蘋果MdSGR2基因克隆及植物超表達載體構建
2022-05-12 10:47:18趙惠英楊光凱張小軍郝燕燕
華北農學報 2022年2期
關鍵詞:植物
趙惠英,楊光凱,高 燕,2,張小軍,2,郝燕燕,2
(1.山西農業(yè)大學 園藝學院,山西 太谷 030801;2.果樹種質創(chuàng)制和利用山西省重點實驗室,山西 太原 030031)
果實色澤是衡量果實外觀品質的重要指標之一,主要由花青素、葉綠素和類胡蘿卜素含量決定[1]。蘋果果實色澤由底色和表色共同決定,其中底色主要受葉綠素和類胡蘿卜素含量影響,表色主要受花青素含量影響[2]。在果實成熟前,多數(shù)品種蘋果底色均呈綠色,隨著果實成熟果皮葉綠素不斷降解,使底色呈現(xiàn)黃白色,此現(xiàn)象稱為果實褪綠[3-4]。
植物葉綠素降解主要發(fā)生在葉片衰老和果實成熟過程中,其降解是一個多種酶共同調節(jié)的復雜過程。目前,已鑒定出6種葉綠素代謝酶(Chl catabolic enzymes,CCEs)調控葉綠素降解[5],其中,葉綠素b還原酶(Non-yellow coloring 1,NYC1)[6]、脫鎂葉綠酸a加氧酶(Pheophorbide a oxygenase,PAO)[7-8]、紅色代謝產物還原酶(Red chlorophyll catabolite reductase,RCCR)[9]等葉綠素降解關鍵酶在多種植物中已得到分離。此外,滯綠基因STAYGREEN(SGR)也是葉綠素降解的關鍵基因[10]。Armstead等[11]首次在豌豆中鑒定出SGR基因為葉綠素降解的關鍵調控因子。此后,陸續(xù)從許多植物中克隆鑒定了SGR基因,如水稻[12]、擬南芥[13]和番茄[14]。在高等植物中,SGR基因家族分為2個亞家族,包括SGR和SGR-LIKE(SGRL)亞家族[14],單子葉植物[15-16]和雙子葉植物[13,17]中都含有這2個家族。但在不同的植物類群中同源基因的數(shù)目不同,已有研究表明,擬南芥有3個SGR同源基因(AtSGR1/NYE1、AtSGR2、AtSGRL)[13],大豆有5個SGR同源基因(GmSGR1、GmSGR2、GmSGR3a、GmSGR3b、GmSGR4)[18],水稻有2個SGR同源基因(OsSGR、OsSGRL),柑橘有2個SGR同源基因(CsSGRa和CsSGRb)[19]。近年來,SGR基因的功能已在水稻[16]、柑橘[19]、擬南芥[20]、番茄[21-22]等物種中有了廣泛的研究。Zhu等[19]研究發(fā)現(xiàn),超表達CsSGRa顯著降低了煙草葉片中葉綠素含量,而超表達CsSGRb煙草葉片中葉綠素含量沒有明顯變化;CsSGRa突變抑制了棕橙果實和葉片中葉綠素的降解。Sakuraba等[20]研究發(fā)現(xiàn),AtSGR2負調控擬南芥葉片葉綠素的降解。Hu等[22]通過RNAi干擾沉默番茄中的SlSGR1,降低了番茄葉片和果實葉綠素的降解。但目前有關蘋果SGR基因的相關研究尚未見報道。
本研究以成熟澳洲青蘋蘋果果皮為試驗材料,克隆獲得MdSGR2基因,并對其編碼的蛋白結構特征進行了生物信息學分析,構建其植物超表達載體,旨在為進一步研究MdSGR2基因在蘋果果皮葉綠素降解中的生物學功能奠定基礎。
1 材料和方法
1.1 試驗材料
供試材料為澳洲青蘋成熟果實,種植于山西農業(yè)大學園藝試驗站。2019年10月下旬進行樣品采集,果實采收后迅速帶回實驗室,用人工打皮刀取胴體部位的果皮(果皮厚度約1 mm),錫箔紙包好后液氮速凍,-80 ℃保存待用。
1.2 試驗方法
1.2.1MdSGR2基因克隆 利用熱硼酸法提取蘋果果皮總RNA,經瓊脂糖電泳凝膠檢測以及核酸蛋白儀(Thermo ND2000C)測定濃度合格后,利用HiScript lI 1st Strand cDNA Synthesis Kit(+gDNA wiper)反轉錄試劑盒合成cDNA第一鏈。根據澳洲青蘋轉錄組測序結果分析得到MdSGR2基因的CDS序列(XM_029098422.1),利用Primer 5.0設計引物,即上游引物MdSGR2-F:5′-CGAGCTCATGGGCGTTTTGGCTGC-3′,下游引物MdSGR2-R:5′-GGGACTAGTCTAGTTTGATTTGCAC-3′,引物由北京擎科生物科技有限公司合成。以反轉錄得到的cDNA為模板,進行PCR擴增,擴增體系為:cDNA 1.0 μL(1 ng/μL),2×Phanta?Flash Master Mix 25.0 μL,引物(10 μmol/L)各2.0 μL,ddH2O 20.0 μL,共50.0 μL。擴增程序為:95 ℃預變性3 min;95 ℃變性15 s,56 ℃退火15 s,72 ℃延伸1 min,35個循環(huán);72 ℃延伸5 min,4 ℃保存。PCR產物經1.2%瓊脂糖凝膠電泳檢測回收后,與克隆載體pMD-18T連接,連接產物轉化大腸桿菌DH5α感受態(tài)細胞,經藍白斑篩選和菌落PCR檢測后,選取陽性單克隆送往北京擎科生物科技有限公司測序。
1.2.2MdSGR2基因生物信息學分析 利用ExPASY ProtParam tool(https://web.expasy.org/protparam/)在線工具分析MdSGR2的理化性質;MdSGR2蛋白的二級結構和三級結構分別利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和Phyre2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)進行預測;利用TMHMM 2.0 Server(http://www.cbs.dtu.dk/services/TMHMM/)在線軟件預測MdSGR2蛋白跨膜區(qū);利用PSORT Ⅱ Prediction(https://psort.hgc.jp/form2.html)在線軟件分析蘋果MdSGR2蛋白亞細胞定位;MdSGR2基因啟動子分析利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)完成。
1.2.3MdSGR2基因超表達載體的構建 將克隆載體pMD-MdSGR2和表達載體pCAMBIA2301分別用QuickCutTMSacⅠ和QuickCutTMSpeI酶進行雙酶切。雙酶切體系:質粒(0.5 ng/μL)4 μL;QuickCutTMSacⅠ酶1 μL;QuickCutTMSpeⅠ酶1 μL;Buffer 4 μL;ddH2O 30 μL。雙酶切產物經瓊脂糖凝膠電泳檢測后,用瓊脂糖凝膠DNA回收試劑盒(天根)分別對克隆載體目的片段和pCAMBIA2301空載體進行回收。隨后將目的片段和空載體按摩爾比1∶3的比例進行連接。連接體系為:目的基因1.7 μL,表達載體5.3 μL,T4連接酶1 μL,Buffer 2 μL,16 ℃過夜。將連接產物轉入大腸桿菌DH5α感受態(tài)細胞,經藍白斑篩選后,選取白色單菌落進行PCR驗證。挑取陽性單克隆送往北京擎科生物科技有限公司測序,將測序正確的菌液進行質粒提取并保存。
2 結果與分析
2.1 MdSGR2基因的克隆
以澳洲青蘋果皮cDNA為模板,使用同源克隆引物MdSGR2-F和MdSGR2-R進行PCR擴增,結果獲得與預期結果相符的目的片段(圖1),測序結果表明,目的基因cDNA序列為840 bp,編碼279個氨基酸。
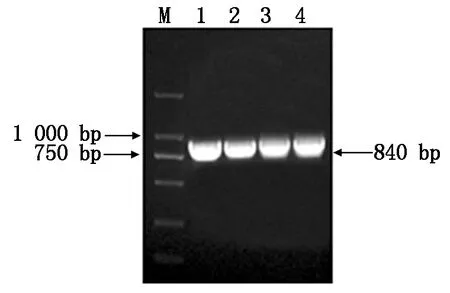
M.D2000 DNA Marker;1—4.MdSGR2基因PCR擴增產物。M.D2000 DNA Marker;1—4.PCR products of MdSGR2.
2.2 MdSGR2蛋白同源性分析
利用NCBI網站在線工具Blast檢索得到MdSGR2蛋白的同源蛋白,并進行同源性分析。利用DNAMAN進行氨基酸多序列比對,結果顯示,MdSGR2蛋白與秋子梨SGR蛋白具有較高同源性(圖2),其中與秋子梨(KAB2632630.1)同源性為84.12%,與甜櫻桃(XP 021803389.1)的同源性為72.83%,與桃(XP 007209459.1)的同源性為73.55%,與草莓(XP 004298724.1)的同源性為68.68%。

Md.蘋果;Pu.秋子梨;Pp.桃;Pa.甜櫻桃;Fa.草莓。Md.Malus domestica;Pu.Pyrus ussuriensis;Pp.Prunus persica;Pa.Prunus avium;Fa.Fragaria vesca subsp.vesca.
同時利用ClustalX軟件和MEGA 7.0軟件繪制蘋果與秋子梨、梅、桃、獼猴桃、草莓、杏、陸地棉等物種間的系統(tǒng)進化樹(圖3)。結果發(fā)現(xiàn),蘋果MdSGR2與秋子梨聚為一小分支,表現(xiàn)為親緣關系越近同源性越高。

圖3 蘋果MdSGR2與其他同源蛋白系統(tǒng)進化分析Fig.3 Phylogenetic analysis of MdSGR2 protein and other homologous proteins
2.3 MdSGR2基因編碼蛋白的理化性質分析
利用ExPASY網站在線軟件ProtParam對MdSGR2基因編碼蛋白質進行了理化性質分析,結果表明,MdSGR2基因編碼279個氨基酸,蛋白分子質量為31.27 ku,理論等電點pI為8.52,編碼的蛋白分子式為C1403H2175N391O400S11;由20種氨基酸組成,其中負電荷氨基酸殘基總數(shù)為29,正電荷氨基酸殘基總數(shù)為33;不穩(wěn)定系數(shù)為46.64,屬于不穩(wěn)定蛋白(小于40為穩(wěn)定);脂肪指數(shù)為82.19,總平均親水性為-0.291,屬親水性蛋白。
2.4 MdSGR2蛋白的二級結構和三級結構預測
采用SOPMA在線工具對MdSGR2蛋白二級結構進行了預測,結果顯示,其主要由α-螺旋、β-轉角、無規(guī)則卷曲和延伸鏈構成,其中,α-螺旋占40.50%,β-轉角占2.51%,延伸鏈占12.54%,無規(guī)則卷曲占44.44%,α-螺旋、無規(guī)則卷曲占的比例較大。采用在線軟件Phyre2預測了MdSGR2蛋白三級結構,結果顯示(圖4),其主要由α-螺旋和無規(guī)則卷曲構成,與二級結構預測結果相一致。
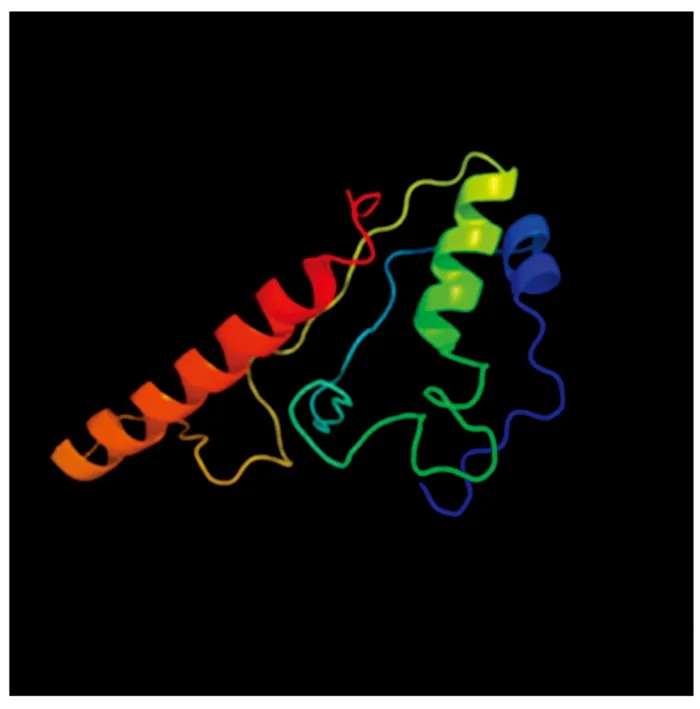
圖4 MdSGR2蛋白三級結構模型Fig.4 Tertiary structure model of MdSGR2 protein
2.5 MdSGR2蛋白結構域、跨膜區(qū)、亞細胞定位預測
根據NCBI的Conserved Domain Database(CDD)(https://www.ncbi.nlm.nih.gov/ Structure/cdd/wrpsb.cgi)對MdSGR2蛋白的結構域進行了預測,結果表明,該蛋白屬于Staygreen超家族,具有很高的保守性。
采用TMHMM 2.0 Server在線軟件預測了MdSGR2蛋白的跨膜區(qū),結果顯示,MdSGR2蛋白不具有跨膜區(qū),在胞外的可能性比較大,屬于胞外蛋白。
植物體中不同基因編碼的蛋白質有不同的存在部位,利用PSORT Ⅱ Prediction在線軟件分析了MdSGR2蛋白的亞細胞定位,結果表明,該蛋白定位在線粒體的概率為56.5%,定位在細胞質的概率為13.0%,定位在細胞核的概率僅有4.3%。
2.6 MdSGR2基因啟動子分析
利用PlantCARE對NCBI數(shù)據庫中查找到的MdSGR2基因轉錄起始位點上游2 000 bp的啟動子序列進行分析,查找到其所包含的順式作用元件,并對順式作用元件的種類、序列、功能和數(shù)量進行了分析,結果表明(表1),MdSGR2基因啟動子區(qū)域片段包含ABRE元件,表明該基因受脫落酸的誘導與調控;MdSGR2基因啟動子區(qū)域片段包含CGTCA-motif元件、TGACG-motif元件,表明MdSGR2基因參與茉莉酸甲酯的誘導反應;MdSGR2基因啟動子區(qū)域片段包含TCA-element元件,表明MdSGR2基因參與水楊酸反應;MdSGR2基因啟動子區(qū)域片段包含LTR元件,表明MdSGR2基因參與低溫反應。

表1 MdSGR2基因啟動子區(qū)順式作用元件及功能Tab.1 Cis-active elements and functions in the promoter region of MdSGR2
2.7 pCAMBIA2301-MdSGR2超表達載體構建
對克隆載體pMD-MdSGR2和超表達載體pCAMBIA2301分別進行雙酶切后(圖5-A、B),回收MdSGR2目的片段和超表達載體的雙酶切產物,連接轉化后,選取陽性菌落測序,將測序正確的菌液進行了質粒的提取,成功構建pCAMBIA2301-MdSGR2植物超表達載體(圖5-C)。
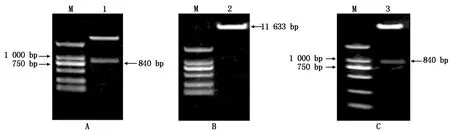
A.pMD-MdSGR2雙酶切;B.pCAMBIA2301載體雙酶切;C.超表達載體pCAMBIA2301-MdSGR2鑒定結果。M.D2000 DNA Marker;1.pMD-MdSGR2質粒雙酶切產物;2.pCAMBIA2301載體雙酶切產物;3.超表達載體pCAMBIA2301-MdSGR2雙酶切產物。
3 結論與討論
SGR蛋白參與葉片衰老和果實成熟,其對植物葉綠素降解至關重要。在高等植物中,SGR通過激發(fā)多個葉綠素降解酶與捕光復合物Ⅱ(Light-harvesting complex Ⅱ,LHCⅡ)相互作用,形成SGR-CCE-LHCⅡ復合體,進而調控葉綠素的降解[23]。蘋果葉綠素降解程度對果實色澤有重要影響,目前研究主要集中于不同植物激素處理對采后果實葉綠素降解代謝的影響[24-25],而有關SGR基因調控蘋果果實葉綠素降解尚未報道。本研究克隆獲得澳洲青蘋MdSGR2基因,結構域和序列比對分析表明,MdSGR2蛋白具有典型的Staygreen結構域,屬于Staygreen超家族,與青綠苔草[26]、番茄[27]、黑麥草[28]預測的結果相一致。亞細胞定位預測結果表明,該蛋白位于線粒體的概率最大,為56.5%。而前人在日本結縷草[29]、水稻[12]、蒺藜苜蓿[12]等SGR蛋白亞細胞定位的研究結果顯示,SGR蛋白均定位于葉綠體中,屬于典型的葉綠體蛋白。本研究預測結果與前人研究結果不一致。因此,后續(xù)還需進一步構建亞細胞定位載體35S-GFP∷MdSGR2并瞬時轉化煙草葉片,經共聚焦顯微鏡觀察進一步確定MdSGR2蛋白的亞細胞定位。
MdSGR2基因啟動子在線分析結果表明,該基因啟動子區(qū)域片段包含了脫落酸、水楊酸、茉莉酸甲酯多個激素響應元件。前人研究發(fā)現(xiàn),植物葉片衰老或果實成熟相關植物激素參與調控SGR基因的表達。張倩麗[30]研究表明,外源ABA處理促進了菊花腦葉片的衰老,且促進了CnSGR1基因的表達和葉綠素的降解。Yang等[31]研究表明,在ABA處理下,番茄中過表達SlSGRL加速了葉片的黃化;酵母雙雜試驗表明,SlSGRL主要通過與SlPPPH和SlCHa2相互作用參與ABA誘導的葉綠素降解。Tan等[32]研究發(fā)現(xiàn),外源施用茉莉酸甲酯(MeJA)加速了卷心菜葉片衰老,且BrPAO1、BrNYC1、BrPPH1和BrSGR1基因的表達量顯著提高。由此推斷,MdSGR2基因可能亦受果實成熟相關植物激素的調控,這為今后進一步研究ABA、MeJA等植物激素對MdSGR2基因表達的調控機制提供了思路。
目前,構建植物表達載體進行模式植物遺傳轉化是驗證目的基因生物學功能的有效手段。近年來,利用轉基因技術解析SGR基因調控植物葉綠素降解生物學功能的研究已有較多報道。劉凌云等[26]利用花序侵染法將青綠苔草滯綠基因CbSGR轉化擬南芥,過表達株系表現(xiàn)出黃化,加速了植株的衰老。檀鵬輝等[33]利用轉基因技術,將ZjSGR基因整合到煙草基因組中,轉基因煙草出現(xiàn)黃化現(xiàn)象,并且提高了煙草中SAG113、SAG12、NCED和AAO3衰老基因的表達量。周志湘等[34]研究發(fā)現(xiàn),ZjSGR基因轉化至匍匐剪股穎其葉綠素含量明顯加速下降。這些結果都表明,SGR基因參與調控多種不同植物中葉綠素降解代謝,因此,本研究構建了植物超表達載體pCAMBIA2301-MdSGR2,進一步探究MdSGR2基因在葉綠素降解中的作用。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
