利用分子標記輔助選擇技術培育抗螟蟲轉基因雜交稻恢復系
2022-04-06 02:32:50劉開強李孝瓊王小姣李林娟郭嗣斌
西南農業學報 2022年2期
關鍵詞:水稻
劉開強,李孝瓊,韋 宇,陳 穎,王小姣,李林娟,郭嗣斌
(廣西農業科學院水稻研究所/廣西水稻遺傳育種重點實驗室,南寧 530007)
【研究意義】水稻是我國的主要糧食作物之一,其產量和品質對保障我國糧食安全至關重要[1-2]。螟蟲(二化螟、三化螟和稻縱卷葉螟等)是水稻的主要害蟲,對世界水稻生產造成的損失巨大,其中,二化螟在我國稻區分布較廣,年防治面積達1500萬hm2,年防治費用約6.5億美元,年產值損失8.4億美元[3-4]。使用化學殺蟲劑防治螟蟲效果雖然較好,但耗費巨大,而且還會造成環境污染,其農藥殘留甚至威脅人類健康。培育抗螟蟲水稻新品種是最經濟有效的水稻螟蟲防治措施,可在提高水稻產量的同時保護生態環境安全[5]。但我國的水稻品種資源多為低抗或中抗螟蟲材料,抗性低且不穩定,高抗螟蟲品種資源較少,采用常規雜交育種方法很難培育出抗螟蟲水稻品種,而利用轉基因技術改良水稻抗蟲性已成為抗蟲育種的新方法[6]。在前人研究基礎上,以轉Bt基因水稻材料為供體與農藝性狀優良的水稻品系雜交、回交及多代自交,通過分子標記輔助選擇(Molecular marker-assisted selection,MAS)技術結合田間農藝性狀篩選,是選育兼具優良農藝性狀和抗蟲新品種的有效途徑[7-10]。因此,通過MAS技術進行抗螟蟲水稻新恢復系選育,對我國雜交稻抗性育種具有重要意義。【前人研究進展】MAS可有效縮短育種周期和提高育種效率,在水稻、玉米和小麥等糧食作物及多種經濟作物上已廣泛應用[11-12]。蘇云金芽孢桿菌(Baclillusthuringiensis)蛋白(Bt蛋白)對鱗翅目昆蟲具有專一毒性,對人類無毒害作用。Fujimoto等[13]利用轉基因技術將cryIA(b)基因轉入粳稻,并使crylA(b)蛋白成功表達,從而培育出抗螟蟲粳稻品種。Cheng等[14]通過農桿菌介導的轉化獲得2600個轉cryIA(b)和cryIA(c)基因水稻植株,并已證實這些基因能在R0和R1代中整合、表達和遺傳。Tu等[15]在Bt明恢63植株中檢測到Bt融合蛋白cryIA(b)/cryIA(c)含量為20.0 ng/mg;對轉基因雜交組合Bt汕優63進行田間卷葉蟲和黃莖螟抗性試驗,發現在不降低水稻產量的情況下,轉基因雜交稻高抗卷葉蟲和黃莖螟。Wang等[16]利用轉Bt基因水稻品系KMD1和KMD2與常規水稻品種雜交,并鑒定其后代F1、BC1和F2群體具有抗蟲遺傳性,其中Bt水稻對螟蟲的抗性為顯性遺傳,易于在雜交水稻生產中利用。高方遠等[17]將優質抗稻瘟病水稻恢復系成恢177與轉基因水稻Bt明恢63雜交并回交1次,采用PCR分析、試紙條檢測和田間抗性鑒定相結合的方法,育成兼具轉基因抗蟲性和稻瘟病抗性的水稻新恢復系Bt5198。李榮田等[18]、孔夢瑩等[19]以水稻空育131為受體,利用根瘤農桿菌介導法創制了轉cry1C*和cry2A*基因的抗蟲水稻獨立系,并對不同生長發育階段Bt基因表達量進行了測定。隨著對轉基因抗蟲水稻研究的深入,我國于2009年11月通過了對轉Bt基因抗蟲水稻華恢1號和Bt汕優63的生物安全認證,轉Bt基因抗螟蟲水稻技術已日趨成熟。華中農業大學已成功將多個Bt基因(cry1Ab/c、cry1C*和cry2A*)導入生產上大面積推廣應用的三系雜交稻恢復系明恢63中,并篩選出標記基因已經丟失的抗蟲恢復系材料TT51,以其配組的雜交組合抗蟲1號和抗蟲2號已完成在多個省市的生產性試驗[20-21]。崔海瑞等[22]已育成對二化螟、三化螟和稻縱卷螟等鱗翅目害蟲具100%抗性的克螟稻,以此為種質初步育成具有推廣前景的大批秈稻新品系和新組合,并開始較大面積示范。實踐證明,以轉Bt基因抗蟲水稻材料為供體,以農藝性狀優良的水稻品系為受體,通過MAS技術結合農藝性狀篩選,可選育出兼具優良農藝性狀和抗蟲的新品系,這些抗蟲新品系均能穩定遺傳Bt抗蟲基因,并能成功表達Bt蛋白,且具有與供體親本相當的抗蟲能力[20-22]。【本研究切入點】近年來廣西水稻螟蟲發生面積不斷增加,危害加重,造成產量損失增大,某些地區局部爆發螟蟲危害產量損失達50%甚至絕收,而目前針對廣西抗螟蟲雜交稻恢復系選育的研究鮮見報道。【擬解決的關鍵問題】通過雜交、回交結合MAS技術將轉Bt基因水稻中的抗蟲基因cry1C*導入廣西骨干恢復系中,培育適合廣西稻區推廣應用的抗螟蟲轉基因水稻恢復系,為雜交水稻抗蟲育種提供參考依據及基礎材料。
1 材料與方法
1.1 試驗材料
試驗于2017—2020年在廣西南寧市進行。含抗蟲基因cry1C*和與目標基因連鎖的抗除草劑基因(bar基因)的抗螟蟲轉基因材料T1C-19由華中農業大學作物遺傳改良國家重點實驗室提供。其他水稻材料包括廣西雜交稻骨干恢復系親本廣恢998、桂恢1561和桂恢110,以及利用這些恢復系回交轉育而成的抗螟蟲轉基因恢復系材料KH998、KH1561和KH110均由廣西農業科學院水稻研究所分子育種研究室提供。
1.2 試驗方法
1.2.1 抗螟蟲材料回交轉育 以轉基因材料T1C-19為供體,以廣西雜交稻骨干恢復系親本廣恢998、桂恢1561和桂恢110為輪回親本進行1次雜交、3次回交和連續8次自交,結合MAS和抗蟲鑒定獲得抗螟蟲轉基因恢復系材料KH998、KH1561和KH110。
1.2.2 抗螟蟲材料及雜交組合的農藝性狀考查和抗蟲性鑒定 2017年晚季配制雜交組合野香A/KH998和天豐A/KH998,2018年早季以特優63為對照(CK1)種植抗蟲轉基因雜交組合野香A/KH998和天豐A/KH998。2020年早季配制雜交組合華浙2A/KH110、天豐A/KH1561、華浙2A/KH1561和天豐A/KH110,2020年晚季以天優華占為對照(CK2)種植抗蟲轉基因雜交組合華浙2A/KH1561、華浙2A/KH110、天豐A/KH1561和天豐A/KH110。試驗設2次重復,每份材料種5行,每行種10株,種植密度為18.9 cm×16.5 cm。收獲時,取行中間8株考察株高、每株有效穗數、穗長、每穗實粒數、每穗總粒數、結實率、千粒重和單株重等性狀,取8株的平均值作為分析數據。
采用室內離體接蟲鑒定法結合室外自然鑒定法進行轉基因材料抗蟲鑒定。室內離體接蟲鑒定法具體操作如下:于拔節期從稻株上取莖稈,切成5.0 cm長的莖段,放入養蟲管,每管放5個莖段,接入20頭一齡三化螟,置于人工氣候箱,5 d后統計三化螟死亡率,3次重復。
1.2.3 抗螟蟲材料的分子標記鑒定 水稻全基因組DNA通過改良CTAB法[23]抽提獲取。根據cry1C*基因的序列設計引物檢測水稻基因組DNA中是否含有該基因。參考Tang等[24]的方法進行PCR擴增,擴增引物為F:5′-TTCTACTGGGGAGGACATCG-3′,R:5′-CGGTATCTTTGGGTGATTGG-3′,擴增片段長度為602 bp。PCR反應體系20.0 μL:DNA模板30~50 ng,10×Buffer 2.0 μL,2 mmol/L dNTP 1.5 μL,25 mmol/L MgCl22.0 μL,10 μmol/L引物(F/R)各0.5 μL,加ddH2O至20.0 μL;擴增程序:94 ℃預變性5 min;94 ℃ 1 min,57 ℃ 1 min,72 ℃ 1 min,進行35個循環;72 ℃ 5 min。擴增產物進行1%瓊脂糖凝膠電泳檢測。
根據bar基因序列設計引物檢測水稻基因組DNA是否含有該基因。PCR擴增引物為F:5′-GGGTCTAGAATGGAGCCCAGAACGACGCCG-3′,R:5′-GCTCGGGATCCTCAGATCTCGGTGACG-3′,擴增片段長度為561 bp。PCR反應體系20.0 μL:DNA模板50 ng,10×Buffer 2.0 μL,2 mmol/L dNTP 1.5 μL,25 mmol/L MgCl22.0 μL,10 μmol/L引物(F/R)各0.5 μL,Taq酶1 U,加ddH2O至20.0 μL;擴增程序:94 ℃預變性5 min;94 ℃ 1 min,57 ℃ 1 min,72 ℃ 1 min,進行35個循環;72 ℃ 5 min。擴增產物進行1%瓊脂糖凝膠電泳檢測。
1.2.4 抗螟蟲材料殺蟲蛋白含量檢測 于黃熟期取參試材料的新鮮綠色葉片檢測殺蟲蛋白表達量,參照cry1C平板試劑盒(美國Envirologix公司)說明進行檢測。具體方法如下:稱取20.0 mg水稻新鮮葉片,用試劑盒隨帶的抽提液進行研磨抽提,在酶標儀450 nm處讀取吸光度值,繪制標準曲線,計算樣品中的Bt蛋白含量。
1.3 統計分析
試驗數據采用Excel 2007進行統計分析。
2 結果與分析
2.1 抗螟蟲轉基因恢復系的選育歷程
抗螟蟲轉基因恢復系的選育過程見圖1。以骨干恢復系(廣恢998、桂恢1561和桂恢110)為母本,與T1C-19進行雜交,獲得F1代雜種。種植F1植株,根據株葉形態特征去除假雜種,選擇真雜種作父本與輪回親本(骨干恢復系)進行回交,獲得BC1F1代種子。種植BC1F1植株,移栽后逐株取葉片進行MAS,選取檢測結果為陽性的植株為父本與輪回親本繼續進行回交,獲得BC2F1種子;同樣在BC2F1中選取陽性單株繼續回交獲得BC3F1,最終根據分子標記檢測和農藝性狀考察結果,在BC3F1中收獲10~20株陽性植株,收取自交種。將收獲的10~20株陽性BC3F2群體各種植100苗,結合分子標記檢測結果和農藝性狀表現,最后在15個群體中選擇30個Bt陽性單株收種。種植來自BC3F2的30個BC3F3株系,每個株系50苗,苗期逐株進行PCR檢測,篩選出10個Bt基因純系。10個Bt基因純系田間自然鑒定均無螟蟲發生,根據農藝性狀表現,選擇與輪回親本差異最小的株系,再經過連續5代自交穩定后,正式定名。通過此方法選育出與廣西雜交稻骨干親本廣恢998、桂恢1561和桂恢110分別對應的抗螟蟲轉基因恢復系材料KH998、KH1561和KH110。
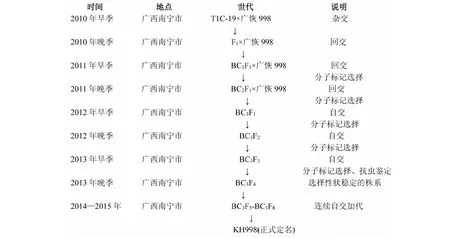
圖1 抗螟蟲轉基因恢復系KH998的選育過程Fig.1 Breeding process of insect-resistant restorer line KH998
2.2 抗螟蟲轉基因恢復系及其雜交組合的性狀表現
2016年早季種植KH998、KH1561和KH110及其原始親本廣恢998、桂恢1561和桂恢110,以T1C-19為對照(CK3),考查農藝性狀表現。由表1可知,KH998、KH1561和KH110與對應的原始親本相比,均對螟蟲具有抗性,且抗性與CK3相當;在農藝性狀方面較其對應的原始親本有所改善,如KH998、KH1561和KH110的千粒重均低于CK3,KH1561的千粒重比其原始親本桂恢1561變小;而KH998和KH110的千粒重較其相應原始親本的千粒重分別增加1.40和3.40 g,產量優勢變大。

表1 T1C-19衍生品系的農藝性狀和抗螟蟲表現
2018年早季和2020年晚季種植抗螟蟲轉基因雜交組合、親本、CK1和CK2,收獲后考查其農藝性狀表現,結果(表2)表明,天豐A/KH110表現抗蟲,其產量比CK2增加1.29%;野香A/KH998和天豐A/KH998表現抗蟲,但產量比CK1分別減產8.69%和0.49%;華浙2A/KH1561、天豐A/KH1561和華浙2A/KH110表現抗蟲,但產量比CK2分別減產1.89%、3.24%和5.56%。
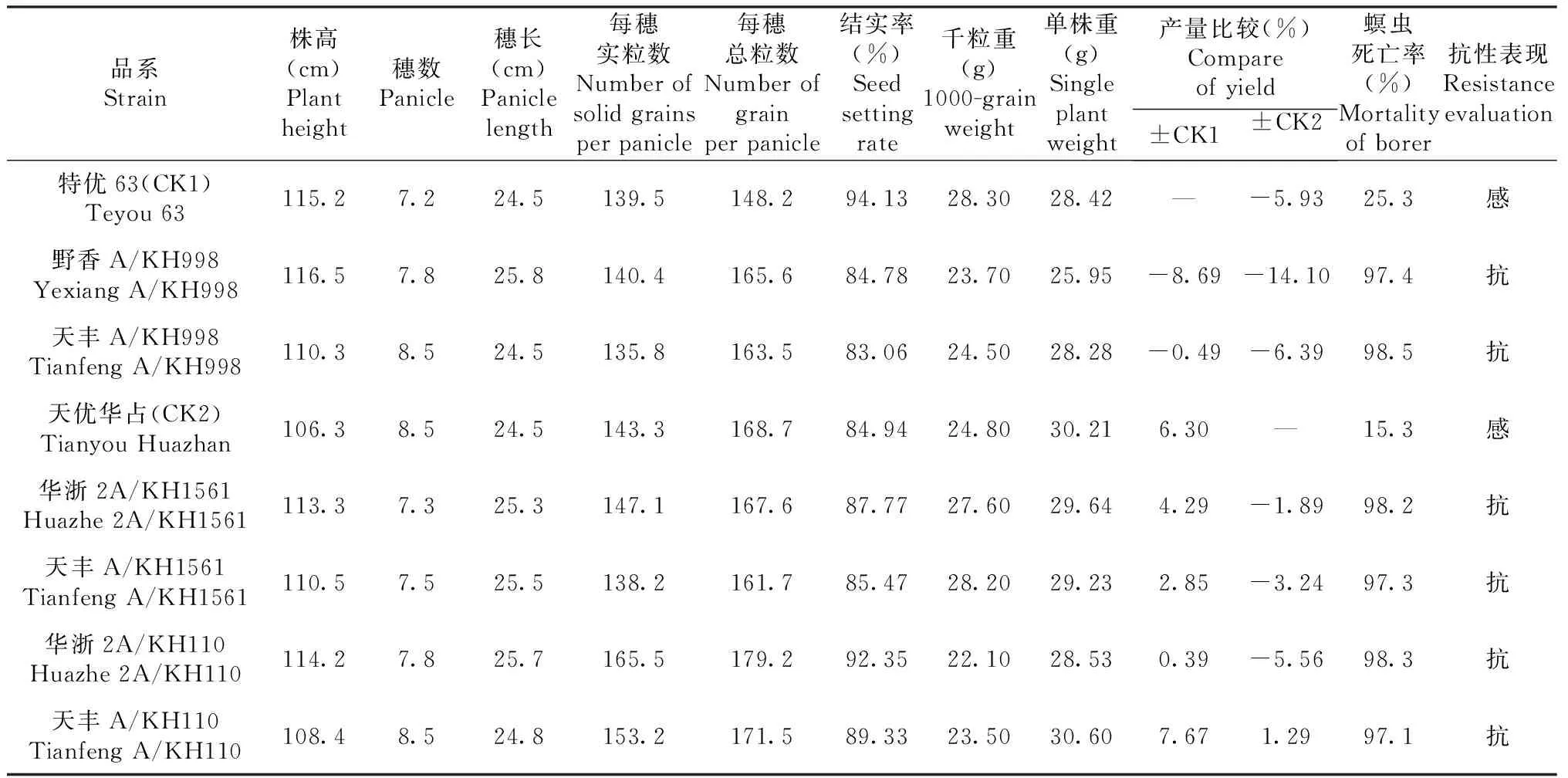
表2 T1C-19衍生品系雜交組合的農藝性狀和抗螟蟲表現
2.3 抗螟蟲轉基因恢復系分子標記檢測結果
水稻移栽返青后,在KH998、KH1561和KH110及其原始親本廣恢998、桂恢1561和桂恢110中分別隨機選取10株的葉片抽提基因組DNA作模板,分別用于與cry1C*基因和bar基因對應的特異引物進行PCR擴增,cry1C*基因在KH998、KH1561和KH110中均能擴增出長度為602 bp的片段,而在陰性對照廣恢998、桂恢1561和桂恢110中無條帶顯示(圖2);bar基因在KH998、KH1561和KH110中均擴增出長度為561 bp的片段,而在陰性對照中無條帶顯示(圖3)。說明抗蟲基因cry1C*和耐除草劑基因bar均已轉入受體親本中,并能在受體親本中穩定遺傳。
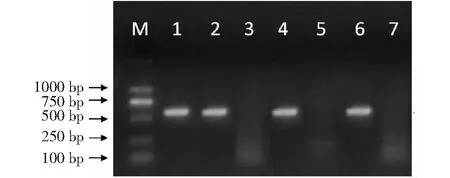
M:DNA Marker;1:T1C-19(陽性對照);2~7分別為KH998、廣恢998 (陰性對照)、KH1561、桂恢1561(陰性對照)、KH110和桂恢110 (陰性對照)的單株;目標產物片段為602 bpM:DNA Marker;1:T1C-19(positive control);2-7 were the individual plants of KH998,Guanghui 998 (negative control),KH1561,Guihui 1561(negative control),KH110 and Guihui 110(negative control);the target product size fragment was 602 bp圖2 KH998、KH1561和KH110的cry1C*基因PCR檢測結果Fig.2 PCR detection results of cry1C*gene in KH998,KH1561 and KH110
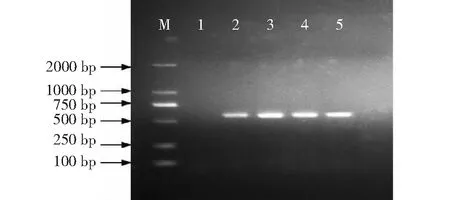
M:DNA Marker;1:陰性對照;2:T1C-19(陽性對照);3~5分別為KH998、KH1561和KH110 的單株;目標產物片段為561 bpM:DNA Marker;1:Negative control;2:T1C-19(positive control);3-5 were the individual plants of KH998,KH1561 and KH110;The target product size fragment was 561 bp圖3 KH998、KH1561和KH110的bar基因PCR檢測結果Fig.3 PCR detection results of bar gene in KH998,KH1561 and KH110
2.4 抗螟蟲恢復系的殺蟲蛋白含量檢測與分析
在KH998、KH1561和KH110的單株中各隨機選擇30個單株,測定其Bt蛋白的表達量(Bt蛋白含量),結果(圖4)顯示,在3個群體中,Bt蛋白含量最大值為1.70 μg/g FW,最小值為0.02 μg/g FW,平均為0.77 μg/g FW,單株間Bt蛋白含量存在較大差異;KH998的Bt蛋白含量較總體水平偏低,平均為0.24 μg/g FW,且個體間差異不明顯,說明該抗性群體的Bt蛋白能穩定表達;KH1561的Bt蛋白含量平均為0.65 μg/g FW,且個體間差異不明顯,說明該抗性群體的Bt蛋白也能穩定表達;KH110的Bt蛋白含量較總體水平偏高,平均為1.41 μg/g FW,個體間差異較明顯(0.02~1.70 μg/g FW),說明該抗性群體的Bt蛋白表達不夠穩定。

KH998:1~30;KH1561:31~60;KH110:61~90圖4 抗性植株中cry1C*殺蟲蛋白含量檢測結果Fig.4 Detection of cry1C*insecticidal protein content in resistant plants
3 討 論
前人研究認為,培育抗螟蟲水稻新品種才是最經濟有效的水稻螟蟲防控措施[5],但我國水稻種質資源中抗螟蟲材料較少,采用常規育種方法很難培育出抗螟蟲水稻品種,而以轉Bt基因水稻材料為供體,通過MAS技術結合農藝性狀篩選,是選育抗螟蟲水稻品種的有效途徑[17,20-21]。
已有研究表明,在通過遺傳轉化獲得轉基因抗蟲水稻的基礎上,借助MAS和回交育種手段可改良轉Bt基因抗蟲水稻中的一些不良性狀,選育出既具有高抗螟蟲又高產優質特性的水稻新品種或品系[25-27],如高方遠等[17]將優質抗稻瘟病水稻恢復系成恢177與轉基因水稻Bt明恢63雜交并回交1次,采用PCR分析、試紙條檢測和田間抗性鑒定相結合的方法,育成兼具轉基因抗蟲性和稻瘟病抗性的水稻新恢復系Bt5198;劉艷等[20]將轉基因水稻Bt明恢63與水稻三系恢復系先恢207雜交并回交1次,采用PCR分析和田間抗性鑒定相結合的方法,育成抗螟蟲水稻新恢復系;田雨等[21]以轉基因水稻Bt明恢63為供體,以水稻品種9311為受體,利用MAS技術對9311的螟蟲抗性進行改良,進一步篩選出抗螟蟲雜交稻組合。這些利用MAS技術進行的水稻抗螟蟲育種,既減少了煩瑣的遺傳轉化工作,又能為轉基因抗蟲育種提供更多優良的親本材料。本研究以T1C-19為抗蟲基因供體,通過回交轉育結合MAS技術育成的抗螟蟲恢復系KH998、KH1561和KH110,均能穩定遺傳cry1C*基因,能成功表達Bt蛋白,且具有與供體親本T1C-19相當的抗蟲能力,能有效抵御螟蟲危害,且農藝性狀與原始親本廣恢998、桂恢1561和桂恢110相近,所配制的雜交組合野香A/KH998、天豐A/KH998、華浙2A/KH1561、華浙2A/KH110、天豐A/KH1561和天豐A/KH110也具有與供體親本T1C-19相當的抗蟲能力,但產量與對照相當或減產,分析其產量優勢不強的原因,一方面選用的都是小粒型、米質優的不育系,所配制雜交組合的千粒重明顯較對照偏低,因此產量優勢不明顯,另一方面也說明雜交組合的特殊配合力不高,下一步需加強雜交組合優勢選育方面的研究,在兼顧米質的同時提高產量。
利用MAS和回交育種技術選育的抗螟蟲水稻新品系,其抗螟蟲性狀均得到顯著提高,表現抗蟲[17-20],但Bt蛋白在不同材料中的表達量不一致,有的能穩定表達,有的表達量差異較明顯。本研究育成的3個抗螟蟲恢復系中,KH998和KH1561的Bt蛋白含量在個體間差異不明顯,說明Bt蛋白能穩定表達;而KH110的Bt蛋白含量個體間差異較明顯(0.02~1.70 μg/g FW),說明該抗性群體的Bt蛋白表達不夠穩定,與田雨等[21]在育成具有9311背景的抗蟲水稻品種時獲得同一時期單株間Bt蛋白含量存在較大差異的研究結果相似。可見,通過轉育獲得的抗螟蟲水稻新品系均表現抗蟲,但Bt蛋白的表達量可能不一致。因此,在轉育抗螟蟲水稻新品系時,在獲得穩定抗蟲性新材料后,建議對其進行Bt蛋白含量的單株測定,以獲取Bt蛋白能穩定表達的株系。
4 結 論
通過回交轉育結合MAS育成的3個抗螟蟲轉基因恢復系KH998、KH1561和KH110均可穩定遺傳cry1C*基因,成功表達Bt蛋白,且具有與供體親本T1C-19相當的抗蟲能力。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
