低溫預處理對不同儲存時間黃帚橐吾種子發芽的影響
2022-05-11 10:44:44任釗江王玉琴王宏生宋梅玲
草原與草坪 2022年1期
關鍵詞:差異
任釗江,王玉琴,王宏生,宋梅玲
(1.青海大學農牧學院,青海 西寧 810016;2.青海大學畜牧獸醫科學院(青海省畜牧獸醫科學院),青海 西寧 810016)
種子萌發是種子植物生活史中實現種群更新和物種延續的關鍵環節之一[1]。植物種子發芽能力是其價值的重要體現。種子萌發期和幼苗建植期易受外界環境的影響,導致其死亡率較高[2]。種子能否成功發芽并定植是其種群更新的重要因素[3]。植物種子發芽能力受多種環境因素的影響,其中溫度是影響發芽能力的重要因素,由于溫度影響種子內部各種酶的活性,進而影響種子內部物質的轉運以及種子和外界的物質交換,溫度過高或過低都不利于種子萌發[4]。種子萌發及萌發后的生長狀況受溫度條件的影響,其萌發所需的最適溫度往往與其起源地生態條件和所處的生存環境有關[5]。因此,研究溫度對種子萌發的影響對于明確其在特定生長環境中的發芽和生長機制具有重要意義。
盡管種子可在較大的溫度范圍內完成萌發過程,但低溫處理后,種子的休眠狀態和發芽特征等卻有顯著差異[6]。種子休眠是指在預定的時間范圍內本該有生命活力體征的種子,在預計正常、利于種子萌發的一些環境因子的組合條件下,不能按期正常萌發的一種生物現象[7]。休眠是植物的一種生存策略,不但可以為種子的傳播以及擴散爭取時間,還能促使種子在合適環境條件下萌發,可以有效地調節種子萌發的時空分布[8]。種子休眠對于植物而言是有利的生態適應特征,但卻成為人工育種的主要障礙,需要較長時間或者復雜的催芽方法才可令種子萌發,給草業生產造成一定程度的困難[9]。因此,尋找有效打破種子休眠的方法具有重要的現實意義。解除種子休眠的方法有多種:用物理方法去除種皮的硬實性,減少種皮對發芽的障礙,提高種子發芽率;化學物質和激素刺激種子發芽;清水漂洗和光照處理解除休眠;低溫層積加快種子后熟,促進種子發芽等[8-9]。有研究報道,適當的低溫預處理能夠打破種子休眠,提高萌發率[6]。運用低溫預處理的方法,研究種子發芽率和幼苗生長的變化,有利于探究和揭示植物種子的萌發機制。
青藏高原高寒草甸是我國高山草原的典型代表,具有溫度低、相對濕度高、生長季節短等特征[10]。黃帚橐吾(Ligulariavirgaurea)是菊科、橐吾屬多年生灰綠色草本植物,是一種對高寒草甸有著嚴重危害的毒害草,是高寒草地退化的標志性有毒植物之一[11],在青藏高原有著極強的入侵能力,對草地生產力和畜牧業發展造成了嚴重的影響[12]。黃帚橐吾體內可以產生特殊揮發物質,如萜類物質,抑制草原上其他植物種子的萌發及生長,使原本正常的草原退化,且其無性繁殖能力強大,如根莖繁殖,最終形成黃帚橐吾的單優勢種群[12]。有研究發現,黃帚橐吾種子在較大的溫度范圍內均可萌發,且光照、種子大小等均會對黃帚橐吾種子的萌發和幼苗生長產生不同程度的影響[13-14],也有研究發現黃帚橐吾種子的萌發率不高[4,13],具體原因可能是種子休眠難以被打破,也可能是種子萌發期間對外界環境條件過于敏感[15]。本研究以黃帚橐吾種子為對象,通過模擬高寒環境低溫或超低溫的條件,觀察黃帚橐吾種子萌發率、萌發速度和萌發后幼苗生長的變化,研究不同低溫預處理對黃帚橐吾種子萌發的影響,以期為毒害草型退化草地治理以及黃帚橐吾的有效防除提供理論依據。
1 材料和方法
1.1 種子來源
供試種子來自于青海省海北藏族自治州海晏縣青海湖鄉達玉德吉村(N 37°4′1″,E 100°52′48″),海拔3 230 m,氣候屬高原大陸性氣候,年均氣溫0.2~3.4 ℃,年均降水量277.8~499.5 mm,降水多集中在5-9月,年均日照2 580~2 750 h。草地類型以草甸化草原為主,以線葉嵩草(Kobresiacapillifolia)、矮嵩草(K.humilis)、垂穗披堿草(Elymusnutans)為主要建群種,伴生種為高原早熟禾(Poaalpigena)、中華羊茅(Festucasinensis)、扁蓿豆(Melissitusruthenica)等,牧草生長期4-5個月,土壤以高山草甸土為主。種子采集時間為2018年9月底和2019年9月底,兩次均于同一地點采集,采集區域面積為1.0 hm2,隨機采集種子,將黃帚橐吾花序剪下裝入樣袋,置于實驗室風干后手動將種子脫落,室溫下儲藏,溫度約為20 ℃。
1.2 試驗設計
于2020年1月初進行室內發芽試驗,觀察不同溫度預處理后黃帚橐吾種子萌發能力的變化。
本試驗設置了2個溫度預處理,分別為-20 ℃和4 ℃,-20 ℃下的種子處理時間分別為0.5、1和2 h,4 ℃下的種子處理時間分別為24、48和72 h。使用TP法進行種子萌發試驗,培養皿(直徑為9 cm)內放置2層干燥濾紙,用裝有蒸餾水的注射器潤濕濾紙,每個培養皿放置50粒種子。每個處理設置4個重復。置于光照培養箱中,變溫15~25 ℃(10 h/14 h)條件下培養,光照強度3 000 lx。試驗期間始終保持濾紙濕潤,并且每天記錄發芽種子數,試驗時間為17 d。最后,每個培養皿內隨機選取5株幼苗,測定并記錄根長(胚根延伸長度)和苗長(胚芽延伸長度),計算發芽指數和發芽率。
發芽指數:GI=∑(Gt/Dt)
式中,Gt為在時間t日的發芽數,Dt為相應的發芽天數[16]。
1.3 統計分析
所有數據均采用Excel計算并作圖,使用SPSS 20.0軟件進行統計分析。相同年份,不同處理之間的發芽指數、發芽率、根長和苗長的差異顯著性采用單因素方差分析方法進行檢驗。不同年份,相同處理之間的發芽指數、發芽率、根長和苗長的差異顯著性采用獨立樣本T檢驗,顯著性區間定義為95%水平。
2 結果與分析
2.1 4 ℃預處理對黃帚橐吾種子發芽率與發芽指數的影響
2018年采集的黃帚橐吾種子經過4 ℃預處理24、48和72 h后發芽率與對照相比均有所提高(圖1),其中,24 h處理下達到顯著水平(P<0.05)。2019年采集的黃帚橐吾種子經過4 ℃預處理24和48 h后,其發芽率顯著高于對照,處理72 h后種子發芽率與對照無顯著差異(P>0.05)。2019年采集的黃帚橐吾種子發芽率在各處理時間下均顯著高于2018年采集的種子(P<0.05)。
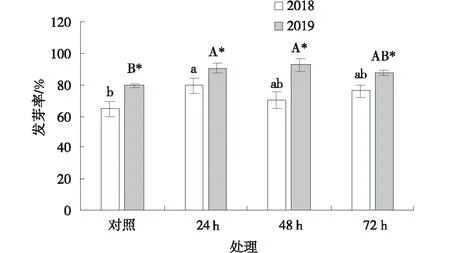
圖1 4 ℃預處理下黃帚橐吾種子發芽率Fig.1 Germination rate of Ligularia virgaurea seeds pretreated at 4 ℃注:*表示不同年份之間差異顯著(P<0.05),不同小寫字母表示2018年種子在不同處理時間之間差異顯著(P<0.05),不同大寫字母表示2019年種子在不同處理時間之間差異顯著(P<0.05)。下同
采于2018年的黃帚橐吾種子經過4 ℃預處理24 h和48 h后,其發芽指數與對照之間差異不顯著(P>0.05),而處理72 h后發芽指數顯著低于對照(P<0.05)(圖2)。2019年采集的黃帚橐吾種子經4 ℃處理24 h后與對照差異不顯著,處理48、72 h后,發芽指數顯著低于對照(P<0.05)。2019年采集的黃帚橐吾種子發芽指數高于2018年采集的種子,而經過4 ℃預處理后2個年份之間差異不顯著(圖2)。
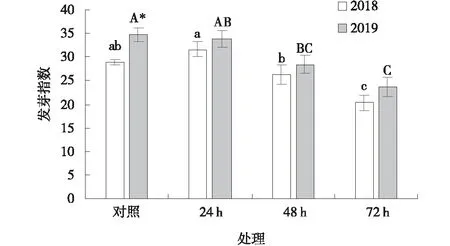
圖2 4 ℃預處理下黃帚橐吾種子發芽指數Fig.2 Germination index of Ligularia virgaurea seeds pretreated at 4 ℃
2.2 4 ℃預處理對黃帚橐吾種子根長與苗長的影響
采于2018、2019年的黃帚橐吾種子經過4 ℃預處理24 h后,其幼苗根長與對照差異不顯著(P>0.05),而處理48 h和72 h后,其根長顯著低于對照(P<0.05)(圖3)。2019年采集的黃帚橐吾種子根長在各處理時間下均顯著高于2018年采集的種子(P<0.05)。
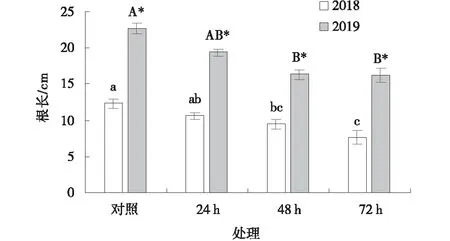
圖3 4 ℃預處理下黃帚橐吾種子根長Fig.3 Root length of Ligularia virgaurea seeds pretreated at 4 ℃
2018年采集的黃帚橐吾種子經過4 ℃預處理24、48 h后,種子苗長相比于對照沒有顯著差異(P>0.05),處理72 h后,其苗長顯著低于對照(P<0.05)(圖4)。采于2019年的黃帚橐吾種子經過4 ℃預處理24、48和72 h與對照相比均顯著降低(P<0.05)。2019年采集的黃帚橐吾種子苗長顯著高于2018年,經過4 ℃預處理后2個年份間無顯著差異(圖4)。

圖4 4 ℃預處理下黃帚橐吾種子苗長Fig.4 Seedling length of Ligularia virgaurea seeds pretreated at 4 ℃
2.3 -20 ℃預處理對黃帚橐吾種子發芽率與發芽指數的影響
2018年采集的黃帚橐吾種子經過-20 ℃預處理0.5、1和2 h 后發芽率與對照相比無顯著變化(P>0.05)。而2019年采集的種子經過-20 ℃處理0.5、1和2 h后發芽率均顯著高于對照(P<0.05)。2019年采集的黃帚橐吾種子發芽率在各處理下均顯著高于2018年采集的種子(圖5)。
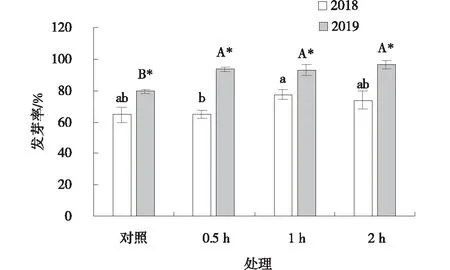
圖5 -20 ℃預處理下黃帚橐吾種子發芽率Fig.5 Germination rate of Ligularia virgaurea seeds pretreated at -20 ℃
2018年與2019年采集的黃帚橐吾種子經過-20 ℃預處理0.5 、1和2 h后發芽指數與對照相比均無顯著差異(P>0.05)。2019年采集的黃帚橐吾種子發芽指數高于2018年采集的種子(P<0.05),而經過-20 ℃預處理后,二者之間無顯著差異(圖6)。
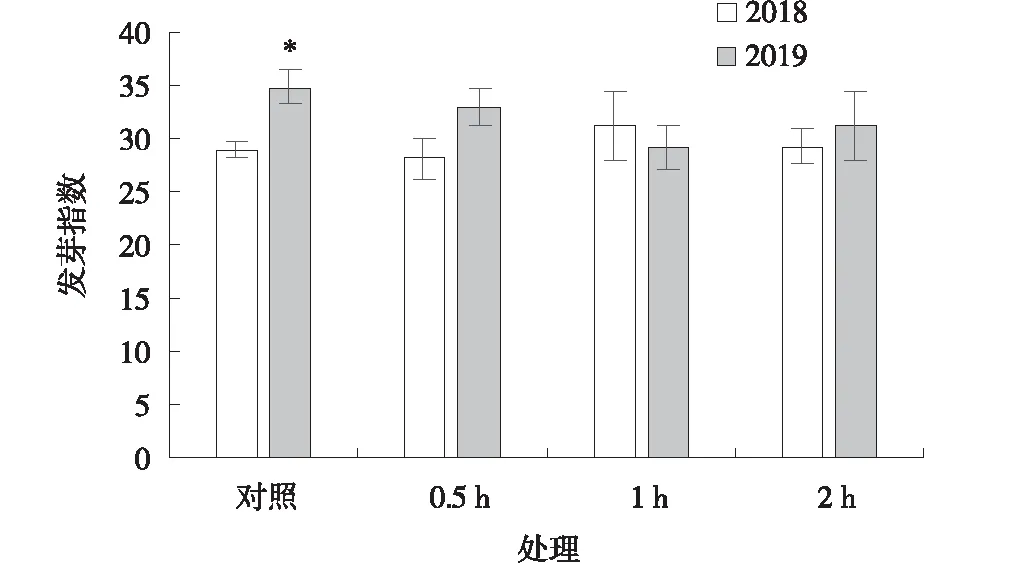
圖6 -20 ℃預處理下黃帚橐吾種子發芽指數Fig.6 Germination index of Ligularia virgaurea seeds pretreated at -20 ℃
2.4 -20 ℃預處理對黃帚橐吾種子根長與苗長的影響
采于2018年和2019年的黃帚橐吾種子經過-20 ℃預處理0.5、1和2 h后幼苗的根長與對照相比均無顯著變化(P>0.05)。在各處理下,2019年采集的黃帚橐吾根長均顯著高于2018年采集的種子(P<0.05)(圖7)。
2018年采集的黃帚橐吾種子經過-20 ℃預處理0.5 h后種子苗長與對照相比顯著提高(P<0.05)(圖8),處理1 h和2 h后相比于對照沒有顯著差異。2019年黃帚橐吾種子經過-20 ℃預處理0.5 h和1 h后相比于對照沒有顯著差異(P>0.05),處理2 h后相比對照顯著降低。2019年采集的黃帚橐吾的苗長顯著高于2018年(P<0.05),但是經過-20 ℃預處理后,2個年份之間差異不顯著(圖8)。

圖7 -20 ℃預處理下黃帚橐吾種子根長Fig.7 Root length of Ligularia virgaurea seeds pretreated at -20 ℃
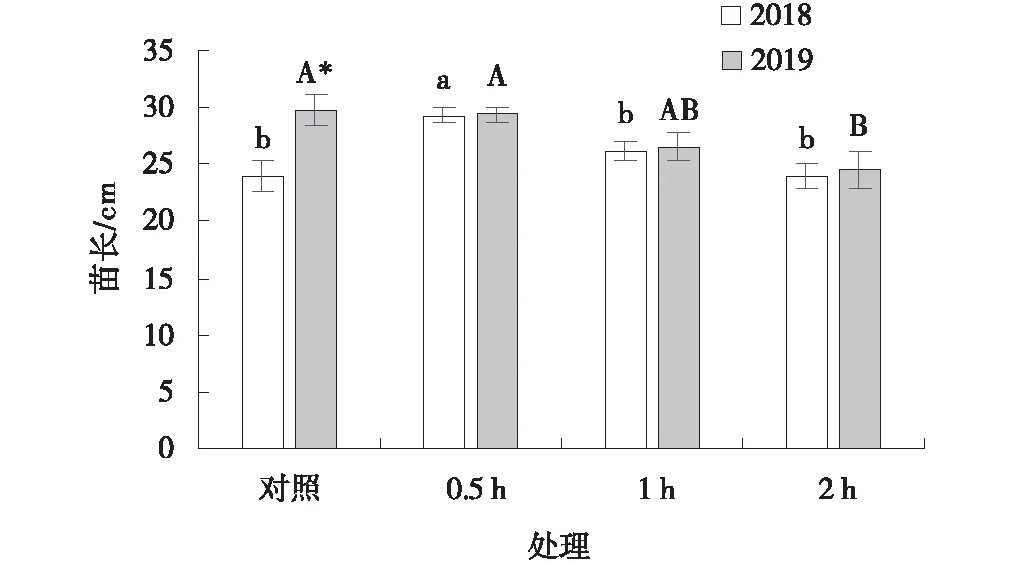
圖8 -20 ℃預處理下黃帚橐吾種子苗長Fig.8 Seedling length of Ligulariavirgaurea seeds pretreated at -20 ℃
3 討論
溫度過低,會抑制種子內酶的結構和代謝,從而影響酶的活性,最終抑制種子的正常代謝和發芽[17-18]、延長種子萌發時間[19],但低溫也可打破種子的熱休眠,促進種子萌發[20]。在不同情況下,低溫對種子發芽造成的影響也不同。本研究中黃帚橐吾種子的發芽率普遍不高,與之前的研究結果一致[4,21],這或許與黃帚橐吾種子自身的保護機制有關系[22]。也有研究結果證明,在生境不適宜種子萌發時,黃帚橐吾種子可能存在選擇性休眠,一部分隨著擴散媒介散播到其他生境中,而沒有擴散的種子當環境一旦適宜時就會萌發[14]。溫度的變化能促進種子胚的發育,進而提高種子的萌發率[23]。本研究中無論是4 ℃預處理還是-20 ℃預處理均提高了黃帚橐吾種子的發芽率,這可能是由于低溫預處理增加了黃帚橐吾種子的活力或促進了種子胚的發育,進而提高其發芽率,這是黃帚橐吾種子長期在高寒環境下生存而產生的適應性機制。另一方面,采于2019年的黃帚橐吾種子的發芽率普遍高于采自2018年的種子,有研究表明,黃帚橐吾種子儲存時間越長,萌發率越低[14],本研究與之一致。另外,2018年生長季降水量低于2019年,因此,同一時間采集的種子,2個年份之間的成熟度存在差異,這也是不同年份間黃帚橐吾種子發芽率有差異的原因之一。
發芽指數是發芽率指標的細化和深化,它放大了種子活力的特征[24]。本研究中,隨著4 ℃預處理時間的延長,黃帚橐吾種子的發芽指數逐漸降低,雖然短時間的低溫刺激會使種子煥發出活力,然而長時間的低溫預處理則使發芽的周期延長,但黃帚橐吾的種子仍然具有活力并最終萌發,這為其在多變的環境條件下的生存和擴張提供了保障[14]。前人的研究也發現低溫會延長種子的萌發持續時間[14,19],這是黃帚橐吾種子長期適應高寒環境形成的機制,通過延長種子的萌發周期確保在草地中占據更大的生存空間。對于不同年份采集的種子,本研究發現對照條件下2019年的黃帚橐吾種子的發芽指數高于2018年,而經過4 ℃與-20 ℃預處理后,兩個年份之間差異不顯著,表明黃帚橐吾種子儲存時間越長,種子的萌發活力可能越低,與之前的研究結果一致[14]。但是,低溫預處理對于長時間儲存的黃帚橐吾種子,起到了提高其萌發活力的作用,進而在萌發速度上減少了與新鮮種子之間的差距。
在植物一生中,種子萌發后的生長情況對植物后期的建植至關重要[13],種子萌發后其根長和苗長隨著外界環境脅迫強度的增加而降低[25]。之前的研究結果表明,植物種子的萌發和幼苗生長發育會因浸種初始溫度的不同而不同[26]。本研究發現經過4 ℃低溫處理后,隨著處理時間的延長,黃帚橐吾種子萌發后其根長和苗長均呈現降低的趨勢,而-20 ℃處理后,黃帚橐吾幼苗的根長和苗長與對照相比差異不大,甚至高于對照,這是由于黃帚橐吾種子連續處于低溫環境下(24、48或72 h),其萌發和生長活力受到了影響,種子根長和苗長隨著外界環境脅迫強度的增加而降低[13],而-20 ℃處理雖然溫度達到極低狀態,但處理時間不長(0.5、1或2 h),與其高寒的生存環境相近,一旦條件適宜,黃帚橐吾種子的發芽和生長還可能受到一定的促進。這與黃帚橐吾長期在高寒環境下生存形成的生存機制有關。值得注意的是,2019年采集的種子在-20 ℃預處理后,其苗長有所降低,這可能主要由于新采集的種子還未經過足夠時間的后熟等過程,應對外界環境突然變化的能力仍未發育完全,不能為幼苗的生長提供足夠的養分。因此,黃帚橐吾種子的萌發與生境的氣候、土壤、生長年份等均有密切聯系,具體的機制仍需要進一步研究。
4 結論
綜上所述,低溫預處理可以提高黃帚橐吾種子的發芽率;短期的低溫處理對于黃帚橐吾幼苗的生長有促進作用,對發芽指數影響不大;而低溫處理時間過長,則會對種子的發芽指數和幼苗的生長產生不利的影響;存儲時間越長,黃帚橐吾種子的萌發活力越低,但是,一定的低溫預處理能減緩存儲時間造成的影響。這對于黃帚橐吾種群在高寒草地群落中的長期生存和擴張可能發揮了有利作用。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
