玉米泛素連接酶U-box基因家族的全基因組鑒定及表達分析
2022-04-25 02:41:54陳曙,張彧,陳卓,金輝
西南農業學報 2022年3期
關鍵詞:分析
陳 曙,張 彧,陳 卓,金 輝
(1. 中國熱帶農業科學院南亞熱帶作物研究所,廣東 湛江 524091;2. 農業部熱帶果樹生物學重點實驗室,廣東 湛江 524091)
【研究意義】泛素26S-蛋白酶體途徑(UPP)作為生物體內蛋白翻譯后的重要修飾途徑之一,由泛素活化酶E1、泛素結合酶E2、泛素連接酶E3及26S蛋白酶體組成,共同調控生物體中蛋白的修飾和降解過程,是真核生物生命活動中最重要的調控機制之一。在UPP中,E3泛素連接酶負責對靶蛋白特異性識別,并進行泛素化修飾[1]。U-box基因家族作為泛素連接酶E3大家族中一類,其U-box結構域決定了U-box E3基因的活性。研究表明,該部位發生突變會導致E3連接酶活性變弱或者喪失[2],致使其在維持細胞分裂、信號轉導、逆境脅迫、生長發育等多個方面的生物學功能紊亂或者徹底喪失[3-6]。因此鑒定玉米泛素U-box基因家族成員,研究基因功能對深入了解玉米生長發育和逆境脅迫等作用機制具有重要意義。【前人研究進展】U-box泛素連接酶最早在酵母UFD2蛋白中發現,也是最早發現的E3泛素連接酶[7]。U-box結構域由大約70個氨基酸組成,在真核生物中高度保守,在不同的植物基因組中數量差異大,這表明U-box在植物的生長發育過程中具有特異性調控[8]。U-box蛋白與RING蛋白結構具有高度的相似性,兩者都具有β-β-a-β折疊結構,氨基酸序列類似,且促進E2結合酶與泛素分子相結合的部位相同。不同之處在于U-box的結構域是通過靜電之間的相互作用形成,通過疏水核心基團代替結構金屬離子,較RING finger結構域而言更加穩定和保守[9-10]。有別于RING finger結構域中通過金屬離子的作用螯合殘基,擬南芥PUB蛋白(Arabidopsis thaliana plant U-box, 簡稱At PUB)AtPUB14是擬南芥中最早鑒定出來含有U-box結構域的蛋白,含有β-β-a-β折疊結構以及鋅指結構,其2個疏水結構基團上的殘基構成特殊的E3結合位點,并通過氫鍵之間的相互作用使U-box結構穩定[11]。目前為止,在擬南芥中已發現64個含有U-box結構域的蛋白[12],在水稻中,鑒定出了77個[13],蒺藜苜蓿中有41個[14],番茄中鑒定出了56個[15]。PUB蛋白參與植物生長發育過程以及對環境脅迫的防御作用。研究表明,擬南芥中AtPUB17基因參與了植物細胞凋亡過程,對植物生長發育有著正調控作用[16]。在煙草和油菜中分別存在其同源基因ACRE276和ARC1,其中ACRE276與AtPUB17的功能具有同源性,而ARC1與AtPUB17功能則并不相同,ARC1含有ARM結構,具有E3連接酶活性,對油菜的自交不親和狀態進行負調控作用[17-18]。在干旱脅迫條件下,水稻OsPUB41基因通過介導OsUBC25后被激活,并與OsCLC6相互作用,并發揮負調控作用[19]。馬鈴薯的PHOR1基因在反義沉默后,植株長勢變緩且變得矮小,對外源GA表現不敏感,響應能力受損;而將PHOR1基因過表達后發現,植株變得對外源GA敏感,對GA合成抑制劑的抗性增強[20]。U-box蛋白在植物對抗環境脅迫中也發揮著重要作用。CHIP蛋白作為U-box家族中一類具有輔助分子伴侶和E3活性的蛋白,在植物遭遇高溫、強光燈不利因素時能提高植物的抗逆性[21]。植物類病變(lesion mimics)突變體是一類在沒有病原物入侵時就能自發產生壞死斑的突變體,這類突變往往使植株增強抗病。水稻SPL11突變體植株在無病原體侵染的情況下發生系統性的自主死亡過程,葉片自發出現病斑,并且植株對一些真菌和細菌性病原體的抗性增強,如稻瘟病菌和白葉枯病菌[22]。玉米是全球主要的經濟糧食作物之一,在玉米的生長發育過程中泛素化過程具有重要的意義,其過程順利進行與否直接影響玉米植株的生長,同時也影響產量和品質[23]。【本研究切入點】目前已完成玉米全基因組的測序工作,為在全基因組范圍內對玉米各基因家族進行基因結構分析和功能預測提供了數據支持。但迄今為止,尚未見有對玉米U-box基因家族的研究和報導。【擬解決的關鍵問題】本研究利用生物信息學方法,通過對玉米全基因組進行分析,鑒定出U-box基因家族中的基因類型、數目以及對基因的結構、理化性質、染色體定位、啟動子功能和基因的表達模式進行分析等,以期為后續深入開展U-box基因功能的相關研究提供理論依據。
1 材料與方法
1.1 數據來源
玉米(ZeamaysL.)全基因組數據來源于Maize Genetics And Genomics Database數據庫(https://www.maizegdb.org/)及Ensembl Plants基因組數據庫(http://plants.ensembl.org/index.html)。擬南芥U-box基因家族基因和蛋白序列下載于擬南芥TAIR數據庫(http://www.arabidopsis),水稻U-box基因家族基因序列下載于水稻全基因組數據庫(http://rice.plantbiology.msu.edu/)。
1.2 數據處理
1.2.1 玉米U-box基因序列的獲取和家族成員鑒定 以目前擬南芥中鑒定出的64個U-box基因家族成員的序列對Ensembl Plants基因組數據庫(http://plants.ensembl.org/index.html)進行基因搜索和序列比對,查找出玉米基因組中所有U-box基因的CDS和蛋白序列。所得結果利用SMART(http://smart.emblheidelberg.de/)在線分析軟件進一步進行結構分析,剔除無典型U-box結構域的序列,最終得到玉米基因組中所有U-box基因。
1.2.2 玉米U-box家族基因的結構分析 根據已獲得的玉米U-box基因的基因序列和CDS序列,利用在線工具GSDS (http://gsds.cbi.pku.edu.cn)分析基因結構,明確內含子和外顯子的組成數量。并通過Mapinspect工具標注每個U-box基因在染色體上的位置,了解U-box基因家族中所有U-box基因在基因組中的分布情況。
1.2.3 玉米U-box基因氨基酸序列屬性分析 利用ExPASy Proteomics Server(http://www.expasy.org/ proteomics )對所有玉米U-box基因進行蛋白分子量、等電點、不穩定系數等理化性質分析預測。利用Plant-mPLocServer(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/#)在線工具分析U-box蛋白的亞細胞定位。二級結構分析利用SOPMA在線程序(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page =npsa_sopma.html)。
1.2.4 玉米U-box基因序列比對及系統進化樹的構建 利用ClustalW工具對水稻、擬南芥、玉米基因組中所有U-box蛋白序列進行比對,結合序列比對結果,利用MEGA 6軟件采用鄰接法(neighbor joining,NJ)構建系統發育進化樹。參數設置:替換模型設置為“Poisson model”,缺口設置為“Pairwise deletion”,校驗參數Bootstrap method 取值為1000。
1.2.5 玉米U-box基因啟動子功能分析 獲取玉米U-box家族基因上游約1500 bp序列,將序列導入在線工具Plant CARE(http:/ /bioinformatics.Psb.Ugent.be /webtools /plantcare /html/)中,歸類基因啟動子上的作用元件,預測分析各基因啟動子所含基序種類和功能。
1.2.6 玉米U-box家族基因在不同組織中的表達模式分析 通過Ensembl Plants (http:/ /plantsa.ensembl.org /index.html) 網站,根據基因登錄號,搜索下載玉米76個U-box基因在胚、胚囊、胚乳、胚珠、子房、根、葉片、穗、花粉等其它共14個不同組織部位的表達情況。利用百邁客在線軟件( http:/ /console.Biocloud.net /static /index.html)繪制基因表達熱圖。
2 結果與分析
2.1 玉米U-box家族基因的獲取和理化性質分析
通過BLAST比對以及搜索,剔除無典型U-box結構域的序列,最終從玉米全基因組中共鑒定出76個含有U-box結構域的基因,如圖1所示。ZmPUB基因在玉米基因組1~10號染色體上均有覆蓋,每條染色體ZmPUB基因數量從2~12個不等,其中1號染色體上最多,為12個,8號染色體上僅含2個。部分U-box基因較為特殊,在染色體上緊密排列形成了基因簇。另外有兩個基因在染色體上無法定位,分別為ZmPUB1和ZmPUB2。表1結果顯示,玉米U-box基因中分子量最大的蛋白ZmPUB22含有氨基酸殘基數目為1353,分子量大小為144.72 kD,最小的U-box蛋白為ZmPUB57,含有94個氨基酸殘基,分子量僅10.76 kD。對等電點進行分析發現,最小的U-box蛋白為ZmPUB34,等電點為4.86,顯酸性。不穩定指數分析顯示,78個ZmPUB基因中,不穩定指數最小的為ZmPUB14,最大的為ZmPUB59,其中不穩定指數小于40蛋白的有13個,蛋白穩定性較好,其余65個ZmPUB蛋白均為不穩定蛋白。
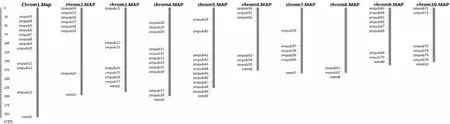
圖1 玉米U-box家族基因在染色體上的位置分布Fig.1 The distribution of maize U-box family genes on chromosomes
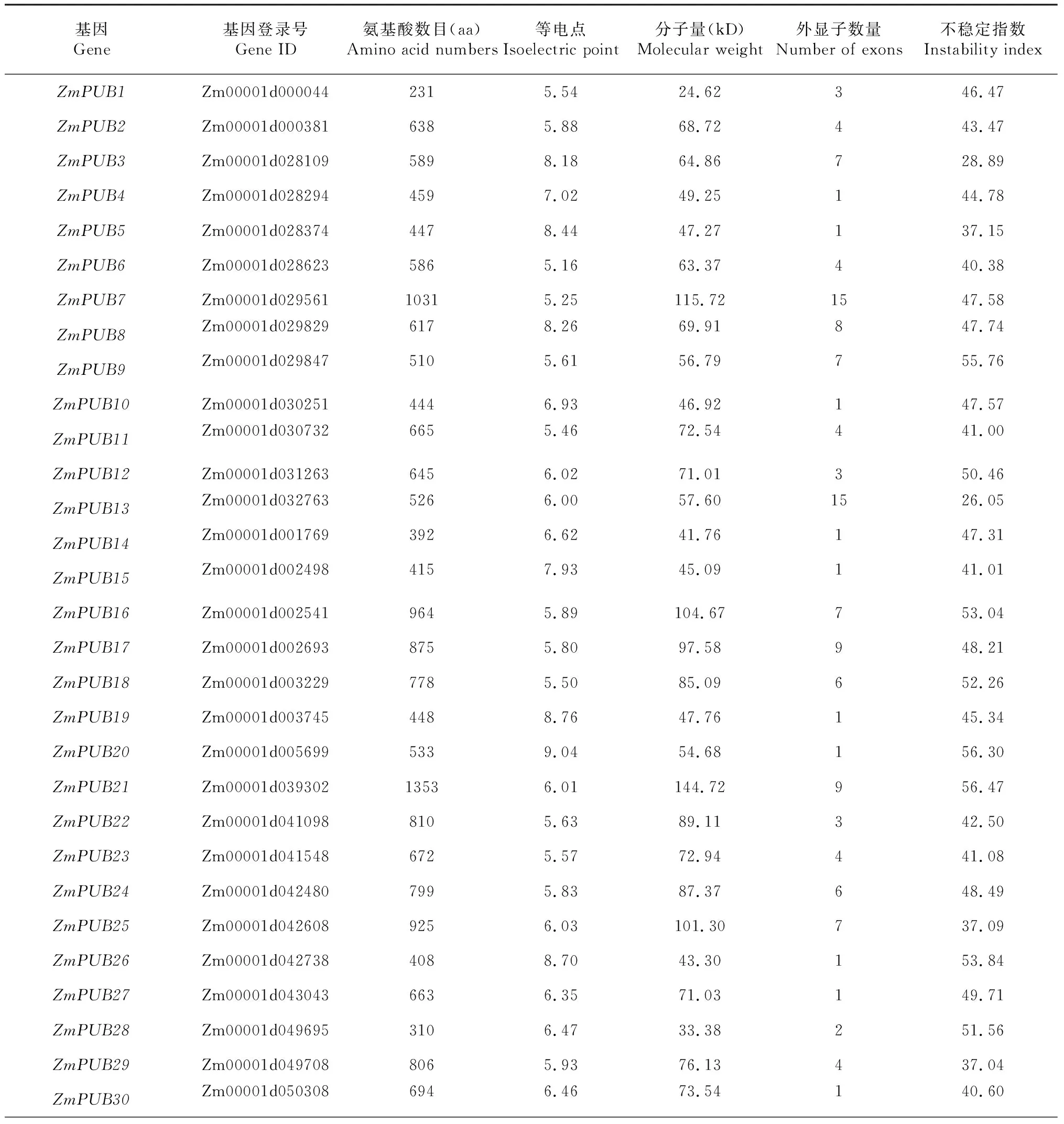
表1 玉米U-box家族基因理化性質分析
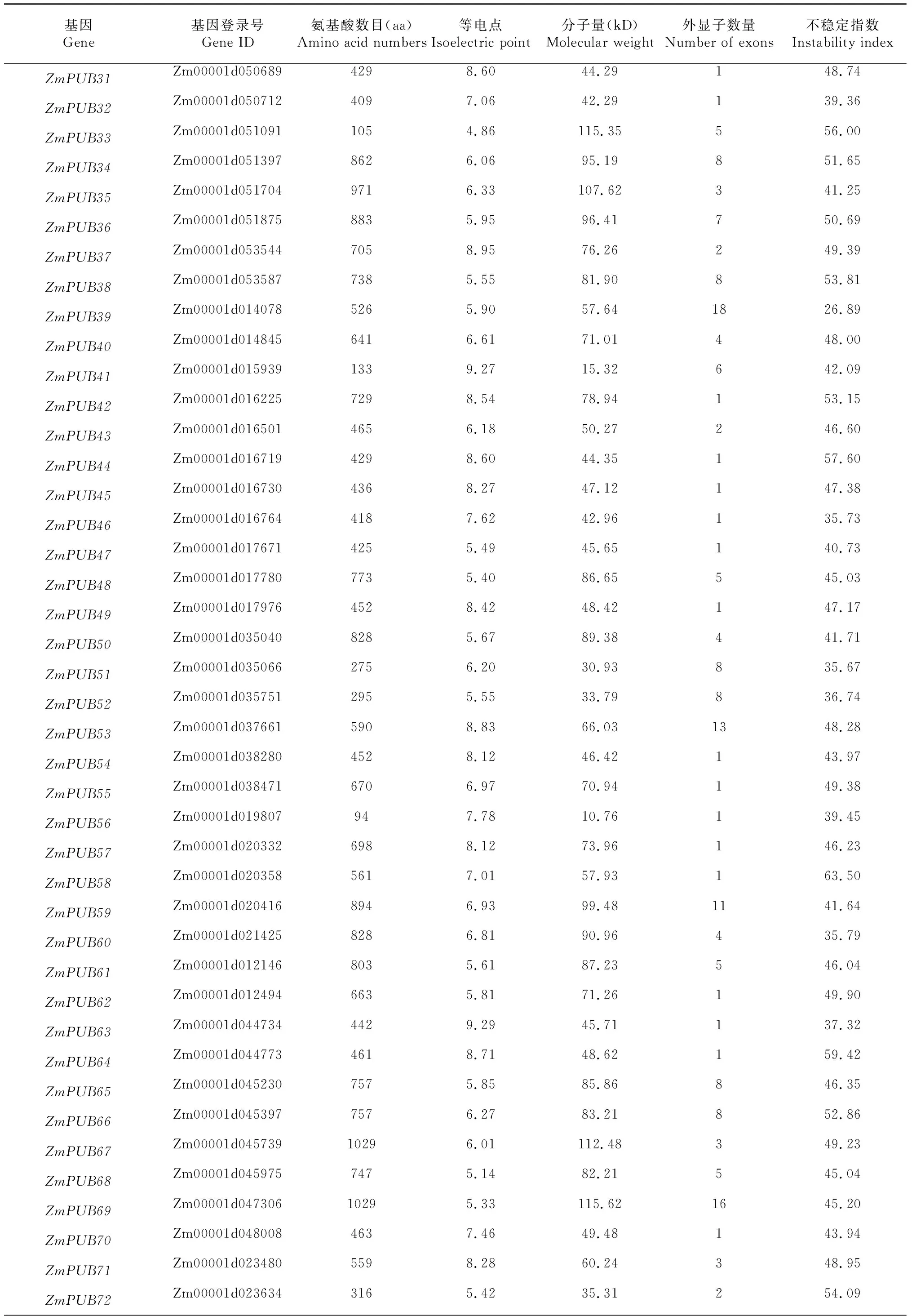
續表1 Continued table 1
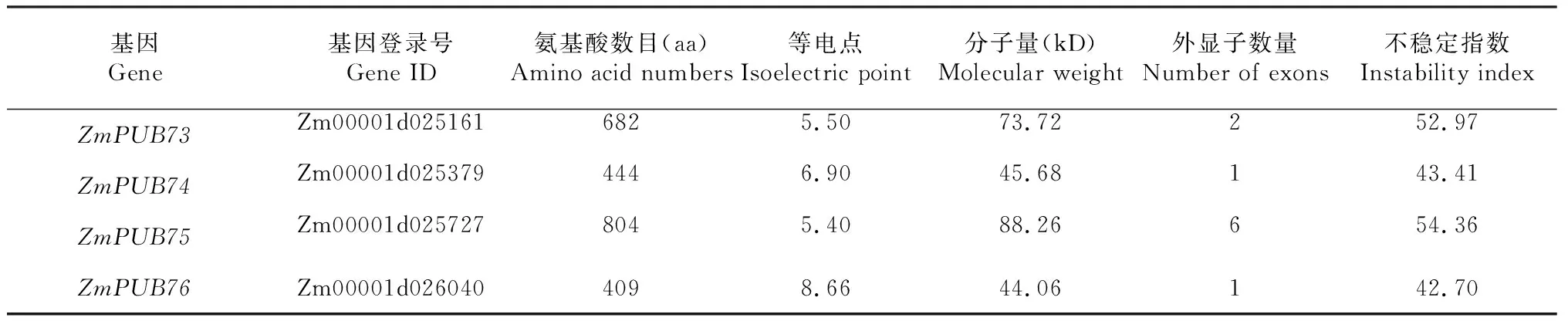
續表1 Continued table 1
2.2 玉米U-box基因家族系統進化及基因結構分析
利用上述篩選出的76個ZmPUB蛋白序列,與62個擬南芥PUB蛋白以及77個水稻PUB蛋白組成數據矩陣,利用MEGA 6進行系統進化分析(圖2),結果顯示玉米、水稻、擬南芥三大U-box基因家族可分為9個亞家族,即PUBⅠ~PUBⅨ。3個作物中各亞家族U-box基因數量差異明顯,亞家族Ⅶ中水稻成員數目最多,為32個,占水稻U-box基因總數的41.6%,在亞家族Ⅲ、Ⅳ、Ⅴ和Ⅵ中則數量較少,分別為0、0、2和1。擬南芥在亞家族Ⅶ中數目為17個,占水稻U-box基因總數的27.4%,數量最多,在亞家族Ⅳ中數目為零,數量最少。玉米在各亞家族中的成員數量分布為2~23個,其中在亞家族Ⅱ中為23個,占玉米U-box基因總數的30.3%,在亞家族Ⅳ和Ⅴ中數目最少,均為2個。

圖2 玉米、擬南芥以及水稻U-box家族基因系統進化分析Fig.2 Phylogenetic analysis of U-box family genes in maize, arabidopsis and rice
亞家族Ⅳ中僅含有2個玉米U-box基因,分別為ZmPUB41和ZmPUB53,不含有水稻和擬南芥成員。表明玉米在歷史進化過程中可能發生了多次家族基因擴增現象,且ZmPUB41和ZmPUB53在經過片段復制后發生了快速變異。利用在線軟件GSDS對玉米U-box基因進行結構分析,并繪制基因結構示意(圖3),結果顯示玉米U-box家族76個成員中共有28個基因無內含子,僅含有1個外顯子,占玉米U-box基因總數的36.8%,表明這些基因進化可能源于轉座子機制。內含子最多的為ZmPUB39,含有17個,其余基因分別含有內含子數目為1~17個不等。
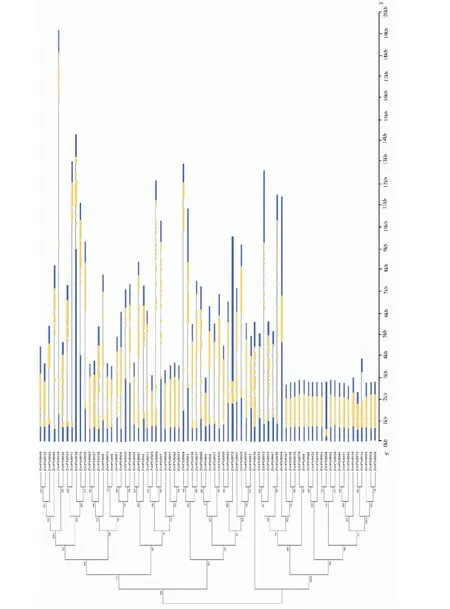
圖3 玉米U-box家族基因結構分析Fig.3 The genetic structure analysis of maize U-box family
2.3 玉米U-box基因啟動子功能預測和分析
考慮到玉米U-box基因家族較大,成員數較多,筆者根據系統進化分析結果,從亞家族PUB1~PUB9中每個亞家族選取1~2個成員,總計14個基因作為分析對象,具體基因編號見表2。表2統計結果顯示,受檢基因所含功能元件主要包括植物激素響應元件、逆境脅迫應答元件、光反應元件和周期性調控相關元件4類。所有基因中所含光反應元件最多,說明玉米U-box基因參與了植株光合作用或相關代謝活動。在植物激素調控方面,主要分為ABA、MeJA、IAA、GA、乙烯、水楊酸6類,各受檢基因所含元件數量差異明顯。14個受檢基因中,部分基因不含乙烯調控元件,其余基因所含數量為1~4個,含量較少,說明各基因在參與乙烯調控中可能不作為主效基因發揮作用。其余基因如ZmPUB35含有ABA調控元件最多,ZmPUB1含有MeJA調控元件最多,ZmPUB41含有IAA和水楊酸調控元件最多,ZmPUB33含有GA調控元件最多。同理在逆境脅迫調控中,對應干旱脅迫、厭氧應答、低溫調控中所含元件數量最多的基因分別是ZmPUB2、ZmPUB1和ZmPUB73,在周期性調控相關元件中ZmPUB75所含元件最多,這些現象說明不同亞家族成員間各基因主效功能不一樣,表明在玉米進化過程中,U-box家族基因在復制、快速擴增的同時也發生了大量的變異,使玉米U-box家族基因功能更加多樣化。

表2 玉米U-box基因啟動子功能元件分析
2.4 玉米U-box基因在不同組織的表達分析
利用搜索出的64個玉米U-box基因表達數據(12個基因未能搜索出相關數據),制作基因表達熱圖(圖4)。基因表達聚類分析結果顯示,玉米U-box基因在不同組織中表達差異較大,各U-box基因表達差異明顯。ZmPUB3、ZmPUB6、ZmPUB7、ZmPUB11、ZmPUB22、ZmPUB29、ZmPUB50、ZmPUB52在種子的發育以及組成器官如胚、胚囊、胚乳、珠心、糊粉層、子房、胚珠中大量表達,表明這些基因可能參與了種子的形成以及萌發等生理活動過程。部分基因如ZmPUB19、ZmPUB21、ZmPUB26、ZmPUB49等在被檢測組織中均未見表達,說明未參與上述器官相關生命活動。ZmPUB12、ZmPUB18、ZmPUB30、ZmPUB40、ZmPUB75在花粉中表達量較高,而在其它被檢測組織中均未見表達,說明這5個基因功能特異性較強,可能在玉米花粉受精過程中發揮了重要作用。
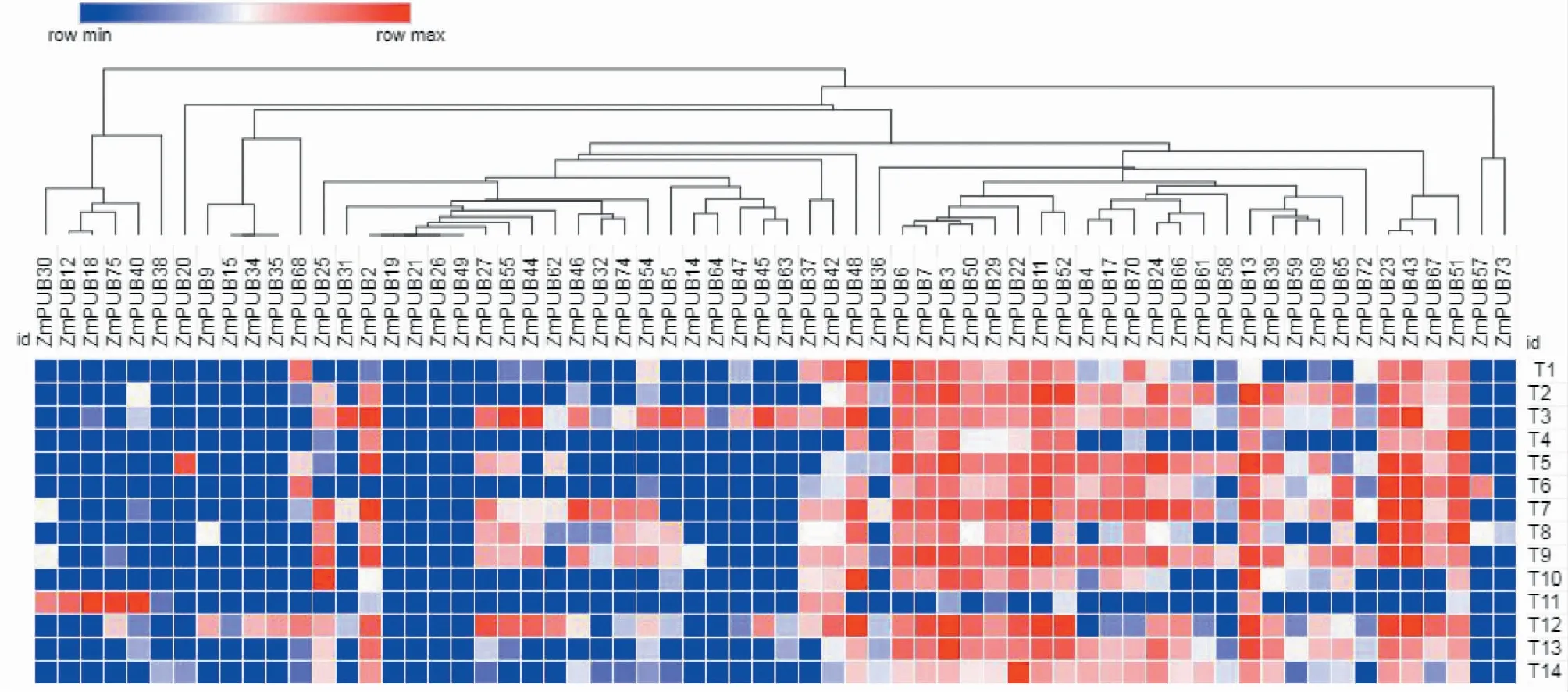
T1:維管束鞘;T2:胚;T3:胚囊;T4:胚乳;T5:花絲;T6:葉片;T7:珠心;T8:糊粉層;T9:子房;T10:胚珠;T11:花粉;T12:根;T13:雄穗;T14:營養分生組織T1:Vascular sheath;T2:Embryo;T3:Embryo sac;T4:Endosperm;T5:Filigree;T6:Leaf;T7:Nucellus;T8:Aleurone layer;T9:Ovary;T10:Ovule;T11:Pollen;T12:Root;T13: Tassel;T14:Vegetative meristem圖4 玉米U-box家族基因在不同組織中的表達分析Fig.4 Expression analysis of maize U-box family genes in different tissues
3 討 論
U-box基因家族是泛素連接酶E3家族中的主要家族之一,在植物的生長發育及逆境脅迫應答中發揮了重要的作用,包括人類在內,U-box蛋白幾乎存在于所有的真核生物中[2]。U-box蛋白最早在酵母中被發現,隨后相繼在擬南芥、水稻、番茄、柑橘、雷蒙德氏棉、苜蓿等作物中被鑒定報導。隨著玉米全基因組測序工作的全面完成,大量的玉米基因家族功能被研究和驗證,極大的推進了玉米基礎研究領域的發展,生物信息學作為研究植物基因功能、調控模式以及系統進化關系等功能的重要工具被廣泛使用。目前玉米E1、E2家族基因已經相繼被鑒定出來,而E3家族包括多亞基家族cullin-RING (CRLs)和單亞基家族如HECT(homologous to E6-AP COOH-Terminus) 、RING (really interesting new gene) 、U-box 共4類家族,目前尚未見有對U-box基因家族的系統性報導。本研究從玉米全基因組中鑒定出了76個U-box基因成員,與水稻U-box成員數相似,比擬南芥、苜蓿和番茄中U-box成員數目多,表明不同物種的U-box成員數量存在差異。部分玉米U-box基因染色體上排列緊密,形成了數目較多的基因簇,加強了基因功能的表達。玉米不同U-box亞家族成員間所含氨基酸數目、等電點、分子量、外顯子數量以及不穩定指數差異較大,說明其理化性質有較大不同,而同一亞家族間各成員上述各理化指標相似度較高,暗示其理化性質相似。基因內含子/外顯子排列特性對基因功能具有重大影響[24]。基因結構及系統進化分析結果顯示,同一個亞家族中,處在不同分支的U-box基因在外顯子、內含子數目以及蛋白序列長度等方面均有差異,但同屬一個分支的旁系同源基因之間如ZmPUB75與ZmPUB18、ZmPUB29與ZmPUB50、ZmPUB4與ZmPUB70及ZmPUB35與ZmPUB67等理化性質及基因結構高度相似。說明玉米泛素連接酶U-box家族基因在進化過程中具有高度保守性,但同時在漫長進化過程中使得基因功能多樣化,以適應環境變遷和種群延續。
啟動子作為位于基因5′端上游的DNA序列結構,控制著基因轉錄的起始以及表達程度,繼而決定了編碼特異性蛋白的功能。玉米U-box基因在植物的各種生命活動如生長發育、逆境脅迫、信號轉導以及細胞周期調控等方面起著非常重要的作用[3-6]。進行啟動子結構和基序分析對研究基因的功能具有重要的作用。本研究通過對14個玉米U-box基因啟動子功能預測分析發現各基因均含有大量光反應元件,說明U-box基因可能參與了植物光合作用調控途徑。在所有受檢基因中均發現有生長素、脫落酸以及茉莉酸甲酯順式作用元件,暗示U-box基因可能參與了玉米植株的生長發育、果穗的成熟以及病蟲害應激性防御過程。在逆境脅迫方面,干旱響應、厭氧應答和低溫調控三類元件在玉米U-box基因中均存在,說明U-box基因在植株應對這些逆境脅迫中均發揮了相應作用。
進行基因組織表達模式分析是了解基因生物學功能的重要方法之一。本研究中,對玉米14個不同組織部位進行基因表達分析發現,不同U-box基因間組織差異性表達明顯。基因表達熱圖顯示,部分基因表達不具有特異性,如ZmPUB3、ZmPUB6、ZmPUB7、ZmPUB22、ZmPUB29、ZmPUB50除在花粉中表達量較低外,在其它13個受檢組織中均有較高表達量。而部分基因表達特異性強,如ZmPUB12、ZmPUB18、ZmPUB30、ZmPUB40、ZmPUB75僅在花粉中有較高表達量,在其它組織中幾乎不表達。這些現象說明玉米U-box家族基因功能具有互補性,揭示了U-box家族在進化過程中,在基因復制擴增的過程中也逐漸產生了新的變異,使得整個基因家族功能更加全面完善。基因在特殊器官中高水平表達意味著其在相關過程中發揮關鍵作用。研究發現柑橘CcPUB10基因能夠調控柑橘葉、花、莖和果的生長發育[25]。蘋果MdPUB24在葉片中高度表達,且對外源ABA、NaCl和低溫脅迫呈負調控反應[26]。本研究中,部分玉米U-box基因在種子相關器官中大量表達,暗示了其可能參與了種子的形成以及萌發等生理活動過程,但具體作用機制還不清楚,需后續對相關基因的功能進行進一步研究和驗證。
4 結 論
本研究依據玉米全基因組數據庫,從中鑒定出了76個玉米U-box家族成員,經篩選、鑒定后其蛋白均含有U-box保守結構域。各成員含有內含子數目差異較大,部分基因內含子數量為零。啟動子功能預測分析發現,U-box基因可能參與了玉米光合作用、激素應答和逆境脅迫應答反應,且響應程度不同,顯示出各成員間基因功能特異性明顯。基因表達模式分析結果顯示玉米U-box基因表達具有組織特異性。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
