泛素結合酶9 通過核因子κB 通路誘導胃癌細胞SGC7901 凋亡的研究
2022-04-27 02:15:30袁金華曾憲晶劉伍才羅亮
藥品評價 2022年3期
袁金華,曾憲晶,劉伍才,羅亮
井岡山大學附屬醫院,江西 吉安 343000
胃癌是消化道常見的惡性腫瘤,根據全球癌癥研究機構的數據統計,胃癌位于全球惡性腫瘤發病的第五位,死亡率位于第三位。我國是胃癌的高發區,發病率和死亡率分別占到全球的42.6%和45.0%[1],且發病呈年輕化趨勢。當前胃癌治療方法主要是手術、放療和化療,但預后較差,尋找有效的治療手段對本病的治療具有重要意義。
研究表明:核因子-κB(NF-κB)是真核細胞轉錄因子,NF-κB 信號通路可以調控細胞増殖、分化和凋亡等重要生理功能,與腫瘤發生密切相關[2]。泛素結合酶9(Ubc9)是小泛素相關修飾物(SUMO)過程中唯一的結合酶,Ubc9 在細胞增殖、有絲分裂、周期進程、分化等方面發揮著重要作用[3],Ubc9 在多種腫瘤中過表達,可能參與了腫瘤發生和進展的病理過程,并且可能與腫瘤的耐藥相關[4],研究已證實Ubc9 是一個促癌基因[5],抑制Ubc9 基因后NF-κB 信號通路的凋亡蛋白降低[6]。基于此,我們推測Ubc9 基因可能通過影響NF-κB信號通路來抑制胃癌細胞的增殖及凋亡,為治療提供理論參考。
1 材料與方法
1.1 主要實驗材料
胃癌SGC7901 細胞株(購自上海賽默生物有限公司),兔抗Ubc9 單克隆抗體、兔抗p65 單克隆抗體、兔抗IKBα 單克隆抗體、兔抗Bcl-2 單克隆抗體、山羊抗小鼠β-actin(美國Abcam 公司),Ubc9 引物(Generay Biotech 公司),RNA 提取試劑盒(上海碧云天生物技術有限公司),PCR 檢測試劑盒(上海碧云天生物技術有限公司),Opti-DMEM、轉染試 劑Lipofectamine 2000(Invitrogen 公 司),Trizol Reagent(Invitrogen 公司),胎牛血清(上海碧云天生物技術有限公司),RPIM-1640 培養基(Hyclone公司)。
1.2 細胞培養、分組、轉染
SGC7901 細胞復蘇后培養于含10%胎牛血清的RPIM-1640 培養基中,于5%二氧化碳、37 ℃培養箱中傳代培養,每3~4 d 按照1∶3 比例傳代1 次,取對數生長期細胞進行相關實驗。按5×104mL 的細胞密度接種至六孔板中后放入恒溫培養箱中繼續培養。將實驗細胞分為四組:正常胃癌細胞A 組(Normal 組),脂質體B 組(Lipo-2000 組),空載質粒C 組(NC-shRNA 組),及目的質粒D 組(Ubc9-shRNA 組),質粒的構建與測序均由上海吉瑪公司負責完成,細胞轉染方法:準備5 個1.5 mL的EP 管,分別向每管內加入0.5 mL Opti-DMEM,其中2 個EP 管做好標記分別加入10 μg 空載質粒及目的質粒,余3 管加入20 μL 的lipo2000,混合后靜置。接著向加入lipo2000 的3 個EP 管分別加入目的質粒、空載質粒以及500 μL Opti-DMEM,混勻,最后將配制好的各EP 管內液體分別加入所對應的培養皿中,置入培養箱中培養。
1.3 qRT-PCR 檢測基因表達
細胞轉染48 h 后,使用RNA 提取試劑盒提取細胞總RNA,并反轉錄得到cDNA,以此為模板進行qRT-PCR 目的基因擴增,結果計算根據LC 480檢測得出的CT 值為cDNA 熒光強度達到所設定閥值的循環數,ΔCt=目的基因CT值-內參基因CT值,每個樣品中mRNA 相對表達量=2-ΔCT× 100%。
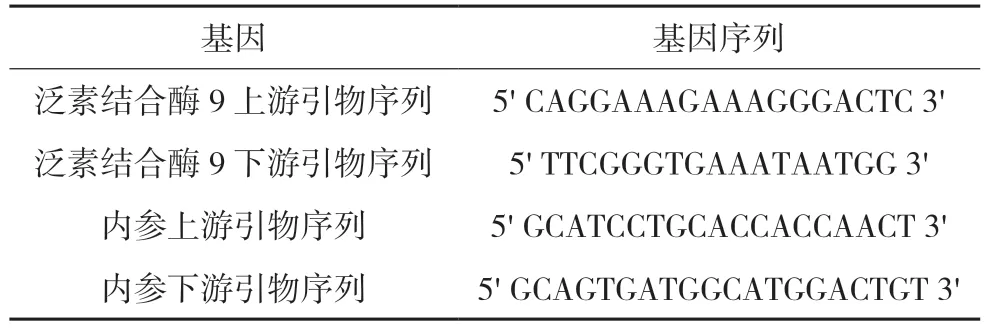
表1 引物序列
1.4 Western blot 檢測蛋白表達
采用蛋白提取試劑盒提取細胞總蛋白,BCA 法測定總蛋白濃度,上樣SDS-PAGE 膠,電泳,轉膜、封閉,抗體孵育,顯影成像,利用凝膠圖像處理系統分析目標條帶灰度值,記錄其相對表達量。
1.5 流式細胞儀檢測細胞周期與凋亡
細胞周期檢測:調整細胞濃度為1×106個/mL,離心,棄上清,重懸后收集細胞,采用1 mL 已配制好的周期試劑檢測各組細胞的周期,按照試劑盒說明書進行反應后,30 min 內進行流式細胞儀檢測細胞的周期。細胞凋亡測定:調整細胞濃度為1×106個/mL,離心,棄上清,重懸后收集細胞,滴 入500 μL Binding Buffer溶液重懸后加入5 μL AnnexinV-FITC 混勻,再加入5 μL 碘化丙啶,混勻,室溫避光孵育,1 h 內應用流式細胞儀檢測各組細胞的凋亡情況。
1.6 統計學方法
2 實驗結果
2.1 轉染效率檢測
24 h 后在熒光顯微鏡下觀察熒光的顯示情況,當質粒成功轉染后,質粒進入胃癌細胞即可在熒光顯微鏡呈現肉眼可見的綠色熒光,見圖1。
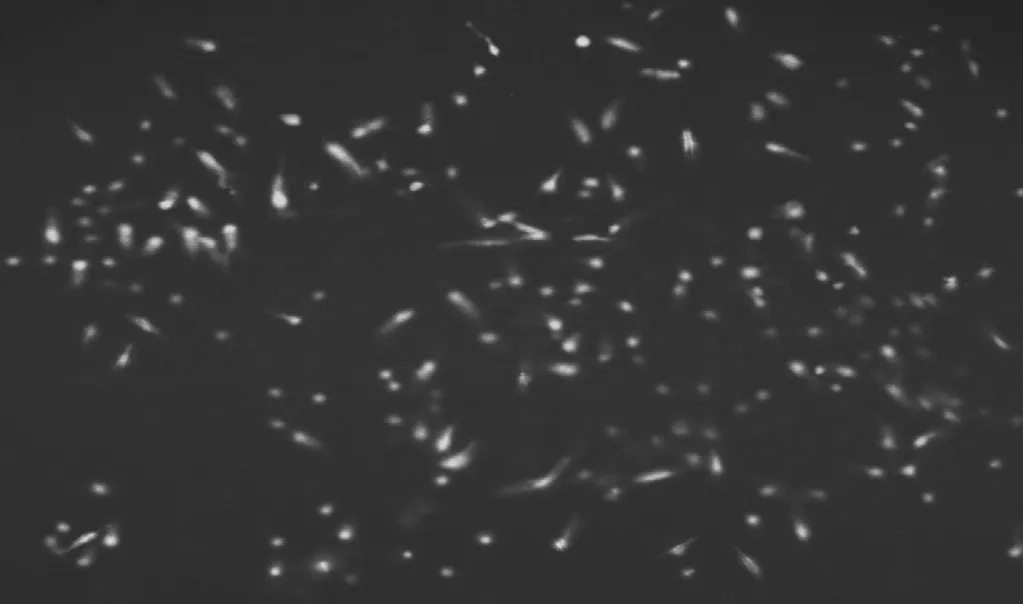
圖1 熒光顯微鏡下觀察mir-Ubc9向SGC 7901細胞轉移情況(×200)
2.2 RT-PCR 檢測轉染后各組Ubc9 基因mRNA 的表達水平
各組細胞經相應的處理24 h 后,將Normal 組,Lipo-2000 組,空載質粒NC-shRNA 組作為對照組,與對照組相比,人工合成的針對Ubc9 基因的miRNA 質粒(Ubc9-shRNA 組)能顯著地抑制胃癌SGC7901 中Ubc9 基因的表達(0.31±0.06),差異有統計學意義(P<0.05)。見圖2。
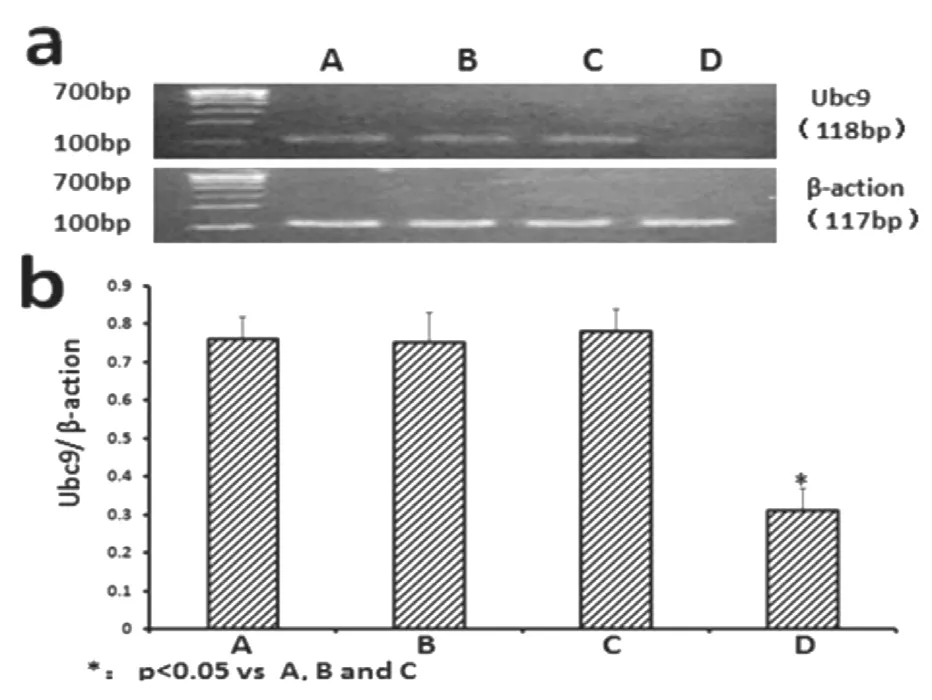
圖2 各組泛素結合酶9 mRNA表達情況:A.正常胃癌細胞組;B.脂質體組;C.空載質粒組;D.目的質粒組
2.3 Western blot 檢測轉染mir Ubc9 后細胞內Ubc9與α-SMA 的表達水平
與對照組相比,人工合成的針對Ubc9 基因的miRNA 質粒明顯抑制了SGC7901 中Ubc9 蛋白的表達,差異有統計學意義(0.28±0.06,P<0.05)。同時SGC7901細胞內α-SMA蛋白表達量也明顯下降,差異有統計學意義(0.28±0.06,P<0.05)。見圖3、表2。
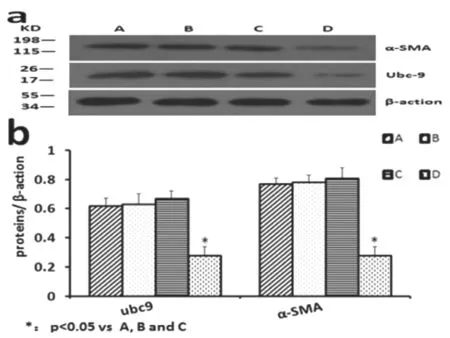
圖3 各組泛素結合酶9和a-平滑肌肌動蛋白表達情況:A.正常胃癌細胞組;B.脂質體組;C.空載質粒組;D.目的質粒組
表2 各組泛素結合酶9和α-平滑肌肌動蛋白蛋白表達水平(,n=4)
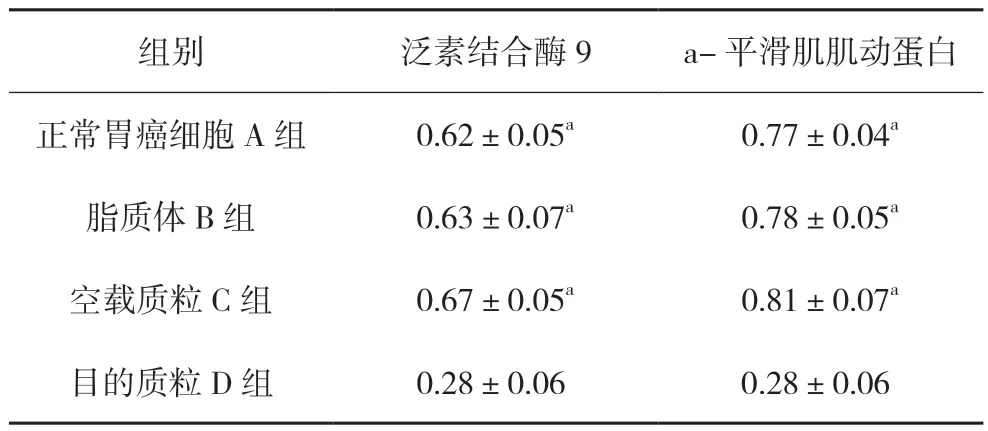
表2 各組泛素結合酶9和α-平滑肌肌動蛋白蛋白表達水平(,n=4)
注:與D組比較,aP<0.05。
2.4 流式檢測結果
2.4.1 細胞周期結果 實驗結果顯示:轉染后胃癌SGC7901 細胞的周期被阻滯在G2/M 期,并且能夠促進SGC7901 的凋亡,與各對照組相比差異有統計學意義(20.45±2.71,P<0.05),提示Ubc9 基因可能調控SGC7901 細胞的細胞周期。見圖4、表3。
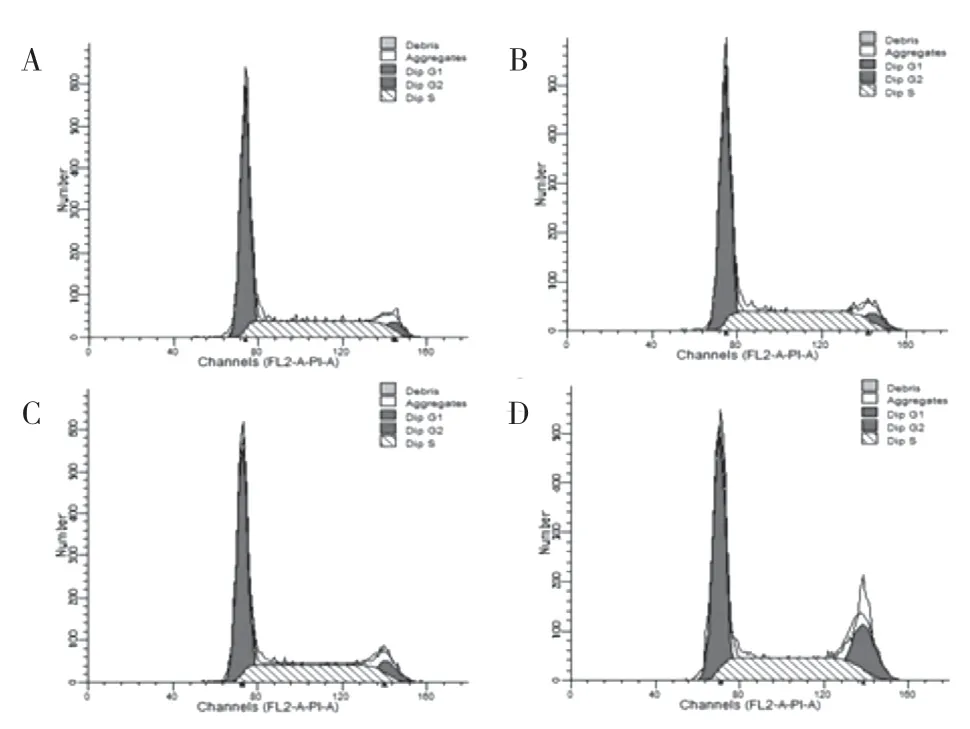
圖4 細胞周期情況:A.正常胃癌細胞組;B.脂質體組;C.空載質粒組;D.目的質粒組
表3 各組SGC 7901細胞周期情況(,n=4)
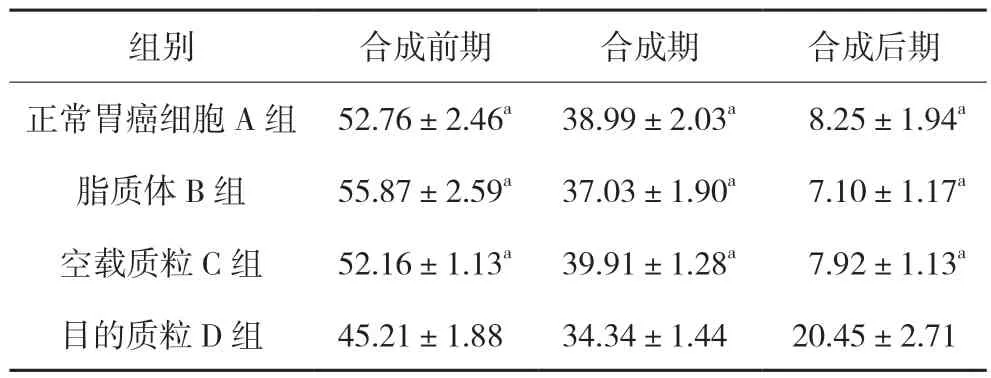
表3 各組SGC 7901細胞周期情況(,n=4)
注:與D組比較,aP<0.05。
2.4.2 細胞凋亡結果 實驗結果顯示:轉染后SGC7901細胞的凋亡增加明顯,與各對照組相比差異有統計學意義(P<0.05),提示Ubc9 基因可能參與調控SGC7901 細胞的凋亡。見圖5、表4。
表4 各組SGC 7901細胞凋亡率(,n=4)
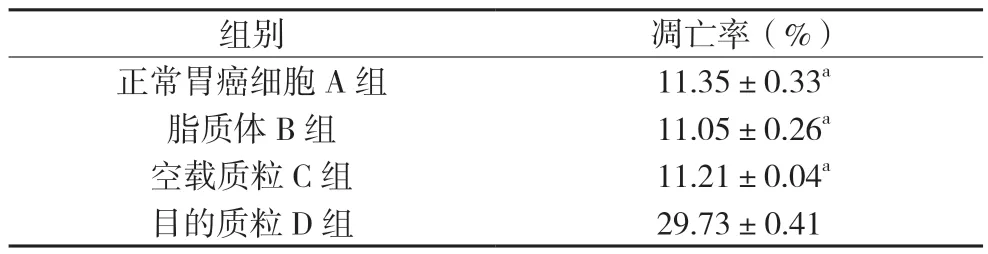
表4 各組SGC 7901細胞凋亡率(,n=4)
注:與D組比較,aP<0.05。
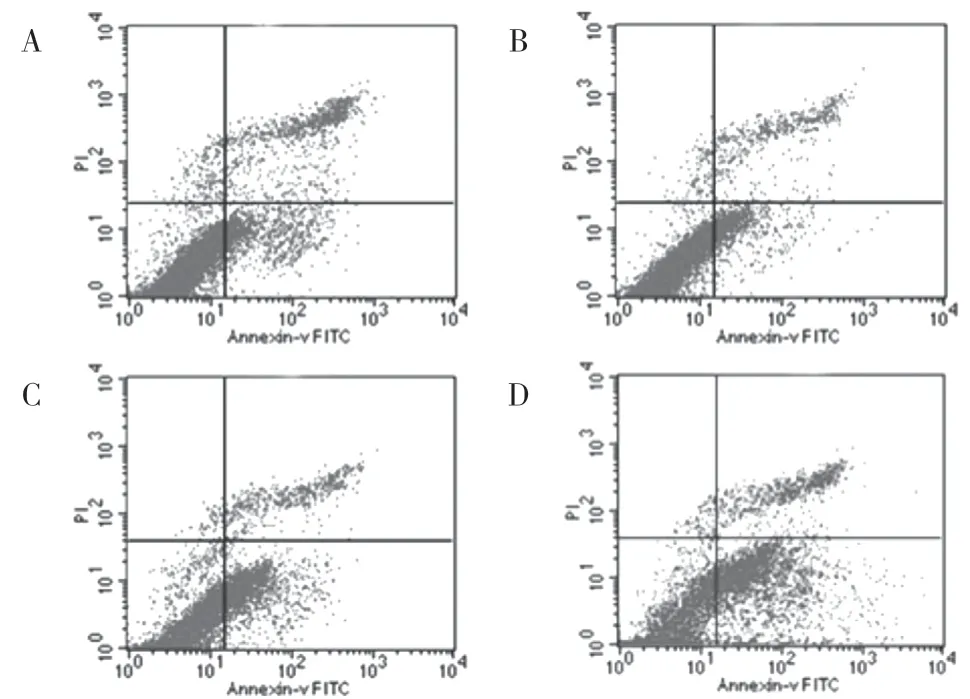
圖5 細胞凋亡情況:A.正常胃癌細胞組;B.脂質體組;C.空載質粒組;D.目的質粒組
2.5 Western blot 檢測細胞IKBα、p65 與Bcl-2 蛋白表達結果
與對照組相比,Ubc9-shRNA 組SGC7901 細胞內p65 與Bcl-2 蛋白表達量明顯下降(0.28±0.06,0.28±0.04,P<0.05),而IKBα 的表達量增加明顯(0.9±0.05,P<0.05),差異有統計學意義(P<0.05)。見圖6 與表5。
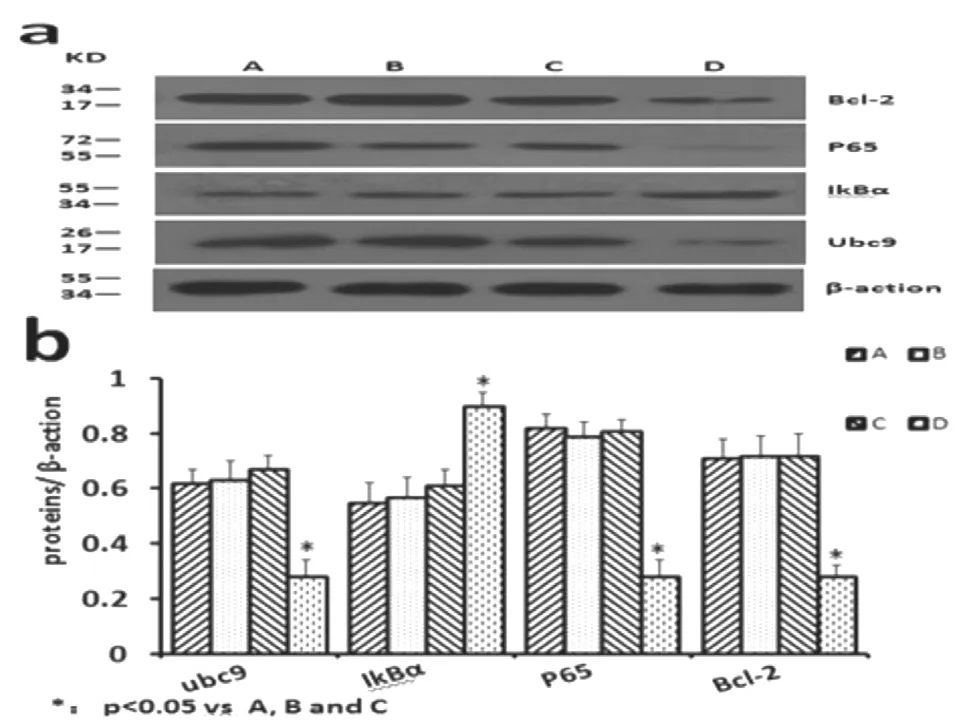
圖6 各組相關蛋白表達情況:A.正常胃癌細胞組;B.脂質體組;C.空載質粒組;D.目的質粒組
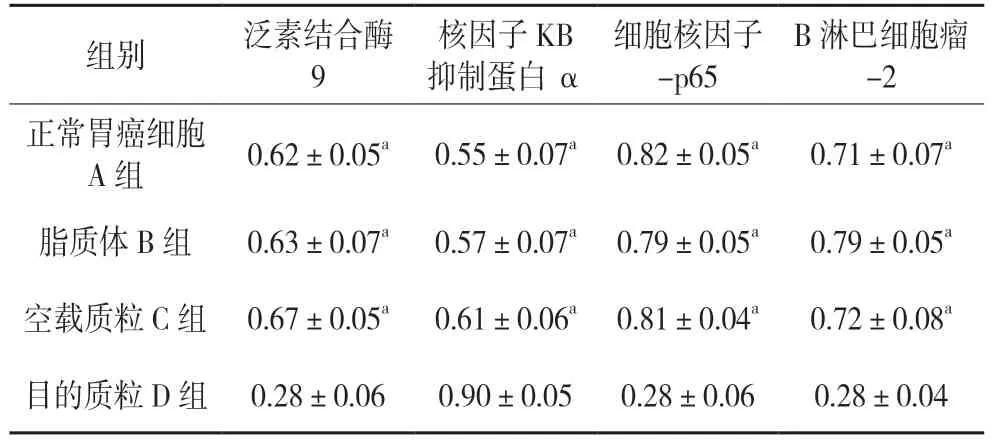
表5 各組相關蛋白表達水平表
3 討論
誘導腫瘤細胞凋亡是抗癌藥物治療腫瘤的重要機制之一[7-8]。研究表明,泛素樣小分子修飾SUMO 參與了多種細胞進程,如細胞周期的調控、細胞增殖、細胞凋亡及DNA 損傷修復等[9]。SUMO是一個重要的、多步驟的蛋白質翻譯后修飾過程,而Ubc9 在SUMO 化這一過程中至關重要,Ubc9 是SUMO 修飾過程中唯一的E2 結合酶[10],能增強SUMO 化修飾從而調控細胞凋亡[11]。
人類胃癌、肝癌、結直腸癌等腫瘤中均發現NF-κB 通路極度活躍,它可通過調節生長、細胞周期、細胞侵襲等相關環節促進癌變[12],NF-κB 是一種重要的核轉錄因子,與炎癥、凋亡等過程密切相關。NF-κB 是普遍存在于細胞質中的一種快反應因子,在靜息狀態下,它與抑制因子IκB 形成復合物,以無活性的潛在狀態存在于細胞質中。NF-κB信號通路是由細胞外的刺激引起的,受體蛋白接受刺激后先活化IκB 激酶(IKK)。IKK 將細胞內NFκB·IκB 復合物的IκB 亞基調節位點的絲氨酸磷酸化,使得IκB 亞基被泛素化修飾,進而被蛋白酶降解,從而釋放NF-κB 二聚體[13]。p50 和p65 組成的二聚體為NF-κB 的主要形式,p50 是與DNA 結合的部位,p65 參與基因轉錄的起始調節,p65 是NF-κB 的主要活性形式,并可促進p50 與DNA 結合,當p65 與細胞質內抑制蛋白IκB 結合,可掩蓋p50 上核定位信號,僅有p65 的蛋白C 末端含有轉錄激活區域,能夠直接啟動基因的轉錄[14-15]。本研究中觀察:Ubc9 能抑制SGC7901 細胞內的p65入核,從而抑制NF-κB 進入細胞核與DNA 特定靶кB 部位結合,進而抑制NF-κB 的活化,同時本研究結果也發現胃癌SGC7901 細胞轉染Ubc9 后,與各對照組比較,Ubc9-shRNA 組IKBα 表達量增加,p65 的表達量減少(P<0.05),提示人工合成的針對Ubc9 基因的miRNA 質粒明顯能夠抑制NF-κB的轉錄激活,證實了文獻[13]研究結論,Ubc9 能與IKBα 結合并發生相互作用,延緩IKBα 的降解和NF-κB 的轉錄激活。
細胞凋亡一種程序性的細胞死亡,Bcl-2 基因主要的生理功能是減少細胞凋亡[16],研究發現Bcl-2 基因在多種惡性腫瘤組織中高表達。研究證實,凋亡是由于caspases 家族蛋白的瀑布式反應而導致的細胞自身溶解的過程,Bcl-2 蛋白不僅能抑制細胞凋亡,亦可能拮抗抑癌基因所介導的細胞凋亡。作為抗凋亡蛋白,Bcl-2 蛋白能夠通過抑制腫瘤凋亡來促進其擴散。轉錄激活后入核的NF-κB 可調控反向凋亡相關基因Bcl-2、Bax 及細胞增殖相關基因Cyclin D1 的轉錄,從而發揮對細胞增殖、分化、凋亡及細胞周期的調控。在本次實驗中,我們觀察發現Ubc9 轉染后胃癌SGC7901細胞的周期被阻滯在G2/M 期,并且細胞的凋亡增加明顯,NF-κB 轉錄激活后Bcl-2 水平降低,與各對照組相比,差異有統計學意義(P<0.05),提示Ubc9 可能是通過NF-κB 通路影響Bcl-2 基因的表達來調控胃癌SGC7901 細胞的凋亡。Mo 等[17]下調Ubc9 后乳腺癌MCF-7 細胞生長受抑制,凋亡增加,Ubc9 是通過Bcl-2 發揮作用,結果表明Ubc9 通過影響Bcl-2 基因表達而具有抗凋亡作用。
綜上所述,人工合成的針對Ubc9 基因的miRNA質粒能夠抑制SGC7901 細胞的增殖并促進其凋亡,為胃癌的治療提供了一種新的實驗依據。然而對于其具體的通路以及如何影響胃癌細胞周期的分子機制尚未明確。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
