2-十三烷酮脅迫下棉鈴蟲FoxAl調控CYP6B6的表達
2022-06-10 03:25:38魏倩劉小寧趙潔
生物技術通報 2022年5期
魏倩 劉小寧 趙潔
(1. 新疆大學生命科學與技術學院,烏魯木齊 830046;2. 石河子大學農學院,石河子 832003)
叉頭框蛋白(forkhead box protein,Fox)家族是一類含有保守的fork head(FH)結構域的轉錄因子,該結構域中的“螺旋-轉角-螺旋”基序與下游基因啟動子區的順式作用元件結合[1-2]。Fox轉錄因子的分布十分廣泛,從酵母、線蟲、昆蟲、海鞘、爪蟾、小鼠到人類中,已經發現了數百種Fox蛋白[3-5]。根據Fox蛋白在N端和C端的變異程度,將Fox家族分為24個亞家族和1個孤兒類別[6]。已發表的昆蟲基因組或轉錄組結果表明,昆蟲體內并不包含所有的Fox亞家族,例如家蠶體內主要缺失FoxAB、FoxE、FoxH、FoxI、FoxK、FoxL1、FoxM、FoxQ1、FoxR和FoxS[7]。目前,昆蟲Fox蛋白的研究主要集中于FoxO和FoxA亞家族,它們在昆蟲的氧化應激、生長發育、細胞凋亡、解毒代謝、糖脂代謝和先天免疫等方面發揮著重要的作用[8-12]。
棉鈴蟲(Helicoverpa armigera)幼蟲是一種多食性的農業重要防治害蟲。目前,關于棉鈴蟲Fox蛋白的研究報道極少。Bao等[13]從棉鈴蟲的蛹內克隆獲得第一個Fox基因,即HaFoxA,并發現FoxA蛋白可以與滯育激素和信息素生物合成激活神經肽(diapause hormone-pheromone biosynthesis-activating neuropeptide,DH-PBAN)基因的啟動子結合,從而參與蛹的激素合成并影響滯育過程。Cai等[14]首次克隆獲得棉鈴蟲的FoxO基因,并發現在20-羥基蛻皮激素的影響下,FoxO蛋白與Broad轉錄因子亞型7(broad isoform 7,BrZ7)基因的啟動子結合,提高BrZ7以及下游羧肽酶A(carboxypeptidase A,CPA)基因的表達量,從而在棉鈴蟲的蛻皮過程中促進蛋白質水解。Li等[15]將棉鈴蟲3齡幼蟲體內的FoxA進行RNA干擾(RNA interference,RNAi)后發現,響應蘇云金芽孢桿菌Bacillus thuringiensis的兩個Cry毒素蛋白受體,即ATP結合盒轉運蛋白(ATP-binding cassette transporter proteins,ABCC2和ABCC3)基因的表達都顯著降低,并且幼蟲對Cry毒素的耐受性明顯增強。
2-十三烷酮(2-tridecanone,2-TD)是一種天然存在于茄科植物中的重要次生物質,它能夠誘導昆蟲體內細胞色素P450解毒酶(cytochrome P450 enzymes)基因的過量表達,例如低濃度的2-TD能顯著提高棉鈴蟲CYP3、CYP6亞家族基因的表達,激活棉鈴蟲的解毒代謝通路[16-17]。CYP6B6是棉鈴蟲中腸內一個重要的解毒酶基因,它在棉鈴蟲的有毒物質代謝和生長發育過程中都發揮重要作用[18]。然而,2-TD長期脅迫卻會降低CYP6B6表達量,導致棉鈴蟲化蛹時間延長、并顯著降低化蛹率和羽化率[19-20]。同時,在低濃度2-TD短期脅迫后,對CYP6B6基因的啟動子進行分析發現,-373/+21 bp處(即HE1片段)是最重要的活性序列[21]。本課題組前期利用酵母單雜交技術,篩選2-TD短期脅迫后能與HE1片段結合的調控因子,發現一個陽性酵母的測序片段中含有FH結構域的部分序列;同時在2-TD短期處理6齡幼蟲的中腸轉錄組中發現一個新的Fox基因,它和CYP6B6一樣都能響應低濃度2-TD脅迫。將克隆得到的新Fox基因與已發表的昆蟲FoxA進行多序列比對后發現,該基因包含Fox家族保守的FH結構域,以及一個疑似FoxA亞家族轉錄激活結構域(Domain Ⅱ)的相關區域,并把該基因命名為FoxA類似基因(forkhead box protein A-like,FoxAl)[22]。隨后,對CYP6B6啟動子進行轉錄因子結合位點預測,發現HE1片段上含有多個Fox轉錄因子的結合位點。基于以上的研究基礎,我們推測FoxAl有可能是CYP6B6的轉錄因子,從而參與棉鈴蟲的解毒代謝和生長發育過程。
因此,本研究先利用酵母自激活試驗,驗證FoxAl蛋白是否具有轉錄激活功能;隨后通過凝膠阻滯試驗,驗證FoxAl蛋白能否與CYP6B6啟動子HE1片段結合。在確定FoxAl是CYP6B6的轉錄因子后,一方面用不同濃度的2-TD脅迫處理棉鈴蟲6齡幼蟲,另一方面注射FoxAl dsRNA于棉鈴蟲5齡幼蟲體內,通過實時熒光定量PCR(Real-time quantitative PCR,qPCR)技術測定不同時間后幼蟲中腸內FoxAl和CYP6B6的表達情況,確定FoxAl與CYP6B6啟動子的調控關系,為FoxAl蛋白參與棉鈴蟲的生長發育和解毒代謝過程提供依據,為進一步探索棉鈴蟲對次生物質的適應機制奠定基礎。
1 材料與方法
1.1 材料
供試昆蟲:棉鈴蟲采集于烏魯木齊市安寧渠鎮,并在本實驗室用人工飼料長期飼養,試蟲種群為第5代。人工飼料配方:玉米粉120 g,大豆粉40 g,瓊脂20 g,山梨酸1 g,酵母粉12 g,青霉素0.1 g,抗壞血酸2 g,維生素C 4 g,肌醇0.2 g,復合維生素B 0.9 g,蒸餾水620 mL。飼養條件:溫度(26±1)℃,相對濕度(75±5)%,光周期16L∶8D。
菌株和質粒:釀酒酵母Saccharomyces cerevisiae Y2HGold菌株購自Clontech公司,最適生長溫度30℃;大腸桿菌Escherichia coli DH5α感受態細胞購自北京全式金生物技術有限公司,最適生長溫度37℃;大腸桿菌Transetta(pET32a-FoxAl)菌株由課題組前期構建,最適生長溫度37℃,蛋白表達最適溫度25℃。pMD18-T-FoxAl質粒由課題組前期構建,pGBKT7質粒購自Clontech公司。
試 劑:DIG-High Primer DNA Labeling and Detection Starter Kit II購自Roche公司;Matchmaker Insert Check PCR Mix 2、2×YPDA Broth、SD/-Trp with Agar和X-α-Gal均購自Clontech公司;EMSA/Gel-Shift結合緩沖液(5×)、EMSA/Gel-Shift上樣緩沖液(無色,10×)和EMSA/Gel-Shift上樣緩沖液(藍色,10×)均購自上海碧云天生物技術;2-十三烷酮購自上海麥克林生化科技有限公司;DL2000 DNA Marker、DL5000 DNA Marker、BamH I、EcoR I和T4 DNA Ligase均購自寶生物工程(大連)有限公 司;Proteinlso Ni-NTA Resin、EasyPure RNA Kit、EasyScript First-Strand cDNA Synthesis SuperMix和PerfectStart Green qPCR SurperMix(+Dye I)均購自北京全式金生物技術有限公司;T7 RiboMAX Express RNAi System購自北京普洛麥格生物技術有限公司;其他化學試劑均為國產分析純試劑。
1.2 方法
1.2.1 FoxAl蛋白的轉錄激活活性測定 以前期測序正確的pMD18-T-FoxAl質粒為模板,PCR擴增獲得FoxAl的開放閱讀框(ORF)序列,其中上、下游引物分別為(5'-GGAATTCATGGCGGCTAGAACA GGT-3',EcoR I酶切位點)和(5'-CGGGATCCCTAGT CCACCGTGATGA-3',BamH I酶切位點)。隨后,用限制性內切酶EcoR I和BamH I將FoxAl的ORF和pGBKT7質粒分別進行雙酶切,酶切產物經T4 DNA Ligase過夜連接后,轉化大腸桿菌DH5α,再通過菌液PCR和酶切鑒定獲得重組后的pGBKT7-FoxAl質粒。按照Y2HGold菌株使用說明,將pGBKT7-FoxAl質粒轉入酵母Y2HGold菌株內,并在SD/-Trp固體培養基上30℃倒置培養3-4 d,挑取生長良好、直徑約2-3 mm的酵母菌落進行PCR鑒定。取適量鑒定正確的Y2HGold(pGBKT7-FoxAl)重組菌株,以1∶100的比例稀釋于0.9% NaCl溶液中,取100 μL重懸菌液分別涂布于SD/-Trp和SD/-Trp/X-α-Gal固體培養基上,30℃倒置培養2-3 d,觀察菌落生長狀況和顯色情況。陰性對照為Y2HGold和Y2HGold(pGBKT7)菌株按同等比例稀釋后,生長于SD/-Trp/X-α-Gal固體培養基上。
1.2.2 FoxAl蛋白和CYP6B6啟動子的結合反應 按照趙潔等[22]的文獻,經表達、純化和超濾獲得融合蛋白His-FoxAl。按照DIG-High Primer DNA Labeling and Detection Starter Kit II的說明書制備CYP6B6啟動子HE1片段的核酸探針,并檢測探針的標記效率。按照EMSA/Gel-Shift結合緩沖液的步驟進行HE1探針(1 ng)和FoxAl蛋白(1 μg或1.5 μg)的結合反應,參考Michael R. Green的《Molecular Cloning:A Laboratory Manual》第二章方案3制備5%的非變性聚丙烯酰胺凝膠,并進行DNA-蛋白質復合物的核酸電泳。利用浸沒式轉印電泳儀將HE1探針轉移至帶有正電荷的尼龍膜上,并進行紫外線交聯固定。按 照DIG-High Primer DNA Labeling and Detection Starter kit II的步驟進行HE1探針的免疫印跡和化學發光成像。此外,為了避免假陽性的阻滯結果,進行未標記探針和無關基因片段的冷競爭反應,分別在結合反應體系中再加入標記探針100倍量的未標記HE1片段或棉鈴蟲肽聚糖識別蛋白(Peptidoglycan recognition proteins,PGRP-B)基因片段。
1.2.3 FoxAl-dsRNA處理棉鈴蟲5齡幼蟲 以棉鈴蟲FoxAl的ORF為模板,經PCR擴增和產物回收獲得含有T7啟動子的目的片段,按照T7 RiboMAX Express RNAi System試劑盒的操作步驟合成和純化獲得FoxAl的dsRNA。取當天蛻皮的5齡幼蟲饑餓2 h后冰上麻醉20 min,在幼蟲腹部第6節和第7節的節間膜處,用微量注射器將FoxAl 的dsRNA注入幼蟲體腔內,dsRNA為5 μg/頭,對照組注射等量的Nuclease-free water。注射結束后,將棉鈴蟲幼蟲置于35 mm的培養皿內,在幼蟲恢復正常活動后放置于含有人工飼料的玻璃管內,使其在正常環境條件下生長。
1.2.4 2-TD脅迫處理棉鈴蟲6齡幼蟲 將所需2-TD溶于60℃預熱的10%乙醇溶液中,充分融化后添加于等質量的棉鈴蟲人工飼料中,分別配制成不同劑量(0,5,10和20 mg/g)的處理飼料。取當天蛻皮的6齡幼蟲饑餓2 h后,放置于含有不同2-TD劑量飼料的玻璃管內,使其在正常環境條件下生長。
1.2.5 qPCR檢測FoxAl和CYP6B6的表達譜 在dsRNA注射后的不同時間(24,48,72和 96 h)和2-TD處理后的不同時間(6,12,20,30和48 h),取3頭仍然存活的幼蟲的中腸組織作為一個生物學樣本,按照EasyPure RNA Kit和EasyScript First-Strand cDNA Synthesis SuperMix的操作步驟提取中腸的總RNA,并反轉錄為相對應的cDNA,每個處理包含3個生物學樣本。按照PerfectStart Green qPCR SurperMix(+Dye I)的說明書,以棉鈴蟲Tubulin作為內參基因,經qPCR技術分別檢測FoxAl和CYP6B6的相對表達情況,每個生物學樣本包含3個技術重復。qPCR反應體系為10.0 μL的2×SYBR Green PCR master mix、8.0 μL的RNase-free H2O、0.5 μL的上游引物、0.5 μL的下游引物、1.0 μL的cDNA。反應 程 序:94℃ 2 min;94℃ 30 s,58℃ 30s,40個循環。FoxAl引物F:5'-TGATGCCGTTGAGAAGACTA-3';R:5'-CACTTTCCCTGACCACTTG-3'。Tubulin引 物F:5'-TCCAACTCACACACTCGCT-3';R:5'-GGAAGCAGATGTCGTATAATG-3'。CYP6B6引物F:5'-TTCAAACTTATACCATGTCCACAA-3';R:5'-CCAATTGACGGAGCTCTAGAATCA-3'。
1.2.6 數據分析 在每個處理時間點,RNAi試驗中以Nuclease-free water組為對照,2-TD脅迫試驗中以0 mg/g組為對照,用2-ΔΔCt法計算FoxAl和CYP6B6的相對表達量。運用Prism 8.0軟件進行統計分析:RNAi試驗,在每個時間點進行t檢驗(Student’s t test);2-TD脅迫試驗,在每個時間點和每個濃度下都進行單因素方差分析(one-way ANOVA)和多重比較(Tukey法)。運用SPSS 20.0軟件,將2-TD脅迫試驗中FoxAl和CYP6B6的相對表達量,按照不同脅迫濃度和不同脅迫時間進行相關性分析。
2 結果
2.1 棉鈴蟲FoxAl的轉錄激活功能驗證
將棉鈴蟲FoxAl的ORF連接至酵母表達載體pGBKT7質粒,構建的重組質粒pGBKT7-FoxAl經EcoR I和BamH I酶切,圖1-A電泳結果顯示酶切片段小于1 000 bp、大于500 bp,其位置與FoxAl ORF(669 bp)的大小一致,表明重組質粒pGBKT7-FoxAl構建成功。將重組質粒轉入酵母Y2HGold菌株感受態后,進行酵母菌落PCR,圖1-B結果顯示擴增條帶略小于750 bp,其位置與FoxAl ORF的大小一致,表明酵母菌株Y2HGold(pGBKT7-FoxAl)構建成功。隨后,將構建好的Y2HGold(pGBKT7-FoxAl)菌液涂布于SD/-Trp和SD/-Trp/X-α-ga1培養基上,通過顏色變化檢測FoxAl蛋白對酵母MEL1報告基因是否有激活作用。圖1-C結果顯示,將兩個對照Y2HGold和Y2HGold(pGBKT7)的菌液直接涂布于SD/-Trp/X-α-ga1培養基上,前者不能生長、后者正常生長但無顏色變化,對照組的結果符合預期,表明整個轉化過程正常且無污染;將Y2HGold(pGBKT7-HaFoxAl)菌液涂布于SD/-Trp和SD/-Trp/X-α-ga1培養基上,酵母菌落正常生長且大小合適,表明插入基因能正常表達,同時該菌液在X-αga1誘導下呈現藍色,表明FoxAl蛋白本身含有轉錄激活結構域,能激活酵母體內下游基因的正常表達。
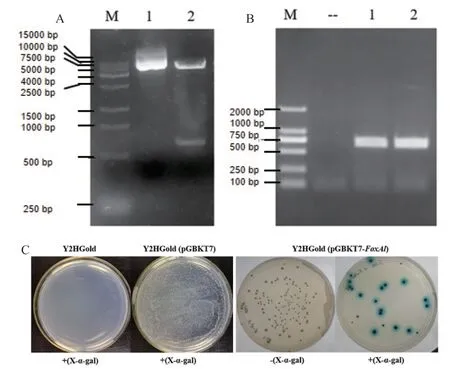
圖1 酵母Y2HGold(pGBKT7-FoxAl)菌株的轉錄激活功能檢測Fig.1 Transcriptional activation test of transformed yeast Y2HGold(pGBKT7-FoxAl)strain
2.2 棉鈴蟲FoxAl與CYP6B6啟動子HE1片段結合
首先,用地高辛標記的HE1探針與有活性的His-FoxAl蛋白進行凝膠遷移率檢測,圖2-A的結果顯示,1 μg和1.5 μg的His-FoxAl蛋白都能與1 ng HE1探針結合并形成阻滯條帶。隨后,通過無關基因片段和未標記探針的冷競爭反應,進一步驗證1 ng HE1探針與1 μg His-FoxAl蛋白結合形成阻滯條帶的真實性。圖2-B的結果顯示,泳道3在反應體系中再加入標記探針100倍量的棉鈴蟲PGRP-B片段時,HE1探針與FoxAl蛋白形成的阻滯條帶仍然存在;而泳道4中再加入100倍量的未標記HE1片段時,原有的HE1-FoxAl阻滯條帶消失不見;這兩個競爭反應結果表明,FoxAl蛋白與CYP6B6啟動子HE1片段的結合是特異性的,是通過FoxAl蛋白的識別螺旋與HE1片段的特定堿基以氫鍵形式結合。

圖2 棉鈴蟲FoxAl蛋白與CYP6B6啟動子HE1片段的結合驗證Fig. 2 Verification of FoxAl in H. armigera binding to the HE1 fragment of CYP6B6 promotor
2.3 棉鈴蟲FoxAl沉默對CYP6B6表達水平的影響
將5 μg的FoxAl dsRNA注射到棉鈴蟲5齡幼蟲體內,在注射后的24、48、72和96 h,檢測中腸內FoxAl和CYP6B6的表達量變化情況。圖3-A的FoxAl相對表達量結果顯示:與對照組相比,注射FoxAl dsRNA后中腸內的FoxAl表達量從48 h開始顯著降低(P=0.001 8),并在96 h降至最小值(P=0.000 6);圖3-B的CYP6B6相對表達量結果顯示:與對照相比,FoxAl dsRNA處理后CYP6B6的表達從24 h開始顯著降低(P=0.001 9),在48 h降至最小值(P<0.000 1),并在96 h內一直被顯著降低(P=0.008 2)。

圖3 FoxAl dsRNA 注射后棉鈴蟲5齡幼蟲中腸內FoxAl和CYP6B6的相對表達量Fig. 3 Relative expression of FoxAl and CYP6B6 in the midgut of 5th instar larvae after FoxAl dsRNA injected
2.4 2-TD處理對棉鈴蟲中腸FoxAl和CYP6B6表達水平的影響
用3種濃度的2-十三烷酮(2-TD)處理棉鈴蟲六齡幼蟲,檢測中腸內FoxAl和CYP6B6的表達量變化情況。圖4-A的FoxAl相對表達量結果顯示:在5 mg/g的2-TD處理組內(F=5.180,P=0.001 9),12 h的表達量最高、30 h的表達量最低,且30 h時顯著低于空白組(P=0.003 5);在10 mg/g的2-TD濃度下(F=12.86,P<0.000 1),12 h的表達量最高、48 h的表達量最低,6 h時極明顯高于對照(P<0.000 1), 20,30和48 h時 顯 著 低 于 對 照(P=0.027 5,P=0.001 4,P=0.042 7);在20 mg/g處理中(F=11.27,P<0.000 1),12 h的表達量最高、48 h的表達量最低,12 h時顯著高于空白對照(P=0.013 1)、而20和48 h時明顯低于空白對照(P=0.006 3,P=0.027 3)。在3個2-TD濃度處理組內,棉鈴蟲FoxAl表達量隨時間變化基本一致,都是短時間內表達量上升、至12 h達到最高值,然后在20 h表達量快速下降。
CYP6B6的相對表達量結果顯示:在5 mg/g的2-TD處理后(F=5.016,P=0.002 3),20 h的表達量最高、30 h的表達量最低,且20 h時明顯高于對照(P=0.023 6);在10 mg/g 2-TD處理組(F=33.63,P<0.000 1),12 h的表達量最高、48 h的表達量最低,12 h時顯著高于對照(P=0.037 3),30和48 h時顯著低于對照(P=0.043 1,P=0.023 4);在20 mg/g的處理中(F=12.95,P<0.000 1),12 h的表達量最高、48 h的表達量最低,12 h時極顯著高于空白對照(P=0.000 8)。在3個2-TD濃度處理后,CYP6B6表達量隨時間變化基本相似,都是在12 h內急劇升高、在20 h內基本維持高水平表達,然后在30 h迅速下降(圖4-B)。
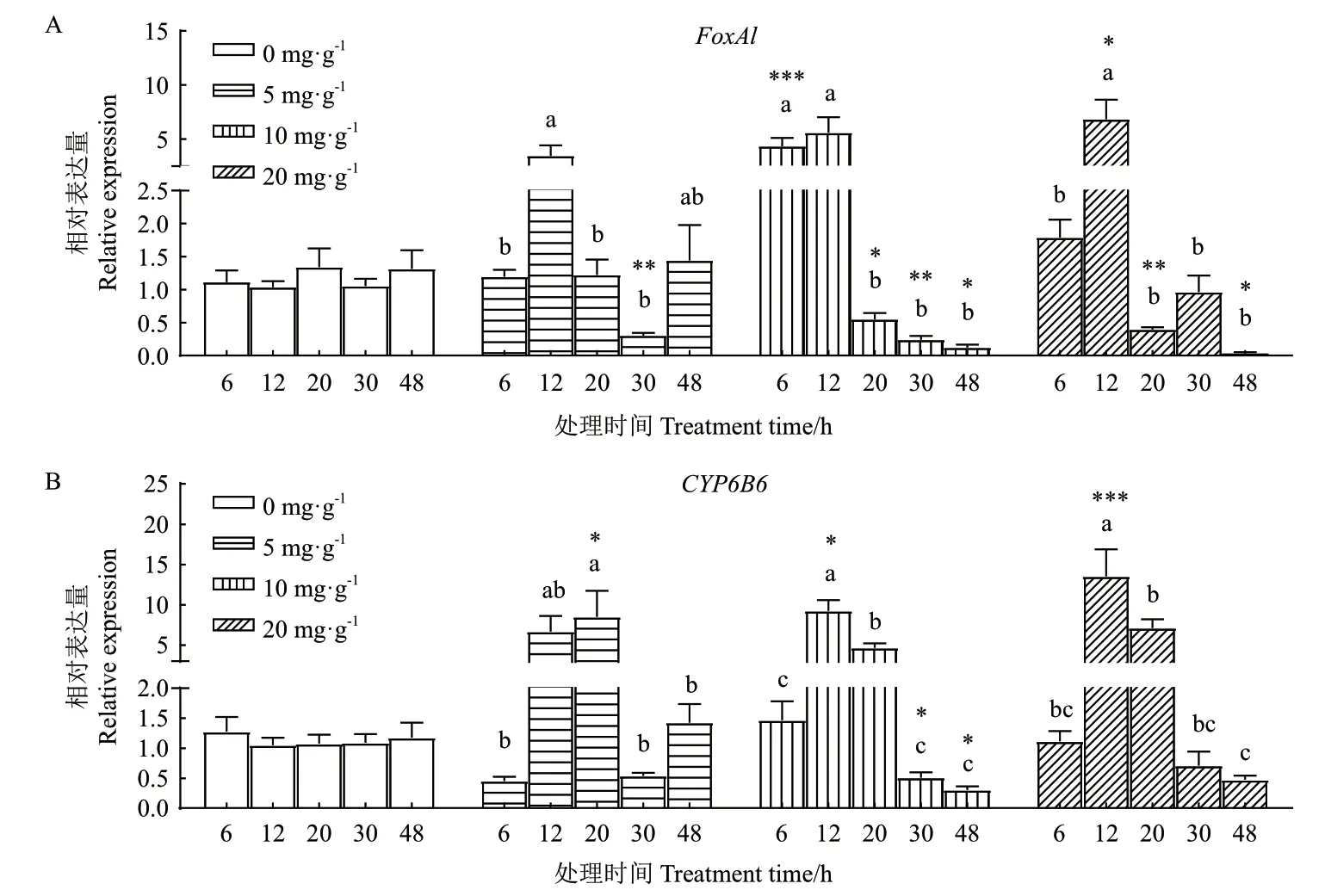
圖4 2-TD脅迫后棉鈴蟲6齡幼蟲中腸內FoxAl和CYP6B6的相對表達量Fig. 4 Relative expression of FoxAl and CYP6B6 in the midgut of the 6th instar larvae after 2-TD treated
將不同2-TD濃度處理后的FoxAl和CYP6B6的相對表達量進行相關性分析。表1結果顯示,3個脅迫濃度下FoxAl和CYP6B6的表達量都是正相關的,兩者的相關系數隨著2-TD濃度增加逐漸增大,并且在20 mg/g濃度處理時兩者呈顯著地高度正相關(r=0.819,P=0.045);除了20 h以外,其他脅迫時間點下FoxAl和CYP6B6的表達量都是正相關的,12 h、30 h和48 h時兩者的表達量是高度正相關(r=0.987,r=0.863,r=0.978),其中12 h和48 h時都是顯著相關(P=0.007,P=0.011)。

表1 FoxAl和CYP6B6表達量的相關性分析Table 1 Correlation analysis of FoxAl and CYP6B6 expression
3 討論
課題組前期分析棉鈴蟲FoxAl蛋白的氨基酸序列時,發現該蛋白只含有一個完整的FH結構域,不包括FoxA亞家族的兩個轉錄激活域[22]。為了明確FoxAl蛋白是否具有轉錄激活活性,本研究通過酵母自激活試驗證明了該蛋白能夠激活MEL1報告基因的轉錄,從而使酵母菌株在缺陷培養基上正常生長并顯藍色。作為棉鈴蟲體內一個新的Fox家族基因,FoxAl的具體功能與蛋白結構密切相關,因此后期還將進一步明確FoxAl蛋白中主要的轉錄激活區域以及相關位點。
已有研究表明,在外界物質的刺激下,昆蟲Fox蛋白的表達量會迅速上升并進入細胞核內,調控下游基因的轉錄[14]。例如用一種大環內酯類免疫抑制劑—雷帕霉素飼喂果蠅Drosophila成蟲,其腸道內FoxA的表達量升高并大量入核,FoxA激活Diptericin和Metchnikowin基因的轉錄,表達兩種腸道抗菌肽,從而參與果蠅免疫系統[10]。本研究在植物次生物質的脅迫下,棉鈴蟲體內FoxAl的表達量變化情況。結果顯示3種低濃度(5,10和20 mg/g)的2-TD脅迫6齡幼蟲后,中腸內FoxAl的表達量在12 h內迅速升高,并在20 h迅速降低,隨著脅迫時間的延長,該基因mRNA含量基本維持穩定。目前棉鈴蟲FoxAl的抗體正在制備中,后期將從蛋白質水平進一步證實FoxAl的表達情況。
細胞色素P450是昆蟲體內一種重要的解毒酶系。大量研究表明,P450基因過量表達是昆蟲產生抗藥性的機制之一,其中CYP6亞家族與植物次生物質和殺蟲劑的耐受性密切相關[23-24]。例如2-TD能誘導棉鈴蟲體內CYP6B7基因的過量表達,并使棉鈴蟲對擬除蟲菊酯產生抗性[25-26]。根據已發表的文章,多次證實低濃度(<50 mg/g)的2-TD短期脅迫棉鈴蟲幼蟲后,中腸組織內CYP6B6的蛋白含量能迅速增加,并參與解毒代謝過程[19-21]。本研究利用低濃度的2-TD處理棉鈴蟲6齡幼蟲不同時間后,檢測中腸內CYP6B6的表達量,本試驗結果與前人的研究結果相似,3個2-TD濃度都能快速提高CYP6B6的mRNA含量,而隨著脅迫時間延長,CYP6B6的表達量都會逐漸降低。在處理30 h-48 h的時間段內,與空白對照組相比,5和10 mg/g處理組的幼蟲取食量減少,而20 mg/g處理組的幼蟲大量提前預蛹,其中部分幼蟲的中腸組織縮短、顏色加深,甚至呈現粉紅色。這些現象表明,即使是低濃度的2-TD,長時間脅迫也會降低幼蟲的解毒能力,甚至影響棉鈴蟲的生長發育。
在預蛹的果蠅轉化品系P[hs-Fkh111]中,37℃熱處理增加唾腺中FoxA的含量后,發現下游P450基因的表達發生改變,其中CYP9B2,CYP6A20和CYP28D1的表達量分別提高3.88,24.39和100.87倍,而CYP4E2和CYP6V1的表達量變化倍數都為-2.15[27]。同時在前期進行CYP6B6啟動子分析時,預測出大量的Fox蛋白結合位點。因此,我們推測新的棉鈴蟲FoxAl蛋白也許能與CYP6B6啟動子結合,從而響應植物次生物質脅迫。本研究通過凝膠阻滯試驗證實了該假設,棉鈴蟲FoxAl蛋白能與CYP6B6啟動子的核心序列HE1片段結合。通過多種軟件的進一步篩選,HE1片段中可能與FoxAl蛋白結合的位點有3個,后期將通過截斷HE1片段,進一步縮小FoxAl蛋白的結合范圍,同時合成相應的突變探針,明確FoxAl蛋白的具體結合位點。
本研究通過腹腔注射法沉默幼蟲中腸內FoxAl的表達后,CYP6B6的表達量也發生顯著降低,這說明FoxAl蛋白對CYP6B6的轉錄調控是正向的。此外,2-TD脅迫后FoxAl和CYP6B6的表達量變化趨勢基本相似,對兩者的表達量進行相關性分析,結果顯示二者總體是正相關的,甚至在20 mg/g處理濃度、以及12,30和48 h處理時間都是高度正相關,該結果表明FoxAl可能是CYP6B6的轉錄激活因子,正向調控CYP6B6的表達。值得注意的是,在處理20 h后FoxAl的表達量顯著降低,而CYP6B6依然高水平地表達,兩者的表達量呈低度負相關。轉錄因子FoxAl表達量出現拐點的時間(12 h)早于下游基因CYP6B6出現的時間(20 h),在此時間段內可能還有其它轉錄激活因子或輔因子,使CYP6B6持續過量表達。同時,因為檢測點相隔時間較長,后期將進一步縮短間隔時間,細化FoxAl和CYP6B6的表達量變化情況。
4 結論
棉鈴蟲FoxAl蛋白能與CYP6B6啟動子結合,并且在有毒物質脅迫下,FoxAl蛋白可能是CYP6B6的轉錄激活因子。