耕作方式對稻茬小麥幼苗莖蘗生長生理和生產力的影響
2022-03-22 06:31:28李福建徐東憶劉凱麗李春燕朱新開丁錦峰郭文善
華北農學報 2022年1期
關鍵詞:差異
李福建,徐東憶,劉凱麗,朱 敏,李春燕,朱新開,丁錦峰,郭文善
(揚州大學 江蘇省作物遺傳生理國家重點實驗室培育點,糧食作物現代產業技術協同創新中心,揚州大學小麥研究中心,江蘇 揚州 225009)
小麥苗期是生長葉片、分蘗和根系等營養器官為主的時期,同時是決定穗數和奠定大穗的重要時期[1]。研究表明,幼苗的根系長度、根數與成熟期生物量、穗數、穗粒數和籽粒產量顯著正相關[2-3]。因此,培育冬前壯苗有助于小麥安全越冬,是獲得豐產的基礎。通過農藝措施的改善能夠實現幼苗質量的提升。適宜的品種、播期、耕作和播種方式[4-6],合理的施肥和灌溉策略[7-8],激素和種子包衣的應用[9-10]均有利于調控小麥苗期的形態生長、營養吸收和生理活性,最終轉化為光合物質生產能力。其中,耕作方式可以通過農機具的物理機械作用改變秸稈分布和土壤結構,以調節土壤水分分布、氣體組成、熱循環和養分有效性[11]。土壤性質的變化所創造的獨特環境對調控作物出苗和生長具有重要意義。研究表明,深翻耕改善了土壤孔隙狀況,促進了根系下扎,利于根系生長,為冬前壯苗奠定了基礎[12]。Li等[13]認為深松或深耕改善了土壤滲透并在更長時間內保持土壤水分,提高了作物水分利用效率。此外,翻耕和旋耕將秸稈與土壤混合,有利于冬前土壤溫度的提高,加快了小麥生長[14]。這些研究明確了秸稈還田下通過改善土壤理化狀況進而提高幼苗質量。然而,稻麥輪作系統中由于水稻秸稈還田量大且還田質量難以保證,加之水稻秸稈碳氮比高,還田后易發生與下茬作物爭奪養分等情況,這些都影響小麥根系和分蘗生長,造成小麥苗情質量差,已成為制約稻茬小麥增產的關鍵因素之一[15]。因此,明確適宜的耕作措施促進稻茬小麥壯苗有助于群體穩健發展和高產形成。
分蘗是指產生于分蘗節上的分枝,是小麥的重要生物學特性之一[1]。前人關于對分蘗特性的研究已有大量報道。Cai等[9]研究發現,小麥分蘗發生受到多種激素共同調控。郝艷玲等[16]在西南麥區的研究表明,小麥單株分蘗之間關系決定其單株形態特征,直接影響產量性狀的形成。王思宇等[17]研究表明,晚播下分蘗持續時間延長,但無效分蘗多,分蘗消亡率增加,且播期越晚,無效分蘗越早消亡,成穗率隨播期推遲而下降,適度早播有助于提高分蘗數和成穗率,進而提高有效穗數以達到高產。可見,生長健壯的分蘗不僅可以提高成穗率,還可提升穗生產力。
雖然已有較多的文獻報道了分蘗的發生與調控,但關于耕作對小麥主莖和分蘗生長生理影響的研究還鮮有報道。本試驗在水稻秸稈全量還田條件下,研究了不同耕作方式對小麥苗期形態指標、主莖和分蘗生理及其成穗產量構成的影響,以期明確不同苗質間生理特性的差異,為稻茬小麥壯苗培育和高產栽培提供理論與技術依據。
1 材料和方法
1.1 試驗地點及供試材料
試驗于2019—2020年在黃淮麥區南片的江蘇睢寧稻麥科技綜合示范基地(33°53′N,117°56′E)進行。試驗田前茬為水稻,半喂入式水稻收割機收割后秸稈全量還田,還田量約為12 000 kg/hm2。土壤類型為輕壤土,播種前0~20 cm土壤相對含水量為83.62%,含全氮1.00 mg/g、堿解氮96.79 mg/kg、速效磷20.89 mg/kg、速效鉀145.42 mg/kg、有機質19.59 g/kg。選用多穗型小麥品種遷麥088,由江蘇省農業科學院農業資源與環境研究所提供。
1.2 試驗設計
試驗采用單因素隨機區組設計,設置耕翻(PR)、旋耕(RR)和免耕(NT) 3種耕作方式。耕翻處理流程為鏵式犁深翻1遍后旋耕機旋耕2遍;旋耕處理流程為旋耕機旋耕2遍;免耕處理流程為直接采用播種機播種。鏵式犁和旋耕機牽引動力為64 kW的LX954型東方紅拖拉機,作業深度分別為20~25 cm,8~10 cm。播種日期為2019年10月18日,采用2BMQF-7/14型條帶免耕寬幅施肥播種機播種。該播種機可一次性完成旋耕(深度約5 cm)、施基肥、寬幅條播、蓋籽、鎮壓等作業,播種行距35 cm、帶寬10 cm。每個小區面積為420 m2,3次重復。3葉期以基本苗為225×104株/hm2定苗。施氮量為240 kg/hm2,基肥∶壯蘗肥∶拔節肥∶孕穗肥為5∶1∶2∶2;磷、鉀肥施用量均為120 kg/hm2,基肥∶追肥為5∶5。其中基肥于播種前施用,壯蘗肥于4.5葉期施用,拔節肥于倒2.5葉期施用,孕穗肥于旗葉露尖施用。其他管理措施同當地高產栽培。
1.3 測定項目與方法
1.3.1 單株莖蘗數、單蘗葉面積和干物質量 在3葉期,每個小區隨機選定長勢一致的60個單株標記,每個葉齡期對每株的分蘗進行標記。按不同蘗位分蘗出現順序分別掛牌標為主莖(0)、第一分蘗(Ⅰ)、第二分蘗(Ⅱ)、第三分蘗(Ⅲ),供試驗取樣。在越冬始期取標記植株20株,洗凈晾干后調查單株分蘗狀況,分別取下主莖和不同分蘗的葉片,用葉面積儀(美國LI-COR,LI-3000C型)測定單株和單蘗葉面積。測定后樣品按莖和葉分開,105 ℃殺青1 h,80 ℃烘干至恒質量,測定單株和單蘗干物質量。
1.3.2 SPAD值、含氮量、可溶性糖含量 烘干葉片研磨混勻,按照H2SO4-H2O2靛酚藍比色法測定含氮量,采用蒽酮法測定可溶性糖含量。采用葉綠素儀(日本KONICA MINOLTA,SPAD-502 PLUS型)測定越冬始期主莖和各分蘗頂部全展葉SPAD值。
1.3.3 氮代謝酶、糖代謝酶和光合關鍵酶活性 于越冬始期選取各處理主莖和分蘗頂部全展葉,清洗吸干水分后液氮速凍、超低溫冰箱保存。采用冷凍混合球磨儀(德國RETSCH,MM400型)混勻研磨作為測定樣品。
參照馬新明等[18]的方法測定硝酸還原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)活性;參照Camp等[19]和賈少磊等[20]的方法測定二磷酸核酮糖羧化酶(Rubisco)和丙酮酸磷酸雙激酶(PPDK)活性;參照牛奎舉[21]的方法測定磷酸丙糖異構酶(TPI)活性;參照Bilska-Kos等[22]的方法測定蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS-Ⅱ)活性。
1.3.4 穗粒數、單粒質量和單穗產量 于成熟期每個小區取10株標記樣本,測定其主莖和各分蘗穗部的穗粒數、單粒質量和單穗籽粒產量。
1.4 數據分析
采用Excel 2016建立數據庫,用SPSS 19.0 軟件進行統計分析,利用Origin 2018軟件作圖。
2 結果與分析
2.1 耕作方式對單株莖蘗數、葉面積和干物質量的影響
由圖1可知,小麥單株莖蘗數、葉面積和干物質量在耕作方式間均表現為:PR>RR>NT。其中PR的單株莖蘗數分別比RR和NT高1.5%,12.8%,處理間均存在顯著差異。PR的單株葉面積和干物質量分別達到94 cm2和787 mg。PR和RR的單株葉面積無顯著差異,但與NT差異顯著;PR的單株干物質量與RR和NT差異顯著。
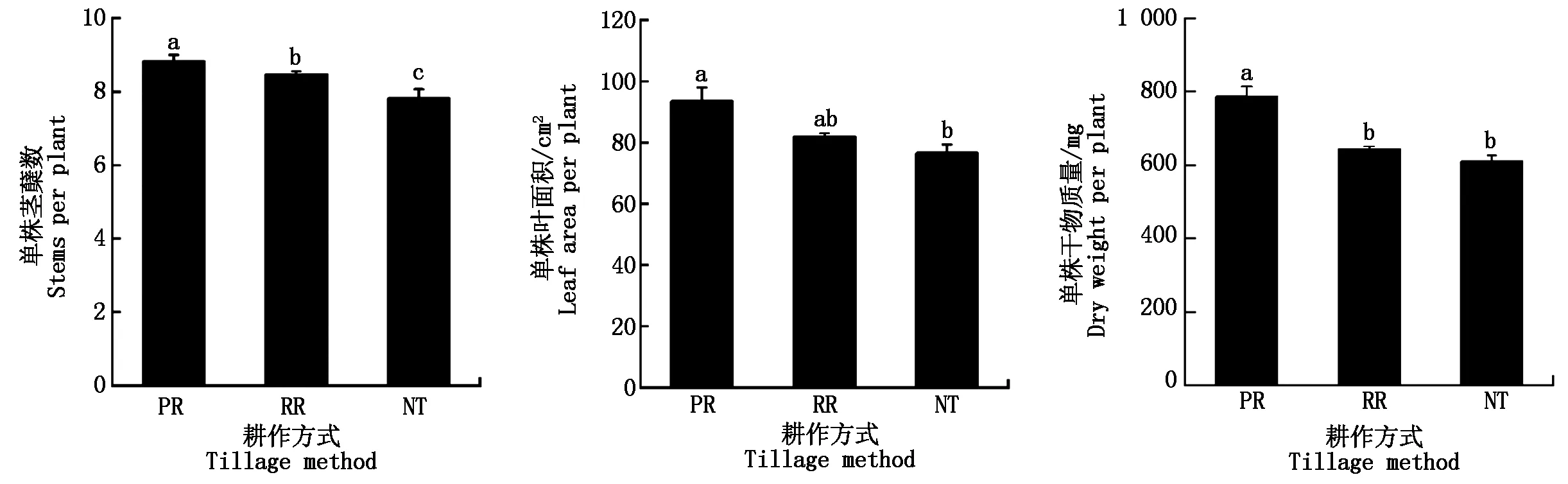
不同小寫字母表示處理間差異顯著(P<0.05)。Different small letters mean significant difference among treatments at 0.05 level.
2.2 耕作方式對主莖和各分蘗葉面積、干物質量、葉片氮含量和可溶性糖含量的影響
由表1可知,耕作方式極顯著影響葉片中的氮含量和可溶性糖含量,但未顯著影響單莖葉面積和干物質量。各指標在不同耕作方式間均表現為PR>RR>NT。單莖葉面積、干物質量、葉片氮含量和可溶性糖含量在不同蘗位間的差異均達極顯著水平。隨著蘗位的提高,各指標呈現逐步下降的趨勢。耕作方式和蘗位兩因素互作顯著影響單莖葉面積和干物質量,極顯著影響可溶性糖含量。不同耕作方式的單莖葉面積在主莖上差異顯著;PR與RR、RR與NT在第一分蘗上差異不顯著,但PR與NT差異顯著;在第二和第三分蘗上差異不顯著。不同耕作方式間主莖的單莖干物質量和可溶性糖含量表現為PR和RR間無顯著差異,但均顯著高于NT。第一、第二和第三分蘗的干物質量在耕作方式間差異均不顯著。可溶性糖含量在第一和第二分蘗上均表現為PR和RR顯著高于NT,而第三分蘗上表現為PR顯著高于RR和NT。這說明PR相比NT顯著提高了小麥幼苗主莖和第一、第二、第三分蘗的葉片可溶性糖含量,而RR僅提高了主莖和第一、第二分蘗的葉片可溶性糖含量,并且PR和RR均明顯提升了主莖和各分蘗的單莖葉面積和干物質量。
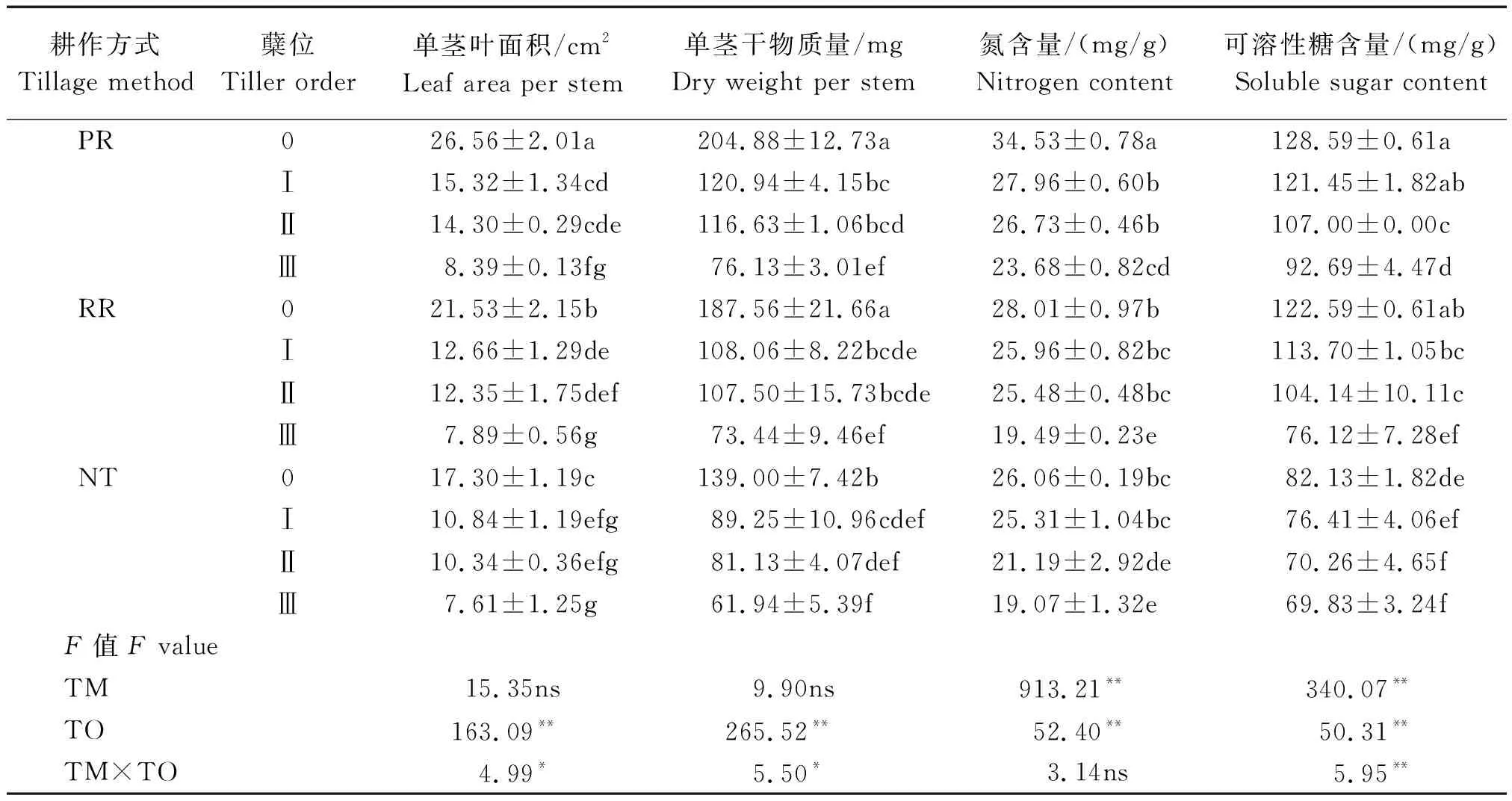
表1 耕作方式對主莖和各分蘗葉面積、干物質量、葉片氮含量和可溶性糖含量的影響Tab.1 Effects of tillage methods on leaf area,dry weight,leaf nitrogen content,and soluble sugar content of main stem and tillers
2.3 耕作方式對主莖和各分蘗氮代謝酶活性的影響
小麥苗期頂部全展葉的硝酸還原酶(NR)、谷氨酰胺合成酶(GS)和谷氨酸合成酶(GOGAT)活性隨著蘗位的升高總體呈現下降的趨勢(圖2)。其中,NR在不同蘗位間均表現為PR顯著高于RR和NT,RR和NT間差異不顯著。GS在主莖和第三分蘗上表現為PR略高于RR,二者均顯著高于NT;第一和第二分蘗上表現為PR顯著高于RR和NT,RR與NT僅第二分蘗上差異顯著。GOGAT在主莖和第二分蘗上表現為PR顯著高于RR和NT,RR與 NT在第二分蘗上差異顯著;第一和第三分蘗上PR和RR無顯著差異,但均顯著高于NT。說明,相比NT,PR可顯著提升小麥幼苗主莖和第一、第二和第三分蘗頂部全展葉的NR、GS和GOGAT活性,而RR僅部分提升了GS和GOGAT活性。
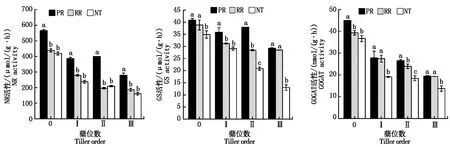
不同字母表示同一蘗位不同耕作處理間差異顯著(P<0.05)。圖3—4同。Different small letters mean significant difference at 0.05 level among tillage methods at the same tiller order.The same as Fig.3—4.
2.4 耕作方式對主莖和各分蘗光合酶活性和葉綠素相對含量(SPAD值)的影響
由圖3可知,小麥苗期頂部全展葉的二磷酸核酮糖羧化酶(Rubisco)、丙酮酸磷酸雙激酶(PPDK)和磷酸丙糖異構酶(TPI)活性和葉綠素相對含量(SPAD)均隨著蘗位的升高呈下降趨勢。相比RR和NT,PR顯著提高了主莖的Rubisco、TPI活性和SPAD,RR和NT間無顯著差異。PR第一分蘗的Rubisco活性分別比RR和NT高20.2%,43.13%,處理間差異顯著;PR第二和第三分蘗的Rubisco活性與RR差異不顯著,但顯著高于NT。PR相比NT顯著提高了第一、第二和第三分蘗的TPI活性,而RR僅顯著提高了第二分蘗的TPI活性。PR第一和第三分蘗的SPAD均顯著高于RR和NT;PR第二分蘗的SPAD與RR無顯著差異,但均顯著高于NT。PR在主莖和各分蘗上的PPDK活性均顯著高于NT;在主莖、第二和第三分蘗上顯著高于RR;而RR在主莖、第一和第二分蘗的PPDK活性與NT差異顯著。這說明,相比NT,PR同步提升了小麥苗期主莖、第一、第二和第三分蘗頂部全展葉的Rubisco、PPDK、TPI活性和SPAD,而RR僅主要提升了主莖、第一和第二分蘗的PPDK活性。

圖3 耕作方式對主莖和各分蘗頂部全展葉二磷酸核酮糖羧化酶(Rubisco)、丙酮酸磷酸雙激酶(PPDK)和磷酸丙糖異構酶(TPI)活性和葉綠素相對含量(SPAD)的影響Fig.3 Effects of tillage methods on the activities of Rubisco,PPDK and TPI,and SPAD of the top full expanding leaves in main stem and different tiller
2.5 耕作方式對主莖和各分蘗糖代謝酶活性的影響
由圖4可知,隨蘗位的提高,小麥苗期頂部全展葉的蔗糖合成酶(SS-Ⅱ)活性和蔗糖磷酸合成酶(SPS)活性呈下降趨勢。不同耕作方式在主莖和各分蘗上的SS-Ⅱ和SPS活性均表現為:PR>RR>NT;并且PR與NT不同蘗位間的SS-Ⅱ和SPS活性差異均達顯著水平。PR與RR主莖和第一分蘗的SS-Ⅱ活性差異顯著,第二和第三分蘗下無顯著性差異;而RR與NT在第一、第二和第三分蘗上SS-Ⅱ活性差異顯著。3種耕作方式的SPS活性在第三分蘗上差異顯著。說明PR相比于NT顯著提升了主莖和各分蘗的SS-Ⅱ和SPS活性;而RR相比NT僅顯著提升了第一、第二和第三分蘗的SS-Ⅱ活性,以及第三分蘗的SPS活性。
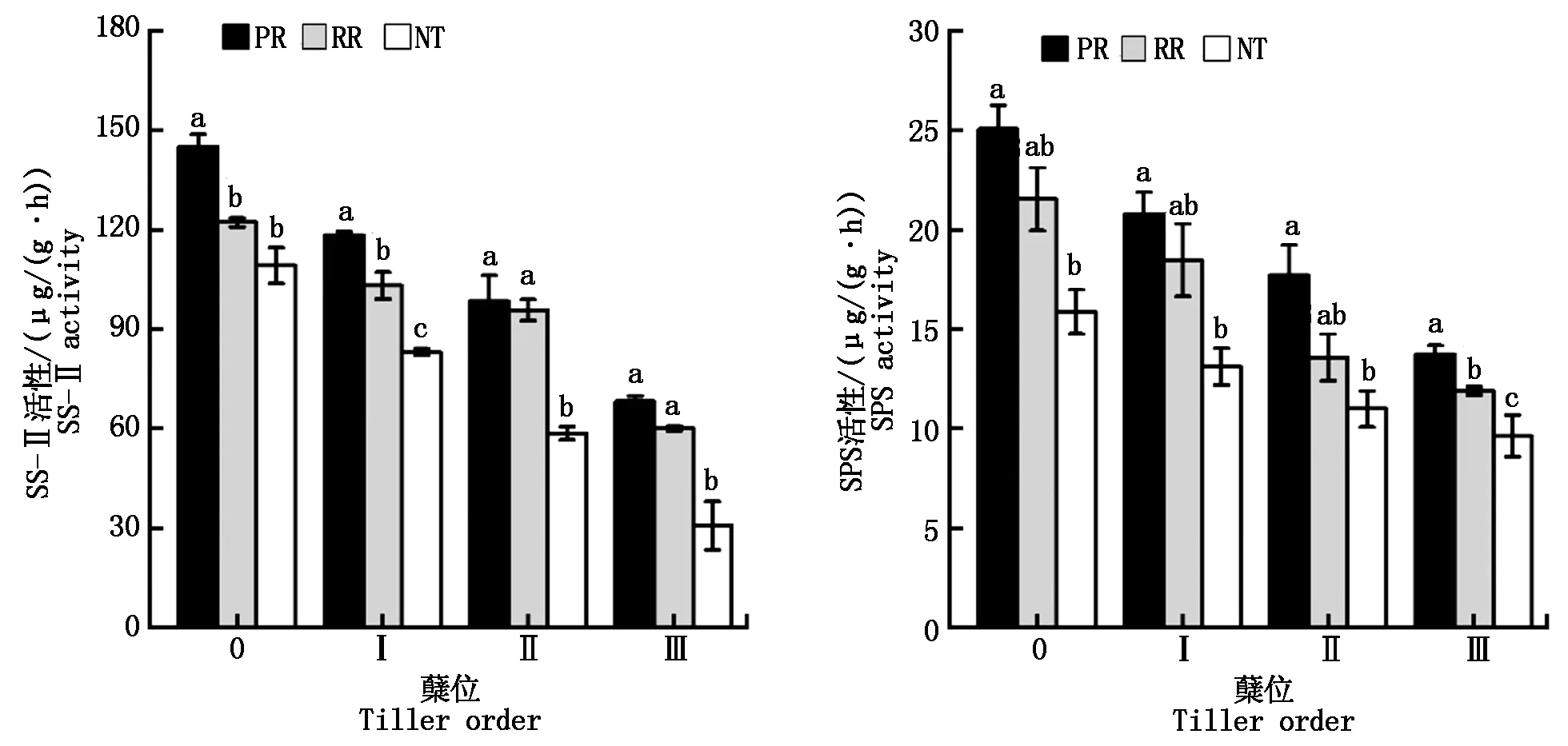
圖4 耕作方式對主莖和各分蘗頂部全展葉蔗糖合成酶(SS-Ⅱ)和蔗糖磷酸合成酶(SPS)活性的影響Fig.4 Effects of tillage methods on the activities of SS-Ⅱ and SPS of the top full expanding leaves in main stem and different tiller
2.6 耕作方式對主莖和各分蘗單穗產量的影響
由表2可知,耕作方式顯著影響穗粒數、單粒質量和單穗產量,各指標在耕作方式間均表現為PR>RR>NT。蘗位極顯著影響穗粒數和單穗產量,對單粒質量影響不顯著;隨著蘗位的提高,各產量指標均呈下降趨勢。穗粒數受耕作方式和蘗位間互作的顯著影響。PR和RR處理主莖的穗粒數分別比NT提高了12.4%,7.5%;PR處理第一和第二分蘗穗粒數顯著高于RR和NT,第三分蘗穗粒數在耕作方式間差異不顯著。PR和RR主莖和各分蘗上的單粒質量和單穗產量差異較小,但明顯高于NT。說明相比NT,PR和RR增加了主莖、第一和第二分蘗的穗粒數、單粒質量和單穗產量。
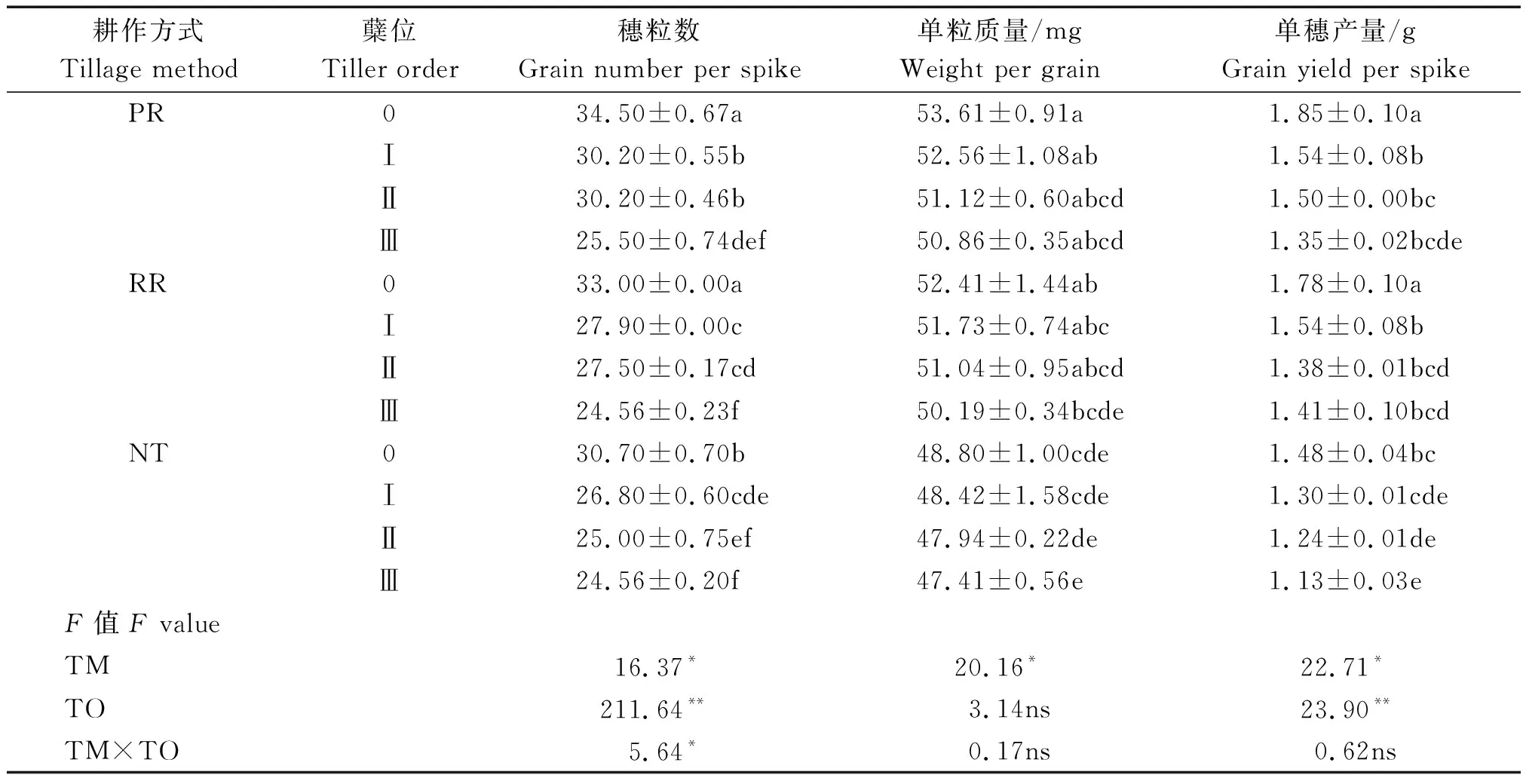
表2 耕作方式對主莖和各分蘗單穗產量性狀的影響Tab.2 Effects of tillage methods on yield components of the single spike in main stem and different tiller
3 討論與結論
3.1 耕作方式對小麥苗期主莖和分蘗生理活性的影響

3.2 耕作方式對小麥苗期主莖和分蘗生長的影響
前人關于耕作方式對小麥苗期生長的影響,梅晶晶等[30]在黃淮平原玉米秸稈還田條件下的研究認為,深耕相比旋耕顯著提高了小麥苗期的莖蘗數、葉面積指數以及單株次生根數。而吳鵬等[31]和李朝蘇等[32]在稻秸稈全量還田下研究認為,免耕相比淺旋耕和深旋耕顯著降低了主莖第一葉位的分蘗缺位率,提高了小麥幼苗的單株分蘗數,增大了葉面積和干質量。由此可知,不同生態條件和土壤特性下,不同耕作方式對小麥幼苗的生長發育調控存在較大差異。本研究在黃淮麥區南部輕壤土地區開展,小麥播種到越冬期降雨較少,水稻秸稈全量深翻還田,增加了土壤蓄水能力。本研究結果表明,相比RR和NT,PR顯著提高了小麥苗期的單株莖蘗數和單株干物質量,也提高了單株葉面積。說明PR處理更有利于為小麥提供適宜的生長環境,促進其生長發育。耕翻方式通過犁耕加旋耕,較好的將秸稈均勻分布在20 cm土層,降低了土壤的緊實程度,綜合增強了土壤保肥蓄水能力和小麥苗期根系的吸收能力,有利于提高小麥的播種質量和幼苗質量[12,33]。同時,秸稈翻耕還田也可調節分蘗節微環境溫度,利于緩解溫度變化對小麥的影響,有利于減少冬季凍害[34]。旋耕處理攪動土壤較淺,未能完全將秸稈深埋,可能由于秸稈還田質量不高,過多的秸稈影響根系生長導致雖對幼苗質量有部分提升,但效果不佳。
趙會杰等[26]研究表明,小麥葉片中的可溶性糖和14C同化物分配存在明顯的主莖優勢現象,光合產物主要積累于主莖中,其分配率隨蘗位的升高逐漸降低。本研究表明,小麥苗期單莖葉面積和干物質量及葉片中的氮和可溶性糖在主莖與各分蘗間存在顯著差異,隨著蘗位的提高,各指標呈現逐步下降的趨勢。研究表明,低位分蘗和高位分蘗間存在限制資源的競爭,早發分蘗通常會攔截更多的光合有效輻射并抑制晚發分蘗生長,因此,低位分蘗在生長早期的不對稱競爭中占據主導地位[35]。這主要與小麥葉蘗同伸規律有關,當第一個分蘗發生時,主莖已有3張全展葉片,之后主莖新生1張葉片便多發生1個分蘗,即主莖與第一分蘗相差3個葉齡,第一分蘗、第二分蘗和第三分蘗分別相差1個葉齡[1,36]。因此,主莖和第一、第二分蘗具有溫光資源利用的先發優勢,這對于生理活性的提高、營養物質的積累和光合物質生產十分重要。所以,分蘗調控和低位分蘗的利用是建立高產高效群體的關鍵因素。
前人研究了施肥、密度和品種等的分蘗特性差異,結果表明,不同措施均對主莖與分蘗具有一定的調控效應[26,37]。本研究結果表明,耕作方式對主莖和分蘗發育也具有調控效應,表現為相比NT,PR增加了小麥苗期主莖和各分蘗葉片中糖、氮含量,且具有較高的單莖葉面積和干物質量。這主要得益于PR下主莖和第一、第二分蘗下氮代謝酶活性的提高,促進了氮素積累、轉運和同化能力。同時,葉片中SS-Ⅱ和SPS活性的提高促進了小麥苗期可溶性糖的合成和運輸。糖作為光合作用最直接的產物,糖含量的提高不僅反映了葉片一端同化物供給力的情況以及庫端對同化物轉運利用的能力,并作為滲透調節物質,有利于提高小麥的越冬抗寒能力[38-39]。同時光合效率的增強,提高了物質轉化能力;葉面積的增大實現了光合“源”的擴大,進而提高了物質生產能力。主莖和第一、第二分蘗生產能力的提升促進了PR下單株生產力的提高,有利于形成小麥壯苗越冬。
3.3 耕作方式對小麥主莖和分蘗的產量的影響
冬前壯苗對于后期健壯個體和健康群體發育起著決定性作用[40]。研究表明,小麥越冬始期的幼苗更多的分蘗利于有效穗數的增加,更高的單莖葉面積和生物量利于單穗產量的提升,指示壯苗生長有助于增產[41]。而壯蘗的培育對于幼穗分化和產量形成起著數量和質量的雙重作用[36,40]。通過限制低位分蘗生長、強化早發分蘗發育,構建良好的穗群組成、促進單莖光合生產可以提升旱茬小麥產量潛力[9]。本研究結果表明,PR和RR通過增強小麥苗期主莖和第一、第二分蘗糖氮代謝水平和光合強度(光合面積大、光合能力強),提高了冬前的生物量,為“強源”奠定了良好的物質基礎。壯蘗早發促進了小麥分蘗節大維管束數的分化,這為壯稈大穗打下了基礎,同時利于植株體內物質運輸通道的建立,給無機養分向地上部運輸、光合產物的轉運創造條件[42]。本研究中PR下主莖和一蘗、二蘗主要通過增加穗粒數,穩定單粒質量,促進了單穗產量的提升,而RR僅提升了主莖和第一分蘗的生產力。Yang等[35]的研究結果表明,花后15 d的維管束數目與單穗產量呈顯著正相關,通過促進主莖和低位分蘗維管束數量的提升,加快了灌漿速率,其粒數和單穗產量較高。另外,不同耕作方式與蘗位間互作單粒質量差異并不明顯,可能是由于單粒質量受到小麥生育后期群體大小和葉片光合能力的影響。本研究主要探討了不同耕作方式下苗期主莖和分蘗生長生理及其與單穗生產力的關系,針對不同耕作下的莖蘗構成與組成比例,以及莖蘗動態發育有待進一步研究。
結果表明,相比免耕,耕翻方式顯著提升了小麥苗期主莖和各分蘗頂部全展葉的氮代謝酶、糖代謝酶和光合酶活性,以及SPAD;耕翻下小麥苗期主莖和第一、第二分蘗的葉片氮含量和可溶性糖含量,單莖葉面積和干物質量也明顯較高。而旋耕對于小麥苗期生理生長的提升較為有限。這說明耕翻通過優化小麥苗期主莖和分蘗的氮糖代謝能力、提高光合生理水平,促進小麥對養分的吸收、利用和同化能力,利于形成冬前壯苗。同時,冬前壯蘗對于增加穗粒數、單粒質量和單穗產量十分有利,耕翻和旋耕通過提高主莖、第一和第二分蘗的穗粒數增加單穗產量。因此,培育壯苗高產群體,需要通過不同的栽培管理措施提高主莖和第一、第二分蘗的生理活性和生產能力,以實現江蘇淮北稻茬小麥產量進一步提升。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
