施氮對高溫脅迫下黃瓜果期熒光特性的影響
2022-05-12 10:47:22石嘉琦劉宇慶王艷玲楊再強
華北農學報 2022年2期
關鍵詞:產量
石嘉琦,劉宇慶,王艷玲,楊再強
(南京信息工程大學 應用氣象學院,江蘇 南京 210044)
夏季設施內溫度頻繁高于35 ℃嚴重影響了設施黃瓜的產量與品質[1]。有研究發現,高溫脅迫下黃瓜花器官發育不良、蕾期黃化凋萎、化果數增加而結實率下降[2];而42 ℃高溫處理7 d后黃瓜植株則無果實[3]。如何有效緩解高溫脅迫對設施黃瓜的損害一直是設施栽培中關注的焦點問題。高溫脅迫下植物葉片光系統Ⅱ(PSⅡ)反應中心最為敏感,而通過快速葉綠素熒光誘導曲線分析方法(JIP-test)可有效判斷高溫脅迫下PSⅡ反應中心光合作用過程的變化,因此,常常被用于作物逆境生理的研究[4-5]。徐向東等[6]研究發現,42 ℃高溫脅迫下,黃瓜葉片PSⅡ的初始熒光(Fo)顯著增加,而最大熒光(Fm)和最大光化學量子產率(Fv/Fm)則顯著降低,葉片PSⅡ受體側受傷,電子傳遞能力降低。醌庫容量(Sm)的大小可以反映電子受體QA完全被還原所需要能量的高低,而高溫脅迫下光葉眼子菜的Sm顯著變小,電子傳遞速率顯著降低[7]。高溫脅迫導致PS Ⅱ供體側受到傷害后,在極短時間內葉綠素熒光強度就會顯著上升,且在0.3 ms出現拐點K;而植物葉片中放氧復合體(OEC)活性的變化可以有效表征PS Ⅱ供體側的損傷程度[8]。有研究發現,35 ℃高溫脅迫導致番茄葉片OEC的功能受損,使得K點的相對可變熒光值較25 ℃顯著增大[9]。高溫脅迫下作物葉片的吸收光能(ABS/RC)、捕獲光能(TRo/RC)、電子傳遞的能量(ETo/RC)等光合機構的能量與單位反應中心的比值(比活性)也會明顯下降[10]。
氮是參與葉片葉綠素合成與分解、光合作用、核酮糖-1,5-二磷酸羧化酶(Ru BPCase)活性等光合作用各個環節的重要營養元素;氮素供應直接或間接地影響著作物的光合作用,調控著光合電子傳遞鏈蛋白、天線色素蛋白復合體和光合酶的組成[11];而作物對氮素的吸收和利用影響著葉片對光能的吸收、轉換、類囊體膜上的電子傳遞及光合磷酸化的能力[12-13]。有研究發現,高溫脅迫下,氮素添加可以促進水稻葉片葉綠素含量的增加,增強葉肉細胞光合活性及葉片吸光強度,保證光合作用的有序進行[14];而合理的氮素供應可以通過能量調節使葉片最大限度利用吸收的光能,在一定程度上減輕高溫導致的過多激發能對光合器官的破壞,從而使PSⅡ的Fv/Fm保持在較高的水平上,提高了作物葉片對高溫逆境的適應性[15-17]。可見,氮素的合理施用是高溫脅迫下有效減緩作物葉片光合機構功能受損的潛在有效措施之一,而關于氮素調控減緩設施黃瓜高溫迫害的熒光機制研究鮮有報道。
因此,本研究基于盆栽試驗,分析了高溫脅迫與不同氮素供應水平下津優101號黃瓜葉片葉綠素含量、熒光產量、光化學活性、PSⅡ反應中心活性、性能指數及熒光差異動力學曲線的變化差異與機制,研究結果可以為設施黃瓜高溫災害防御及氮素優化管理提供科學依據及實踐指導。
1 材料和方法
1.1 試驗設計
試驗于2020—2021年在南京信息工程大學農業氣象試驗站(E118°71′,N32°21′)Venlo型試驗玻璃溫室和智能人工氣候箱(TPG1260,澳大利亞)中進行。在試驗站內選擇地勢平坦田塊,按“S”形采集表層土壤(0~10 cm)15~20點,混合均勻后,取約1 500 kg土壤樣品、風干,挑揀出大的根系與石礫后,磨細過5 mm篩備用。供試品種為黃瓜津優101號(Jinyou 101),采用盆栽種植,盆上口直徑為24.0 cm,盆底直徑為19.5 cm,高為26.5 cm,每盆裝過5 mm篩的風干土8.0 kg。每盆播種3粒,待黃瓜長出四葉一心時,保留一棵壯苗,而后開始進行施氮處理,試驗設0(N0),160(N1),240(N2),320(N3) kg/hm2等4個氮素水平,每個水平重復3次,整個生育期P2O5與K2O的用量分別為90,180 kg/hm2。所施氮肥為尿素(N≥46.4%),磷肥為過磷酸鈣(P2O5≥12%),鉀肥為氯化鉀(K2O≥60%),所有肥料分別在幼苗期(20%)、花期(20%)、果期(60%)水溶解后施入盆中。
待黃瓜第1棵根瓜坐住后,將盆栽放入智能人工氣候箱中進行高溫處理,以28 ℃(日最高氣溫)/18 ℃(日最低氣溫)為對照,設計35 ℃/25 ℃、38 ℃/28 ℃、41 ℃/31 ℃共3個高溫水平,日最高氣溫和逐時氣溫參考南京地區高溫季節日變化設置[18],每小時的溫度通過程序控制。光合有效輻射6:00—18:00設定(PAR)為1 000 μmol/(m2·s),其他時間段為0 μmol/(m2·s),空氣濕度均設定為50%~55%,高溫處理9 d后用植物效率分析儀測定黃瓜葉片熒光參數并繪制葉綠素熒光誘導曲線(OJIP),高溫處理結束10 d后統計黃瓜產量。
1.2 測定指標及方法
1.2.1 土壤理化性質和葉綠素含量的測定 土壤理化性質參考魯如坤[19]方法測定,營養土pH值采用1.0 mol/L KCl溶液浸泡,電位法測定,液土比為2.5∶1.0;有機質采用高溫外熱重鉻酸鉀氧化-容量法測定;土壤總氮采用H2SO4-H2O2消解后,用全自動凱氏定氮儀(K9860)測定消解液中氮含量;葉綠素含量采用紫外分光光度計法測定[20]。
1.2.2 熒光參數測定及快速葉綠素熒光誘導(OJIP)曲線的繪制 葉綠素熒光采用植物效率分析儀(Pocket PEA,英國)測定,每個處理重復測定3株,測定項目包括初始熒光(Fo)、最大熒光(Fm)、最大光化學量子產量(Fv/Fm),光合性能指數(PIabs)、綜合性能指數(PItotal)、Fm與熒光曲線圍成的面積(Area)、醌庫容量(Sm)、單位活性反應中心吸收的光能(ABS/RC)、熱耗散的能量(DIo/RC)、捕獲的用于還原QA的光能(TRo/RC)和用于電子傳遞的能量(ETo/RC),并將熒光強度值與響應時間對數值進行擬合后,以時間對數值為橫坐標繪制OJIP曲線[21]。為了進一步比較不同施氮處理下黃瓜葉片葉綠素熒光誘導曲線的特異性,將不同時間段內的曲線標準化為相對可變熒光(一般用W表示),即將O點的相對熒光強度定義為0,分別將K、J、I和P點的相對熒光強度定義為1進行標準化,并計算O-K和O-J的相對可變熒光的差值(ΔW)[22]。
1.3 數據分析
數據統計分析采用Microsoft Excel 2016和SPSS 20.0軟件,繪圖采用Origin 2017和GraphPad Prism 8.0軟件。
2 結果與分析
2.1 果期黃瓜葉片葉綠素含量的變化差異
與對照溫度相比,果期高溫脅迫下N3處理黃瓜葉片Chla和Chla+Chlb分別顯著增加了25.2%~39.4%和40.5%~51.9%(P<0.05);35 ℃脅迫下N1處理黃瓜葉片Chla、Chlb和Chla+Chlb分別顯著增加43.5%,67.0%,49.9%(P<0.05),38,41 ℃脅迫則無顯著變化;果期高溫脅迫下N2處理黃瓜葉片Chla、Chlb和Chla+Chlb均無顯著變化。與N0處理相比,施氮可以使果期高溫脅迫下黃瓜葉片Chla、Chlb和Chla+Chlb顯著增加1.1~4.3倍(P<0.05)(圖1)。
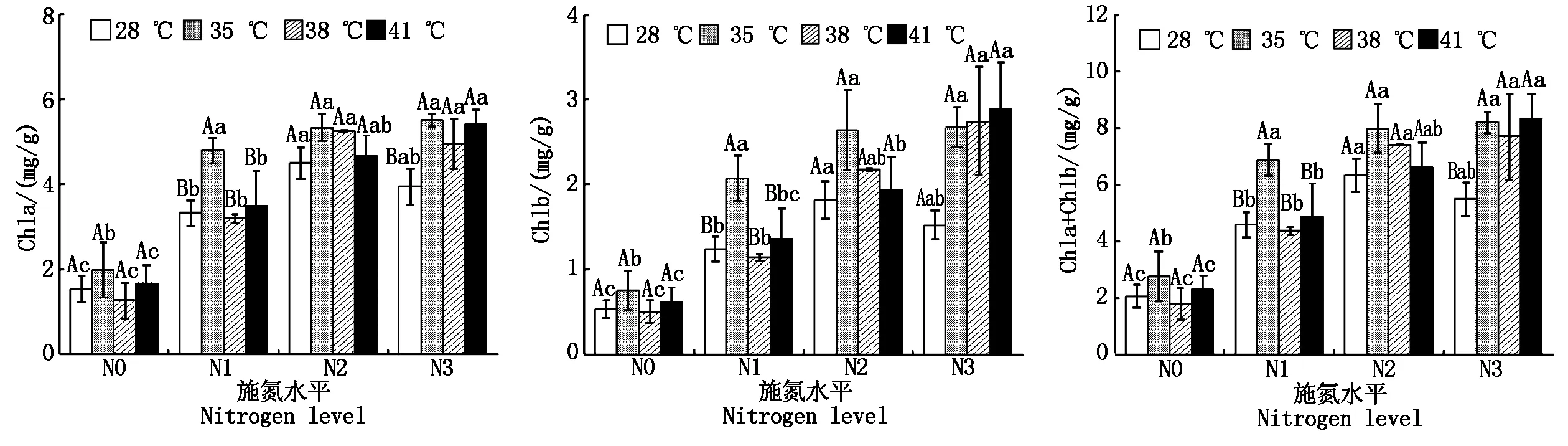
不同小字母表示不同施氮處理間差異顯著(P<0.05);不同大寫字母表示不同溫度間差異顯著(P<0.05)。圖2—4,9同。Different lowercase letter means significant difference in different N treatments(P<0.05);the different uppercase letter mean significant difference in different growth temperatures(P<0.05).The same as Fig 2—4,9.
2.2 果期黃瓜葉片葉綠素熒光參數的變化差異
2.2.1 初始熒光(Fo)和最大熒光(Fm) 與對照溫度相比,果期高溫脅迫下施氮處理的黃瓜葉片Fm顯著降低了6.4%~30.7%(P<0.05);38 ℃脅迫下N0、N1處理的黃瓜葉片Fo分別顯著降低了18.7%,18.5%(P<0.05),41 ℃脅迫下N1處理的黃瓜葉片Fo顯著降低了9.8%(P<0.05),其他處理則無顯著變化(圖2)。與N0處理相比,N2、N3處理使35 ℃脅迫下黃瓜葉片的Fm和Fo分別顯著提高了49.6%,52.5%,14.6%,17.6%(P<0.05),但N2、N3處理間差異不顯著;N3處理使38 ℃脅迫下黃瓜葉片的Fm顯著提高了28.8%(P<0.05),N2處理則使Fo顯著提高了18.4%(P<0.05),其他處理無顯著變化;N2、N3處理使41 ℃脅迫下黃瓜葉片的Fm顯著提高了16.0%,14.9%,Fo無顯著變化(圖2)。
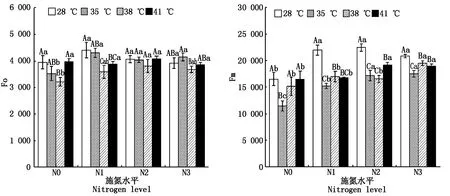
圖2 果期高溫脅迫下施氮水平對黃瓜葉片Fo、Fm的影響Fig.2 Effect of nitrogen added on the Fo and Fm of cucumber leaves under high temperature stress in fruit stage
2.2.2 光化學量子產率(Fv/Fm)和性能指數 與對照溫度相比,果期高溫脅迫下N2處理黃瓜葉片的Fv/Fm顯著降低了5.1%~6.7%(P<0.05);35,41 ℃脅迫下N1處理黃瓜葉片的Fv/Fm分別顯著降低了10.3%,3.9%(P<0.05);35 ℃脅迫下N3處理黃瓜葉片的Fv/Fm顯著降低了6.2%(P<0.05),其他處理無顯著變化(圖3-A)。與N0處理相比,N2、N3處理使35 ℃脅迫下黃瓜葉片Fv/Fm顯著提高了10.2%~10.4%(P<0.05);N3處理使38 ℃脅迫下黃瓜葉片Fv/Fm顯著提高了3.2%(P<0.05);N2與N3處理使41 ℃脅迫下黃瓜葉片Fv/Fm分別顯著提高了3.9%,5.1%,其他處理無顯著變化(圖3-A)。
與對照溫度相比,果期35 ℃脅迫下N1~N3處理的黃瓜葉片PIabs顯著降低了32.1%~70.2%(P<0.05),38 ℃脅迫下N2處理和41 ℃脅迫的N1處理的黃瓜葉片PIabs分別顯著降低了9.3%,51.1%(P<0.05),其他處理無顯著變化。與N0處理相比,N2、N3處理使35,41 ℃脅迫下黃瓜葉片的PIabs顯著提高了9.3,9.1,3.4,4.9倍,使葉片的PItotal顯著提高了12.0,9.0,4.3,4.2倍(P<0.05);N3處理使38 ℃脅迫下果期黃瓜葉片的PIabs和PItotal分別顯著提高了0.9,1.2倍(P<0.05)(圖3-B、C)。
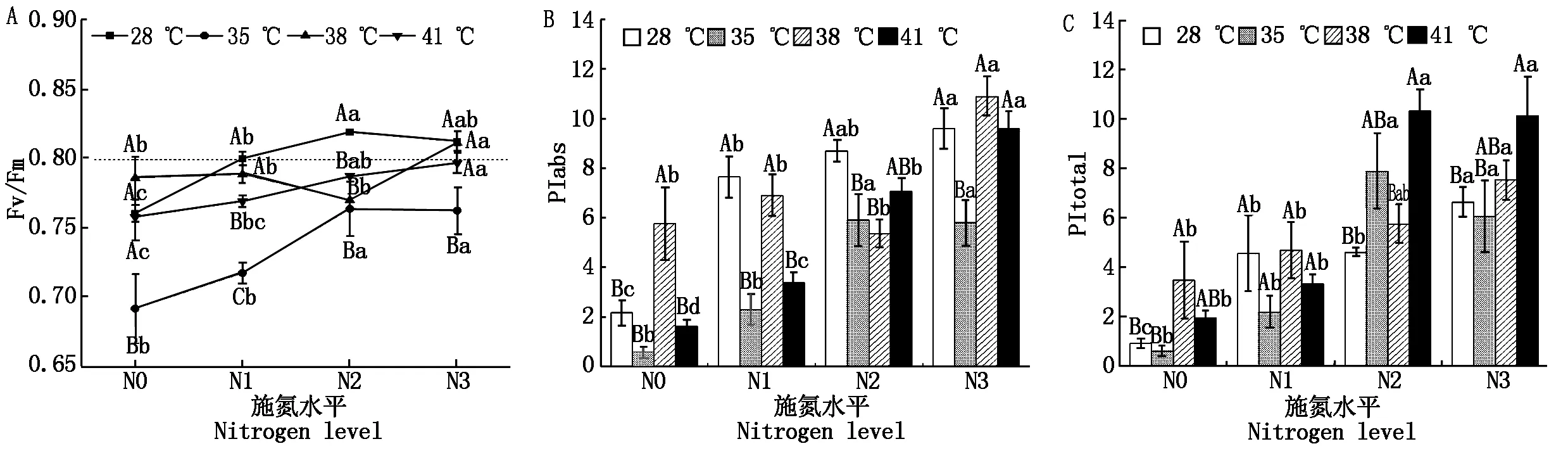
圖3 果期高溫脅迫下施氮水平對黃瓜葉片PSⅡ光化學活性的影響Fig.3 Effect of nitrogen added on the PSⅡ photochemical activity of cucumber leaves under high temperature stress in fruit stage
2.2.3 PSⅡ反應中心活性 與對照溫度相比,35 ℃脅迫下N1處理的黃瓜葉片ABS/RC、DIo/RC分別顯著提高了16.6%,65.0%,ETo/RC則顯著降低了17.0%(P<0.05),TRo/RC無顯著變化;38 ℃脅迫下N2處理的黃瓜葉片ABS/RC、DIo/RC分別顯著提高了19.3%,51.5%(P<0.05),N3處理的TRo/RC、ETo/RC則分別顯著降低了18.3%,19.8%(P<0.05);41 ℃脅迫下N1處理的黃瓜葉片僅ETo/RC顯著降低了16.8%(P<0.05),其他指標無顯著變化。與N0處理相比,N2處理使35,41 ℃脅迫下黃瓜葉片的ABS/RC、DIo/RC、TRo/RC分別顯著降低了46.0%,59.0%,40.4%和18.0%,28.0%,14.4%(P<0.05),ETo/RC則提高了16.7%,40.9%;N3處理使38 ℃脅迫下黃瓜葉片的ABS/RC、DIo/RC和TRo/RC分別顯著降低了24.0%,33.4%,20.8%(P<0.05),ETo/RC無顯著變化(表1)。
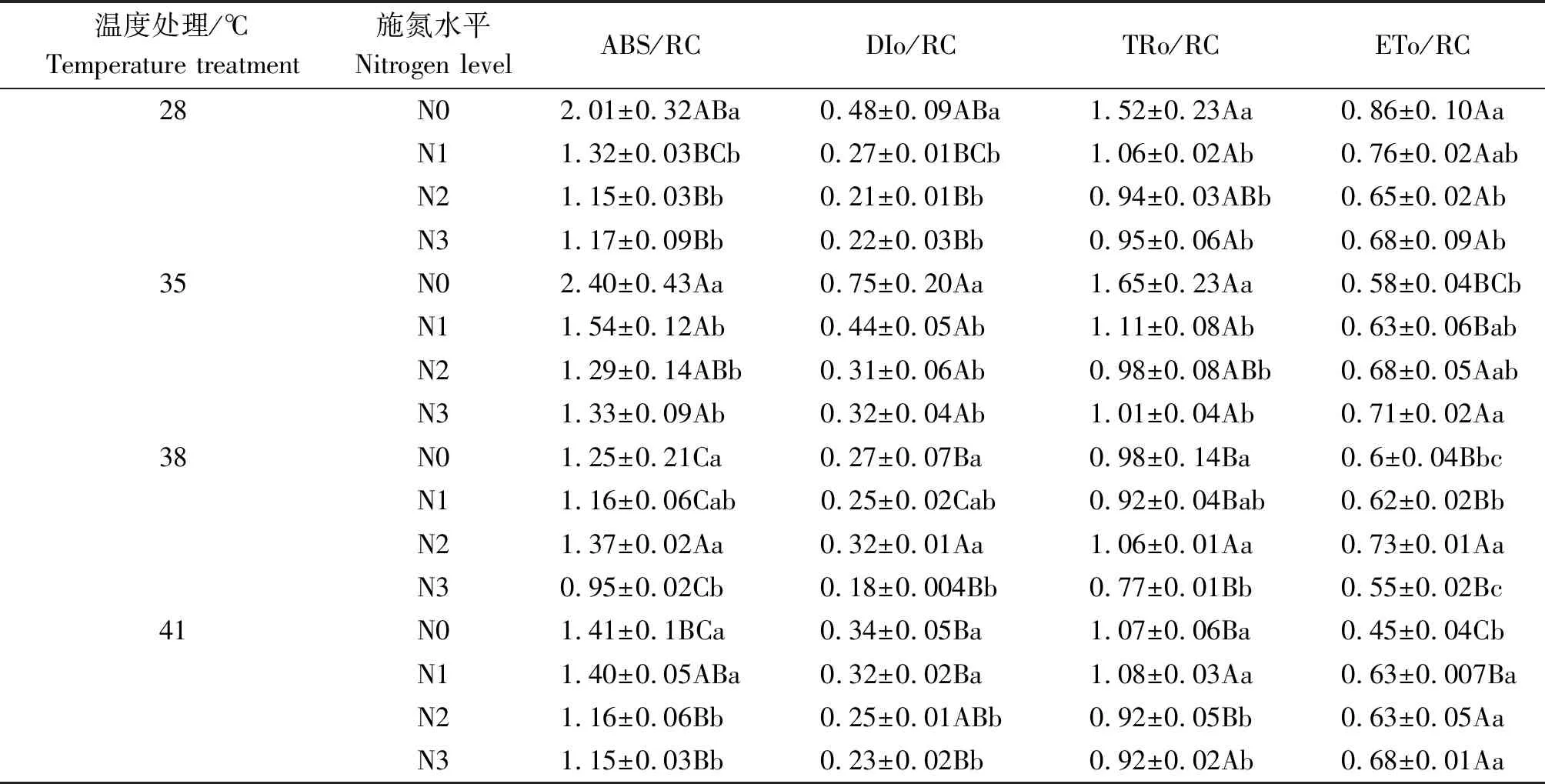
表1 果期高溫脅迫下施氮水平對黃瓜葉片PSⅡ反應中心活性參數的影響Tab.1 Effect of nitrogen added on the PSⅡ photochemical activity in reaction center of cucumber leaves under high temperature stress in fruit stage
2.2.4 PSⅡ受體側醌庫容量 與對照溫度相比,果期高溫脅迫下N1處理的黃瓜葉片Area顯著降低了36.2%~43.2%(P<0.05),35 ℃脅迫下N3處理和38 ℃脅迫下N2處理的黃瓜葉片Area則分別顯著降低了24.7%,22.6%;38,41 ℃高溫脅迫下N1處理的黃瓜葉片Sm分別顯著降低了16.7%,22.9%(P<0.05),而35,41 ℃脅迫下N2處理的黃瓜葉片Sm則分別顯著增加了25.1%,27.7%(P<0.05),其他處理無顯著變化。與N0處理相比,N2、N3處理使35 ℃脅迫下黃瓜葉片的Area和Sm分別顯著提高了2.8,2.7,1.3,1.2(P<0.05)倍;N3處理使38 ℃脅迫下黃瓜葉片的Area和Sm顯著提高了1.3,0.7(P<0.05)倍;N2、N3處理使41 ℃脅迫下黃瓜葉片的Area和Sm分別顯著提高了1.6,1.7倍和1.1,1.3(P<0.05)倍,且N2和N3處理間差異不顯著(圖4)。
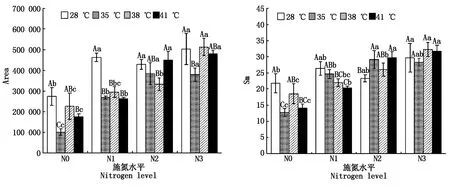
圖4 果期高溫脅迫下施氮水平對黃瓜葉片Area和Sm的影響Fig.4 Effect of nitrogen added on the Area and Sm of cucumber leaves under high temperature stress in fruit stage
2.3 果期黃瓜葉片葉綠素熒光動力學曲線變化
2.3.1葉綠素熒光動力學曲線 與對照溫度相比,果期高溫脅迫下黃瓜葉片I點(30 ms)上升緩慢且P點(200 ms)的葉綠素熒光強度(FI)均有不同程度的降低;各溫度下N0處理的黃瓜葉片FJ(2 ms處熒光強度)均變大且在0.3 ms處出現拐點K,I點(30 ms)上升緩慢且P點的FI最低,施氮后各溫度處理的黃瓜葉片I點和P點的FI均得到提高,其中35,38 ℃脅迫下N3水平的Ⅰ點上升較快;41 ℃脅迫下N2水平的I點上升較快(圖5)。
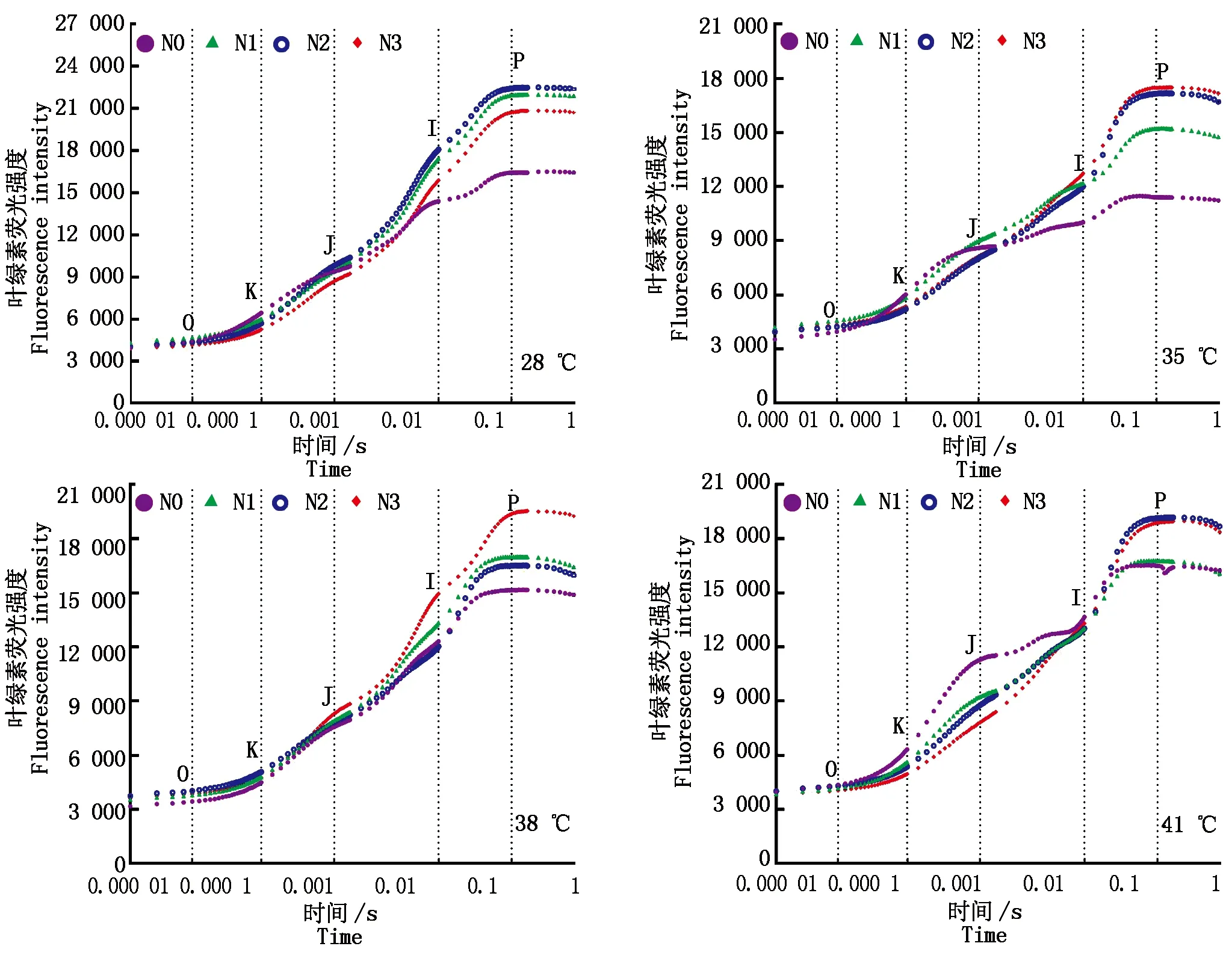
圖5 果期高溫脅迫下施氮水平對黃瓜葉片OJIP曲線的影響Fig.5 Effect of nitrogen added on the OJIP curve of cucumber leaves under high temperature stress in fruit stage
2.3.2 放氧復合體(OEC) ΔWO-J為正值表明OEC失活,為負值表明OEC未失活。由圖6可知,對照溫度和35 ℃脅迫下,各施氮處理的果期黃瓜葉片的ΔWO-J均為負值,OEC未失活;38 ℃脅迫下N2處理的ΔWO-J在0.05~0.70 ms出現正值,OEC出現失活現象,N1、N3處理的OEC未失活,且N3處理OEC活性最強;41 ℃脅迫下N1處理的黃瓜葉片的ΔWO-J在0.05~ 0.30 ms出現正值,OEC輕微失活,N2和N3處理黃瓜葉片的OEC未失活。
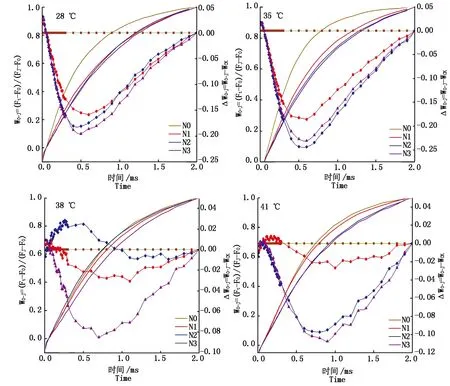
圖6 果期高溫脅迫下施氮水平對黃瓜葉片OJ相熒光差異動力學的影響Fig.6 Effect of nitrogen added on the fluorescence differential kinetics of OJ phase of cucumber leaves under high temperature stress in fruit stage
2.3.3 類囊體間能量傳遞 ΔWO-K為正值說明類囊體之間的能量傳遞受阻,為負值說明光合膜系統完整,能量傳遞順暢。由圖7可知,對照溫度下N2處理的果期黃瓜葉片ΔWO-K始終為負值,類囊體之間的能量傳遞順暢,N1和N3處理的黃瓜葉片ΔWO-K在0.05~0.07 ms出現正值,能量傳遞輕微受阻;35 ℃脅迫下N2處理的ΔWO-K總體為負值且低于N1和N3處理,能量傳遞較為順暢;38,41 ℃脅迫下各施氮處理的果期黃瓜葉片的ΔWO-K整體均為正值,ΔWO-K由低到高排列依次為N3>N1>N2和N2>N1>N3,說明果期38,41 ℃脅迫導致PSⅡ能量連接性受到嚴重阻礙,N3和N2處理分別可以有效增強類囊體之間能量的連接性。
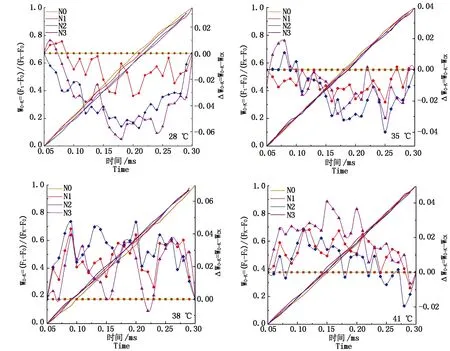
圖7 果期高溫脅迫下施氮水平對黃瓜葉片OK相熒光差異動力學的影響Fig.7 Effect of nitrogen added on the fluorescence differential kinetics of OK phase of cucumber leaves under high temperature stress in fruit stage
2.3.4 PSⅠ末端電子受體 WO-I≥1部分I~P相的振幅反映了PS Ⅰ受體側末端電子受體庫的大小,I~P相振幅越小,則表明PSⅠ受體側末端電子受體庫所受抑制越強,WI-P=0.5(上升曲線達到半值)處所需時間點可用來反映PSⅠ末端電子受體庫的還原速率。由圖8可知,對照溫度下施氮可以提高果期黃瓜葉片WO-I≥1部分I~P相的振幅,減緩PSⅠ受體側末端電子受體庫的抑制狀況,加快PSⅠ末端電子受體庫的還原速率,其中N3處理的I~P相的振幅最大,緩解作用最好,N2處理的PSⅠ末端電子受體庫的還原速率最快。果期高溫脅迫下施氮可以增大黃瓜葉片WO-I≥1部分I~P相的振幅,減緩PSⅠ受體側末端電子受體庫的抑制狀況,其中N2處理效果最好;但各施氮處理均增加了果期高溫脅迫下黃瓜葉片的半衰期,降低了PSⅠ末端電子受體庫的還原速率。
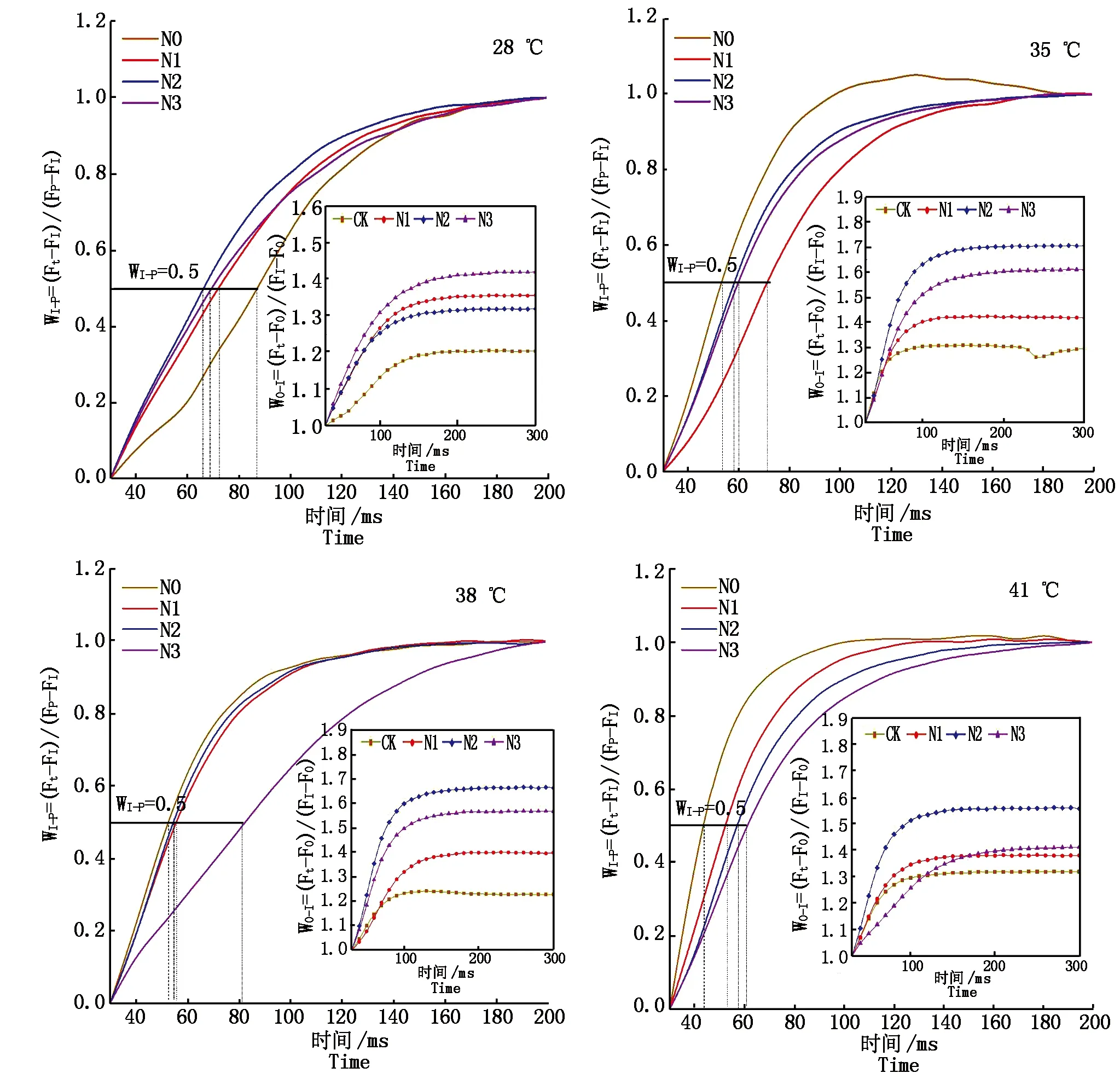
圖8 果期高溫脅迫下施氮水平對黃瓜葉片OI和IP相熒光差異動力學的影響Fig.8 Effect of nitrogen on the fluorescence differential kinetics of OI and IP phase of cucumber leaves under high temperature stress in fruit stage
2.4 黃瓜產量對施氮量及高溫脅迫的響應
與對照溫度相比,果期35 ℃高溫脅迫下N1處理的黃瓜產量降低了38.1%,38 ℃高溫脅迫下N0處理的黃瓜產量降低了31.5%,而N3處理的黃瓜產量顯著增加了35.5%(P<0.05),41 ℃高溫脅迫下N0~N3處理的黃瓜產量顯著降低了55.5%~71.2%(P<0.05),其他處理無顯著變化。與N0處理相比,果期35,38,41 ℃高溫脅迫下施氮處理的黃瓜產量分別顯著增加了87.3%~101.1%,349.2%~422.5%,71.1%~209.6%(P<0.05)。將各溫度下的產量數據與施氮量進行擬合后發現黃瓜獲得最高產量時的施氮量隨著溫度的升高呈先上升后降低趨勢,28 ℃處理下施氮量為195 kg/hm2,黃瓜獲得最高產量12.5 t/hm2,35 ℃脅迫下施氮量為236 kg/hm2時,黃瓜獲得最高產量8.6 t/hm2,38 ℃脅迫下施氮量為283 kg/hm2,黃瓜獲得最高產量12.3 t/hm2,41 ℃脅迫下當施氮量為177 kg/hm2,黃瓜獲得最高產量4.9 t/hm2(圖9)。
2.5 綜合分析
相關分析表明,產量與施氮量、Fm、Area、Sm、PIabs、Chla和總葉綠素含量均存在極顯著正相關關系(P<0.01),與Fv/Fm、Chlb存在顯著正相關關系(P<0.05),與溫度(≥28 ℃)存在極顯著負相關關系(P<0.01),與ABS/RC、DIo/RC、TRo/RC存在顯著負相關關系(P<0.05),與Fo、PItotal和ETo/RC無顯著相關關系(表2)。
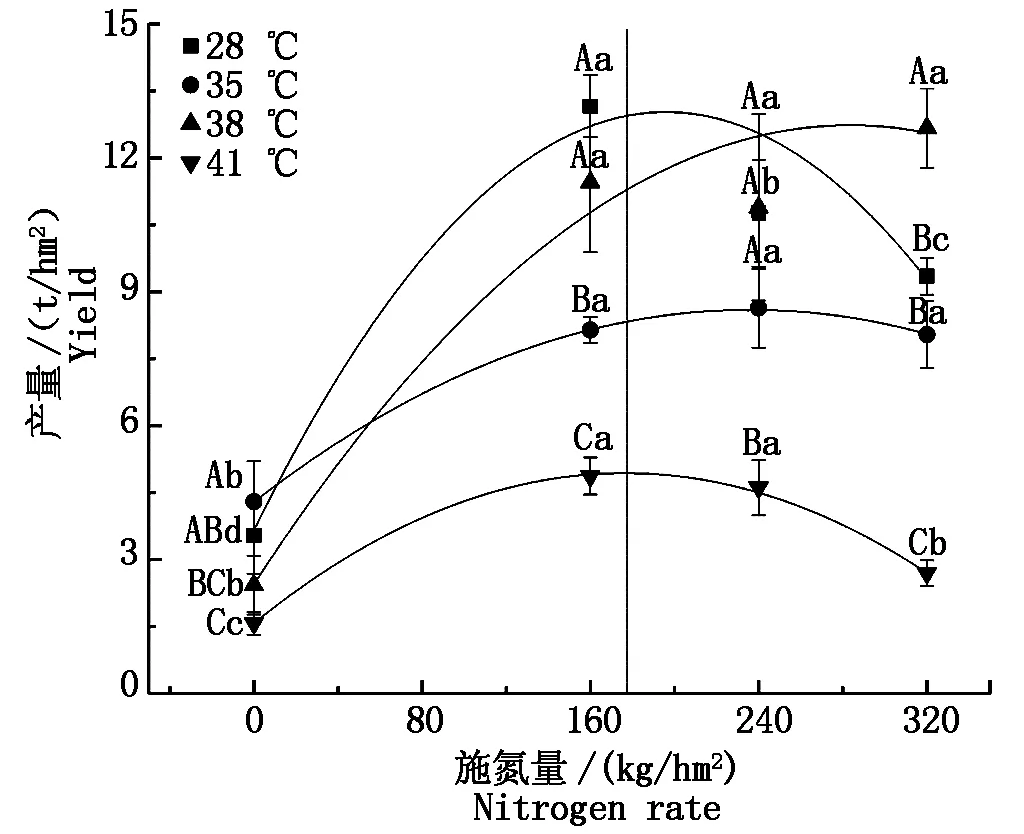
圖9 果期高溫脅迫下施氮水平對黃瓜產量的影響Fig.9 Effect of nitrogen added on yield of cucumber under high temperature stress in fruit stage
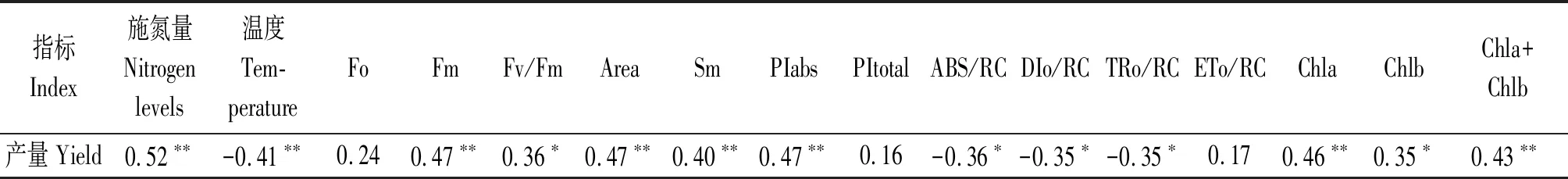
表2 產量與施氮量、溫度、黃瓜葉片綠素含量、各熒光指標間的相關分析Tab.2 Correlation analysis between yield and nitrogen levels,temperature,chlorophyll content and fluorescence indexes in cucumber leaves
方差分析結果顯示,施氮量對黃瓜產量、葉片Fm、Fv/Fm、Area、Sm、PIabs、PItotal、ABS/RC、DIo/RC、TRo/RC、Chla、Chlb和Chla+Chlb等影響均達到極顯著水平(P<0.01),對葉片Fo的影響顯著(P<0.05),而對葉片ETo/RC影響不顯著(P>0.05)。溫度對黃瓜產量、葉片Fo、Fm、Fv/Fm、Area、PIabs、ABS/RC、DIo/RC、TRo/RC、ETo/RC、Chla和Chla+Chlb等影響達到極顯著水平(P<0.01),對葉片PItotal、Chlb影響顯著(P<0.05),對Sm影響不顯著。施氮量和溫度的交互作用對產量、Sm、PIabs、ABS/RC、TRo/RC、ETo/RC的影響均達到極顯著水平(P<0.01),對Fm、Fv/Fm、Area、PItotal、DIo/RC影響顯著(P<0.05),對Fo、Chla、Chlb和Chla+Chlb影響不顯著(表3)。
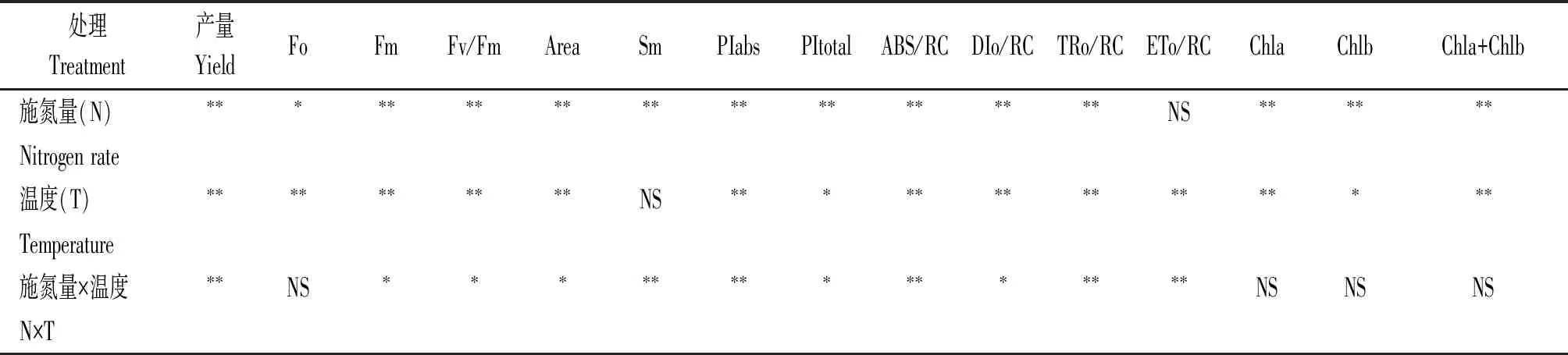
表3 果期高溫脅迫下不同施氮處理的黃瓜產量、葉綠素含量及熒光指標間的方差分析Tab.3 Variance analysis of yield,chlorophyll content and fluorescence indexes of cucumber under different nitrogen treatments under high temperature stress in fruit stage
3 結論與討論
本研究發現,相同溫度脅迫下施氮可以顯著增加黃瓜葉片的Chla、Chlb和Chla+Chlb,相同施氮水平下35 ℃輕度脅迫也可以顯著增加黃瓜葉片的葉綠素含量,這可能是因為溫度影響了葉綠素合成中的酶促反應,輕度高溫和氮肥的施用在一定程度上提高了植物葉片葉綠素合成酶的活性和氮代謝相關酶活性,從而促進葉綠素的合成,增加葉片葉綠素含量[14]。葉綠素熒光參數是反映植物葉片光化學活性的重要指標,可以更加具體的表現逆境下植物對光能的吸收利用與轉化程度[23-24]。本研究發現,果期高溫脅迫下合理施氮可以顯著降低黃瓜葉片的ABS/RC、TRo/RC和DIo/RC,提高ETo/RC、Fm、Fv/Fm、Plabs、Area和Sm。果期高溫脅迫導致了黃瓜葉片PSⅡ受體側受傷,Fm顯著減小,降低了電子傳遞能力,而氮的合理供應一方面可以顯著提高黃瓜葉片Fm和Sm,增大PSⅡ受體側電子受體庫的容量,使得傳遞到電子傳遞鏈的電子超過QA的電子受體的概率上升,從而提高Fv/Fm[25-26];另一方面氮的施用可以顯著降低黃瓜葉片DIo/RC,使被捕獲的光能更多地用于光化學反應,而不是用于QA的還原,進而利于電子受體QA后的電子傳遞,提高ETo/RC,減緩PSⅡ受體側損傷,促進PSⅡ的能量轉化和電子傳遞,本研究結果與Kalaji等[27]、朱榮等[28]、王木蘭等[29]、何海鋒等[30]的研究結果相類似。在受到高溫脅迫后一些黃瓜葉片的PItotal出現比室溫更高的情況,這可能與植物具有一定的抗逆性有關,即黃瓜在受到高溫脅迫后,會激發體內的抗氧化系統,對環境產生一定的抵抗能力[31]。
植物葉片生理的變化信息可以通過熒光參數反映到OJIP曲線上。本研究發現,果期高溫脅迫和不施氮處理的黃瓜葉片OJIP曲線中的I點上升緩慢且P點熒光強度降低,這是因為高溫脅迫和不施氮處理的黃瓜葉片PSⅡ反應中心出現失活,光電子從電子受體QA到QB的傳遞被抑制,PSⅡ和PSⅠ間電子分配不均、電子傳遞受阻導致的[32];施氮后各溫度的黃瓜葉片I點和P點的熒光強度得到提高,其中35,38 ℃脅迫下N3水平的I點上升較快,41 ℃脅迫下N2水平的I點上升較快,這說明合理施氮可以減輕電子傳遞被抑制狀況,促進光系統間電子傳遞均衡。將OJIP曲線標準化為相對可變熒光后發現對照溫度下各施氮水平的果期黃瓜葉片OEC均未失活(ΔWO-J<0)且PSⅡ中心類囊體能量傳遞順暢(ΔWO-K<0),而38,41 ℃脅迫下黃瓜葉片OEC出現失活現象(ΔWO-J>0),類囊體之間的能量連接性降低(ΔWO-K>0)。這可能是由于高溫脅迫下PSⅡ供體側流向反應中心的電子與從反應中心流向受體側的電子失衡導致的,電子流動不平衡會使得PSⅡ反應中心被過剩的電子氧化,破壞PSⅡ供體側結構的完整性和電子傳遞[33-34]。施氮后黃瓜葉片OEC活性增強,類囊體間的能量連接性得到提高,其中41 ℃脅迫下N2處理的OEC整體活性較強,類囊體間能量傳遞最順暢,38 ℃脅迫下N3處理的OEC整體活性和類囊體間能量連接性最強。可見,氮素的合理施用有利于提高植物葉片的OEC活性,促進PSⅡ中心類囊體間的能量傳遞,進而促進光合作用。此外,果期高溫脅迫下黃瓜葉片PSⅠ受體側末端電子受體庫被抑制(WO-I≥1部分I~P相振幅越小,抑制越強),施氮可以增大黃瓜葉片I~P相振幅,減緩電子受體庫的抑制狀況,其中N2處理效果最好,這可能是因為高溫導致PSⅠ產生光抑制,而氮的合理施用可以提高黃瓜葉片光系統的抗逆性。高溫脅迫加速了黃瓜葉片PSⅠ末端電子受體庫的還原速率,且溫度越高還原速度越快,試驗結果與徐超等[35]的研究結果相類似,但高溫脅迫下施氮卻降低了PSⅠ末端電子受體庫的還原速率,具體機理還有待進一步研究。
本試驗結果顯示,果期高溫脅迫顯著降低了各施氮處理的黃瓜產量,其中41 ℃高溫脅迫產量降低幅度最大(減產55.5%~71.2%),施氮后高溫脅迫處理的黃瓜產量顯著增加了71.1%~422.5%,且將產量數據與施氮量進行擬合后發現,黃瓜獲得最高產量時的施氮量隨著溫度的升高呈先上升后降低趨勢。高溫會減弱植物PSⅡ反應中心的原初光化學反應產生的去激發能力,導致植物的PSⅡ系統失活或傷害,進而降低產量,而不同高溫條件下適宜施氮可以提高作物葉片中光合碳同化的關鍵酶,有效緩解葉片的衰老,產生更多的光合產物,并且最大化地向果實轉移,從而增加黃瓜的產量[36]。本研究發現,施氮量和溫度對設施黃瓜葉片熒光特性和產量的影響存在交互效應,說明溫度對黃瓜葉片熒光特性和產量的影響因施氮處理的不同而有顯著差異,反之亦然。分析其原因,可能是因為高溫脅迫使黃瓜同化物向庫器官的分配受到阻礙,輸出時間縮短,輸出速率下降,產量降低,而適量氮肥處理能夠提高同化物轉運能力,促進高溫脅迫下黃瓜營養器官向生殖器官的氮素轉運,從而提高氮素對黃瓜果實的貢獻率[37-38]。
果期高溫脅迫顯著導致了黃瓜葉片光系統反應中心受損,而合理施氮可以有效增強PSⅡ供體側OEC活性和類囊體之間的能量連接性,減輕PSⅠ受體側末端電子受體庫的抑制狀況,緩解果期高溫脅迫對黃瓜光合器官的損傷。施氮量和溫度對黃瓜葉片熒光特性和產量存在顯著的交互效應,黃瓜獲得最高產量時的施氮量隨著溫度的升高呈先上升后降低趨勢,即施氮量為236,283,177 kg/hm2時可分別有效提高35,38,41 ℃的高溫脅迫下黃瓜葉片光合作用并獲得最高產量。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
