的間接途徑,例如瑞士乳桿菌(L.helveticus)CNRZ 32和糞腸球菌(E.faecalis)10C1。在馬爾他芳香卡諾桿菌中,兩種代謝途徑都存在并且起作用。同時由于3-甲基丁醛在特定干酪風味中的重要性,已經提出了許多策略來控制其生產。
有些研究已經將特定的發酵劑添加到干酪中以控制獨特風味的形成。Eugster等[31]在干酪添加乳酸片球菌(Pediococcus acidilactici)FAM18098成熟發酵后發現,該菌株將精氨酸轉化為鳥氨酸、分解代謝絲氨酸和蘇氨酸生成α-氨基丁酸和丙氨酸,乙酸鹽含量也高于不添加該菌株的干酪對照組,顯著影響干酪成熟和風味質構。此外,Hickey等[32]為了評估發酵劑乳酸菌對切達干酪脂肪水解的作用,將添加和不添加發酵劑的牛奶發酵成熟進行實驗對比,結果表明添加發酵劑的干酪能主動產生游離脂肪酸且含量明顯高于無發酵劑的干酪,這表明發酵劑乳酸菌酯酶是干酪中脂肪分解的主要因素。此外,乳酸菌酯酶不僅能催化乳脂甘油酯的水解以釋放游離脂肪酸,還會通過轉移酶反應將甘油酯和醇合成酯。總而言之,乳酸菌的酯酶影響干酪的脂肪分解和酯風味,且對干酪風味的影響可以通過控制酯酶的量、調節醇的利用率和增加乳脂的甘油單酯和甘油二酯含量來控制。
2.2 非發酵劑乳酸菌在干酪風味物質形成過程中的作用
與發酵劑乳酸菌相比,干酪中的非發酵劑乳酸菌源于原料乳和加工環境[33]。非發酵劑乳酸菌屬于外來細菌且產酸能力較弱,但有研究表明,干酪中某些固有的獨特風味可能是天然微生物群而不是發酵劑乳酸菌的產物,而且盡管非發酵劑乳酸菌在凝乳后的濃度可能較低(102~103CFU/g),但在成熟3~9個月后,其種群數量會緩慢增加約4~6個數量級[34]。如圖6所示,尤其是在硬質和半硬質干酪(例如切達干酪和埃門塔爾干酪)中,非發酵劑乳酸菌在干酪成熟期的大部分時間主導著微生物群。切達干酪微生物群中占優勢的非發酵劑乳酸菌主要由嗜溫兼性異發酵乳酸桿菌(L.casei,L.paracasei,L.rhamnosus,L.plantarum和L.curvatus)組成。Gobbetti等[34]研究表明在切達干酪中,由于非發酵劑乳酸菌的存在,蛋白水解水平顯著增加,風味強度也顯著增強。越來越多的研究從高質量發酵乳中篩選出性能優良的非發酵劑乳桿菌株,再添加到干酪中來加強干酪風味的產生,例如陳洋[35]將篩選的非發酵劑瑞士乳桿菌(L.helveticus)SMN2-1再添加到切達干酪的生產過程中,結果表明添加非發酵劑后的切達干酪在成熟過程中風味變化明顯,但是趨勢相同。然而,非發酵劑乳酸菌對干酪風味的作用至今仍存在爭議,一些菌株似乎能生產出高質量的切達干酪,而另一些菌株則會使干酪過酸和產生苦味,O’Sullivan等[36]研究發現,在最初的成熟階段,由于發酵劑乳酸菌的缺乏,過量的碳水化合物(乳糖和半乳糖)和檸檬酸鹽的存在可能會提供非發酵劑干酪乳酸菌過多的基質,導致瑞士類型干酪中產生過量的CO2和孔眼形成缺陷。
3 結語
干酪由于其豐富的營養價值和獨特的風味,越來越受到消費者的青睞。風味是食品感官機能的最重要指標,干酪風味的形成是一個復雜的動態生化過程,而干酪基質中存在的乳酸菌在此過程中起著關鍵作用。因此,詳細綜述了干酪風味形成的代謝途徑,在此基礎上討論乳酸菌在各個代謝途徑相互作用的機制,以期有助于在干酪生產過程中通過選擇特定菌種,賦予產品一致性的風味質構,尤其是需要長時間成熟的干酪,能夠增強其特定的風味甚至開發新風味。
猜你喜歡
語數外學習·高中版中旬(2023年2期)2023-05-10 13:26:53
語數外學習·高中版中旬(2022年5期)2022-07-13 20:47:51
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
