大田野生麥苗穗發(fā)芽抗性的鑒定與遺傳分析
2022-08-19 09:22:12張希太謝淑芹藺桂芬
農(nóng)學學報 2022年8期
關(guān)鍵詞:生長
張希太,肖 磊,董 策,謝淑芹,藺桂芬
(邯鄲市農(nóng)業(yè)科學院生物技術(shù)研究室,河北邯鄲 056001)
0 引言
小麥成熟期穗發(fā)芽(pre-harvest sprouting,PHS)是一種世界性的氣候災害。聯(lián)合國糧農(nóng)組織統(tǒng)計,每年全球因穗發(fā)芽造成的損失占世界小麥總產(chǎn)量的20%,約相當于中國小麥年產(chǎn)量[1]。中國有85%的小麥面積都存在穗發(fā)芽隱患[2],黃淮麥區(qū)、長江中下游麥區(qū)曾多次大面積發(fā)生小麥穗發(fā)芽災害,西南冬麥區(qū)及東北春麥區(qū)穗發(fā)芽災害也頻繁發(fā)生[3]。小麥穗發(fā)芽后不僅產(chǎn)量降低,更主要是造成品質(zhì)嚴重變劣,甚至喪失食用、加工利用的價值。穗發(fā)芽災害不發(fā)則已,一發(fā)則損失慘重。穗發(fā)芽災害發(fā)生的外因是連續(xù)陰雨和潮濕的氣候,內(nèi)因是小麥品種本身對穗發(fā)芽的抗性差。當今科技影響氣候的能力有限,因此,只能通過抗穗發(fā)芽育種提高小麥品種自身的穗發(fā)芽抗性來抵抗災害的發(fā)生。小麥抗穗發(fā)芽育種存在著2 個技術(shù)瓶頸,首先是用于育種的高抗穗發(fā)芽種質(zhì)資源嚴重匱乏;其次是在非小麥穗發(fā)芽災害發(fā)生年份,育種后代材料抗穗發(fā)芽性狀的大田定向選擇困難。高抗穗發(fā)芽種質(zhì)資源的嚴重匱乏是限制小麥抗穗發(fā)芽育種未能取得突破性進展的主要障礙。因此創(chuàng)制或收集高抗穗發(fā)芽的小麥種質(zhì)資源是當前小麥抗穗發(fā)芽育種的首要問題。
小麥抗穗發(fā)芽研究一直受到世界各國的重視。中國科技工作者在小麥抗穗發(fā)芽育種方面做了大量工作。首先對現(xiàn)有的大量小麥品種資源材料進行了抗穗發(fā)芽鑒定篩選。也選出了一些抗性相對較好的材料[3-8],但是受現(xiàn)有小麥遺傳背景的限制并沒有從根本上找到解決穗發(fā)芽問題的抗源基因。
2011 年,國家小麥工程技術(shù)研究中心,利用基因槍法獲得了轉(zhuǎn)反義硫氧還蛋白基因(anti-trxs)的抗穗發(fā)芽轉(zhuǎn)基因小麥材料。通過人工模擬降雨穗發(fā)芽試驗,轉(zhuǎn)anti-trxs基因小麥種子比非轉(zhuǎn)基因?qū)φ招←湻N子的萌動時間明顯延長,穗發(fā)芽受到明顯的抑制[9-10]。2013年,美國農(nóng)業(yè)部農(nóng)業(yè)研究局和堪薩斯州立大學的研究人員發(fā)現(xiàn)并克隆出一個被命名為PHS 的能防止植物提前發(fā)芽的基因[11]。但是這些材料轉(zhuǎn)入的是人工克隆的工程基因,對生物環(huán)境的安全性還需評估,特別是目前國家嚴格限制在糧食作物上轉(zhuǎn)工程基因的形勢下,該材料在小麥抗穗發(fā)芽育種上的應用還需要等待。
通過觀察人們發(fā)現(xiàn)小麥的伴生雜草‘節(jié)節(jié)麥’具有極強的抗穗發(fā)芽特性。四川農(nóng)業(yè)大學利用遠緣雜交技術(shù)帥先將‘節(jié)節(jié)麥’的高抗穗發(fā)芽基因引入小麥,創(chuàng)制出了小麥高抗穗發(fā)芽材料RSP[12-15],為小麥抗穗發(fā)芽育種提供了寶貴的種質(zhì)資源;河南大學在‘節(jié)節(jié)麥’抗穗發(fā)芽基因研究與利用方面也做了大量工作[16-18]。
在“一年兩熟”種植模式的黃淮麥區(qū),一般當年5—6 月小麥收獲后種植大秋作物(以玉米為主),9 月底10 月初大秋作物收獲后再播種冬小麥。秋收后耕地前,大田中往往生長有少量的剛剛出土不久的野生麥苗,這些野生麥苗是小麥收獲時由于粗放操作遺落于大田中的麥粒、麥穗萌發(fā)長成。這些遺落于大田中的麥粒、麥穗在土壤中經(jīng)過了夏季充沛的降雨直到9月底10月初秋收后才萌發(fā)出土,應該具有很強的休眠性和抗穗發(fā)芽能力,于是筆者收集了大量的野生麥苗,進行集中移栽,對其遺傳穩(wěn)定性、穗發(fā)芽抗性、遺傳親緣關(guān)系進行了研究。旨在獲取具有極強穗發(fā)芽抗性的種質(zhì)資源。
1 材料與方法
1.1 野生麥苗的收集移栽與遺傳穩(wěn)定性觀察
1.1.1 野生麥苗的收集與集中移栽 每年9 月底10 月初,秋收后耕地前,對計劃收集野生麥苗的農(nóng)田(夏茬為冬小麥)進行普查,將發(fā)現(xiàn)的野生麥苗進行根部帶土團移栽。大田中生長的野生麥苗分2 種類型:一類是單棵生長的野生麥苗(由落入大田的一粒種子萌發(fā)長成,圖1B),第二類是成簇生長的野生麥苗(由落入土中的整個麥穗上的種子萌發(fā)生長而來,圖1A)。第一類野生麥苗移栽入觀察圃時,對單株進行編號,記錄發(fā)現(xiàn)地塊及移栽時間等;第二類將簇分成單株移栽入觀察圃,編簇號和株號,記錄發(fā)現(xiàn)地塊及移栽時間等。本試驗野生麥苗的收集地為河北省成安縣商城鎮(zhèn)趙橫城村北農(nóng)田。分別于Ⅰ:2014年9月30日。Ⅱ:2015年9月30 日。Ⅲ:2016 年9 月30 日。Ⅳ:2017 年9 月30日。分4批次收集大田野生麥苗集中移栽到本研究室試驗田的觀察圃中。

圖1 秋收后冬小麥播種耕地前大田中出土的野生麥苗
1.1.2 集中移栽的野生麥苗生長一致性觀察與種子收獲 野生麥苗集中移栽入觀察圃后,立即澆水保證移栽苗的成活,加強管理,使移栽成活后的野生麥苗生長茁壯。從第一生育周期開始對第二類同一簇(穗)系內(nèi)的野生麥苗單株進行一致性測試,參照中華人民共和國國家標準(GB/T 19557.2—2004)[19]進行。小麥成熟后,對于第一類野生麥苗,按株號進行單株收獲并按株號保存種子。對于第二類野生麥苗,將生長一致、性狀穩(wěn)定的簇(穗)系內(nèi)所有單株混收并按簇(穗)系號保存種子;對個別遺傳性狀不穩(wěn)定、生長不一致的簇(穗)系進行淘汰或另行試驗,本文不再闡述。
1.1.3 野生麥苗的遺傳穩(wěn)定性觀察 第二個生育周期播種時,將第一類野生麥苗按株系播種;保留的第二類野生麥苗按不同的簇(穗)系播種,每系不播種少于2000株。整個生育期參照GB/T 19557.2—2004 標準觀察兩類野生麥苗各株系和簇(穗)系內(nèi)植株性狀的一致性,繼續(xù)對個別遺傳性狀不穩(wěn)定、生長不一致的株系與簇(穗)系進行淘汰。
第3個生育周期只對第一類野生麥苗株系內(nèi)植株進行生長一致性觀察。兩類野生麥苗的后代,都需進行2年的生長一致性觀察,如果連續(xù)2年株系或簇(穗)系內(nèi)植株的遺傳性狀都表現(xiàn)一致,說明該株系或簇(穗)系已經(jīng)遺傳穩(wěn)定。然后對遺傳穩(wěn)定的野生麥苗系進行穗發(fā)芽抗性鑒定。
1.2 遺傳穩(wěn)定野生麥苗系農(nóng)藝性狀調(diào)查與穗發(fā)芽抗性測定及種質(zhì)系的選擇
1.2.1 遺傳穩(wěn)定野生麥苗系主要農(nóng)藝性狀調(diào)查與相對穗發(fā)芽指數(shù)測定 將遺傳穩(wěn)定的野生麥苗系重新統(tǒng)一編號,按行距20 cm,株距10 cm單穴單粒播種,每系播種數(shù)量不少于500株。整個生育期調(diào)查株高、抗倒性、抗病性(主要調(diào)查白粉病、銹病、葉枯病、赤霉病4種病害)、越冬抗凍性、分蘗率、有效分蘗率、穗粒數(shù)、千粒重等主要農(nóng)藝性狀指標。
穗發(fā)芽抗性測定按照中華人民共和國農(nóng)業(yè)行業(yè)標準(NY/T 1739—2009)[20]進行。計算見公式(1)和(2)。

1.2.2 野生麥苗抗穗發(fā)芽種質(zhì)系的入選標準 本試驗野生麥苗抗穗發(fā)芽種質(zhì)系選擇標準:[1]遺傳穩(wěn)定;[2]相對穗發(fā)芽指數(shù)<0.05;[3]株高≤75 cm;[4]抗倒性為1級;[5]大田無病害發(fā)生;[6]越冬抗凍性為1級;[7]分蘗率≥2000%;[8]有效分蘗率≥80%;[9]穗粒數(shù)≥35;[10]千粒重≥45 g。
1.3 野生麥苗抗穗發(fā)芽種質(zhì)系的遺傳背景(親緣關(guān)系)分析
1.3.1 野生麥苗抗穗發(fā)芽種質(zhì)系遺傳的環(huán)境因素分析秋后萌發(fā)出土野生麥苗種子的長休眠特性,充分說明了在自然界小麥生產(chǎn)龐大的群體中存在著具有強種子休眠性的高抗穗發(fā)芽基因及表型。分析這些高抗穗發(fā)芽基因的來源:①可能是小麥本身基因發(fā)生的突變,但這種可能性幾率很低;②小麥的伴生雜草野麥子(野燕麥Avena.fatua、節(jié)節(jié)麥Aegilops tauschiiCoss.等)的種子都具有極強的休眠特性和抗穗發(fā)芽特性[12,16-17]。這些與小麥屬于同一亞科的抗穗發(fā)芽植物,在長期與小麥的共同生長中存在著天然雜交的可能,通過天然雜交將種子的強休眠抗穗發(fā)芽基因轉(zhuǎn)給小麥的可能性較大。特別是節(jié)節(jié)麥和小麥分別屬于小麥族的山羊草屬與小麥屬,親緣關(guān)系較近,在小麥的進化史上,曾經(jīng)成功的通過屬間天然雜交為現(xiàn)代小麥貢獻了D 染色體組,因此,節(jié)節(jié)麥和小麥發(fā)生天然雜交的可能性極大。通過觀察大田中伴隨小麥生長的野燕麥和節(jié)節(jié)麥發(fā)現(xiàn),野燕麥的揚花期和小麥不同步(圖2B)所以野燕麥和小麥發(fā)生天然雜交的可能性極小。野燕麥和小麥分別屬于燕麥族和小麥族,親緣關(guān)系較遠,即使發(fā)生了遠緣雜交,不通過人工幼胚搶救其雜種幼胚也不能發(fā)育成苗;而大田中生長的節(jié)節(jié)麥的揚花期和小麥同步(圖2A),二者又有較近的親緣關(guān)系和成功發(fā)生天然雜交的歷史事實,因此現(xiàn)代發(fā)生天然雜交的可能性較大。四川農(nóng)業(yè)大學的劉登才等[13]在進行節(jié)節(jié)麥和普通小麥的遠緣雜交研究時,發(fā)現(xiàn)了有些節(jié)節(jié)麥類型和普通小麥遠緣雜交的幼胚不需要胚搶救培養(yǎng)也能形成種子并發(fā)育成苗,因此大田中抗穗發(fā)芽的野生麥苗極有可能是混合生長的節(jié)節(jié)麥和普通小麥的天然雜交后代。

圖2 大田中與小麥同時楊花的節(jié)節(jié)麥和野燕麥
1.3.2 野生麥苗抗穗發(fā)芽種質(zhì)系的疑似親本小麥品種的調(diào)查 調(diào)查收集地塊在收集野生麥苗抗穗發(fā)芽種質(zhì)系之前5~10 年間,種植過的小麥品種,這些種植過的小麥品種都有可能是和節(jié)節(jié)麥發(fā)生天然雜交的親本材料。然后采用這些疑似親本小麥品種的基因組DNA作參考進行遺傳分析。
1.3.3 野生麥苗抗穗發(fā)芽種質(zhì)系基因組DNA分子遺傳分析
(1)SSR分子標記引物的選擇
基于野生麥苗抗穗發(fā)芽種質(zhì)系遺傳的環(huán)境因素分析,在分子遺傳水平上重點檢測,野生麥苗抗穗發(fā)芽種質(zhì)系和節(jié)節(jié)麥的遺傳關(guān)系。因為節(jié)節(jié)麥和現(xiàn)代小麥具有同源的D染色體組,在發(fā)生天然雜交后,節(jié)節(jié)麥和小麥D染色體組間可以實現(xiàn)同源染色體配對,通過同源染色體間易位交換遺傳物質(zhì)的可能性極大。所以首先選用現(xiàn)有開發(fā)成功的小麥D 染色體組上的SSR 位點的引物進行PCR檢測。
(2)檢測材料DNA的提取與模板液制備
檢測用的‘節(jié)節(jié)麥’和被檢測野生麥苗抗穗發(fā)芽種質(zhì)系都來源于同一地塊。疑似親本小麥品種從該品種育種者處收集。采用SDS 法并參照《植物基因工程原理與技術(shù)》[21]從暗培養(yǎng)的黃花苗中提取DNA,純化后取少量的DNA 樣品溶于無菌的超純水中制成濃度約為20 ng/μL的模板液放于4℃冰箱中備用。
(3)SSR-PCR反應體系的構(gòu)建與反應程序
PCR 反應體系的構(gòu)建,采用25 μ L 反應體系,各成分的含量為:TaqDNA 聚合酶(5 U/μL) 0.2 μL、10×Buffer 2.5 μL、MgCl2(25 mmol/L) 2 μL、dNTP(2.5 mmoL) 1 μL、上下游引物(工作濃度)各1 μL、DNA 模板液(20 ng/μL) 0.5 μL、無菌超純水16.8 μL。PCR 反應在英國TECHNE 公司的TC-5000 PCR 儀上進行,反應程序為:94℃預變性5 min,94℃變性50 s,退火溫度(參考引物合成報告單推薦溫度微作調(diào)整)退火時間為50 s,72℃延伸1 min,(30個循環(huán)),72℃后延伸5 min,4℃保存。
(4)SSR-PCR反應產(chǎn)物的電泳檢測
PCR 反應產(chǎn)物采用8%聚丙烯酰胺凝膠進行電泳檢測,每泳道上樣量為8 μL,在200 V 恒電壓下電泳5 h,剝膠,銀染,照相,進行擴增條帶的分析。
2 試驗結(jié)果與分析
2.1 野生麥苗收集移栽與遺傳穩(wěn)定性統(tǒng)計
由表1數(shù)據(jù)知,當年的9月底10月初,在大田中生長的野生麥苗,根部帶土移栽很容易成活,通過4年的移栽試驗,不論是單棵生長還是成簇(穗)生長的野生麥苗,只要移栽后及時澆水成活率都為100%。
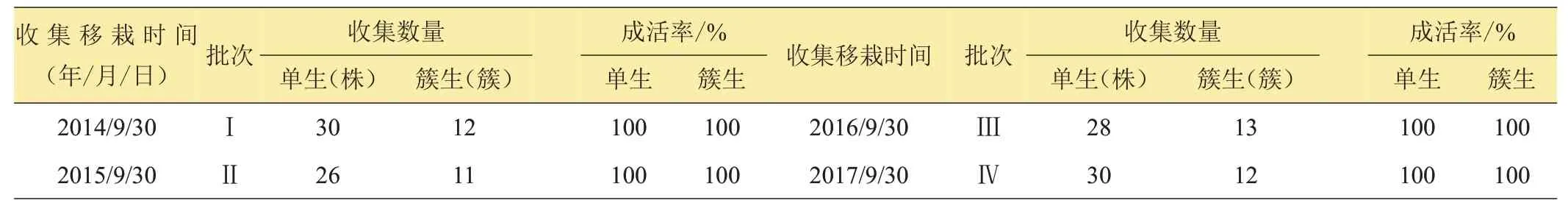
表1 野生麥苗收集移栽統(tǒng)計表
由表2數(shù)據(jù)知,大田中簇生野生麥苗的簇系后代,是由落入土中的一個麥穗上的種子生長而來,在進行第一年大田遺傳穩(wěn)定性觀察時,90%以上簇系的后代都生長一致,不足10%簇系的后代中,出現(xiàn)個別性狀表現(xiàn)不一致的單株,本試驗中將后代生長不一致的簇系淘汰。第二年只對第一年未淘汰的簇系繼續(xù)進行大田生長一致性觀察。第一年后代生長一致的簇系,第二年其后代仍都生長一致。通過連續(xù)2年的大田生長一致性觀察,大田中成簇生長的野生麥苗90%以上的簇系都生長一致且能穩(wěn)定遺傳。
大田中單棵生長的野生麥苗,移栽后的第一年由于沒有形成后代群體,從第二年才開始進行后代生長一致性觀察。由表2 數(shù)據(jù)知,第二年在首次進行后代生長一致性觀察時,大田中單棵生長的野生麥苗只有63%以上的株系生長一致,本試驗中將生長不一致的株系淘汰。第三年只對第二年首次觀察未淘汰的株系進行第二次生長一致性觀察。第二年后代生長一致的株系,第三年其后代仍然生長一致。通過連續(xù)2 年的大田生長一致性觀察,大田中單棵生長的野生麥苗63%以上株系都能穩(wěn)定遺傳。
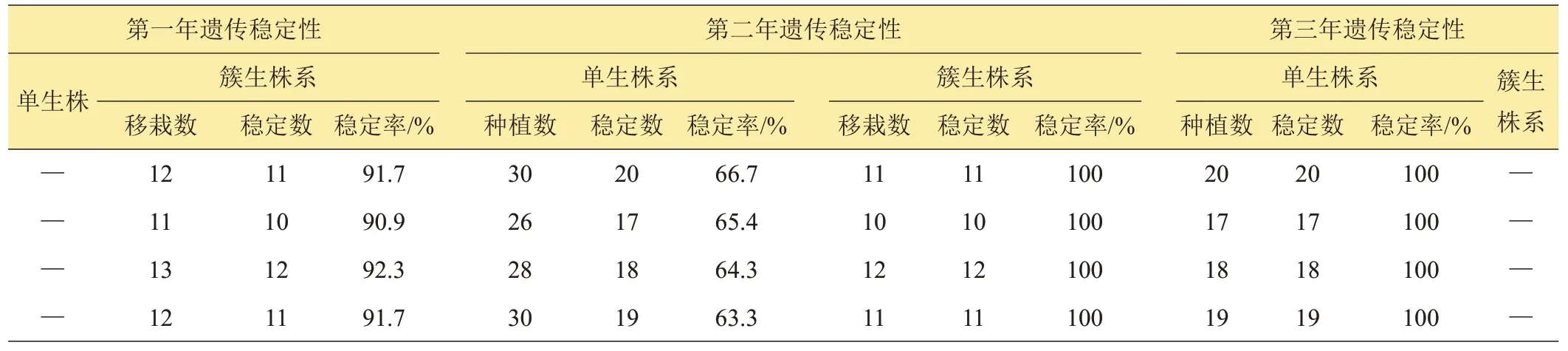
表2 野生麥苗遺傳穩(wěn)定性統(tǒng)計表
2.2 穩(wěn)定遺傳的野生麥苗系主要農(nóng)藝性狀調(diào)查及穗發(fā)芽抗性的測定結(jié)果
通過對生長一致、穩(wěn)定遺傳的118 個野生麥苗系主要農(nóng)藝性狀指標的調(diào)查,數(shù)據(jù)如表3。野生麥苗系的植株都比較高,有65.3%的野生麥苗系的株高都在100 cm 以上,有79.7%的野生麥苗系植株高于80 cm,20.4%的野生麥苗系株高低于80 cm,只有6.8%的野生麥苗系的植株不高于75 cm;野生麥苗系的抗倒性大部分較差,有65.3%的野生麥苗系發(fā)生中等以上倒伏,倒伏面積大于10%。21.2%的野生麥苗系倒伏較輕,只有13.6%的野生麥苗系不發(fā)生倒伏;野生麥苗系的抗病性(白粉病、銹病、葉枯病、赤霉病)普遍較好。在大田環(huán)境中,80.5%的野生麥苗系無病害發(fā)生,只有19.5%的野生麥苗系輕微發(fā)病;野生麥苗系的越冬抗凍性都較好,越冬凍害都不高于2 級,55.9%的野生麥苗系越冬凍害為1 級;野生麥苗系的分蘗率和有效分蘗率都較高,本試驗所有野生麥苗系的分蘗都不低于20 倍,有效分蘗率都不低于80%;野生麥苗系的穗粒數(shù)較多,本試驗所有的野生麥苗系穗粒數(shù)都不少于35粒,但野生麥苗系的千粒重較低,在行距20 cm,株距10 cm單穴單粒種植模式下,78.8%的野生麥苗系千粒重在45 g 以下,只有21.2%的野生麥苗系千粒重不低于45 g。

表3 穩(wěn)定遺傳的野生麥苗系主要農(nóng)藝性狀調(diào)查統(tǒng)計表
分析表4 中118 個野生麥苗系的穗發(fā)芽抗性測定結(jié)果,野生麥苗系的穗發(fā)芽抗性普遍較好。有19.5%的野生麥苗系的相對穗發(fā)芽指數(shù)為0,達到了特高抗水平。78.8%的野生麥苗系穗發(fā)芽抗性都能達到高抗水平,僅有17.8%和3.4%野生麥苗系的穗發(fā)芽抗性為抗和中抗水平。

表4 穩(wěn)定遺傳的野生麥苗系的穗發(fā)芽抗性測定結(jié)果統(tǒng)計表
2.3 野生麥苗抗穗發(fā)芽種質(zhì)系的篩選結(jié)果
根據(jù)本試驗制定的野生麥苗抗穗發(fā)芽種質(zhì)系的選擇標準,符合條件的野生麥苗抗穗發(fā)芽種質(zhì)系應為以下集合的交集:{野生麥苗抗穗發(fā)芽種質(zhì)系}={相對穗發(fā)芽指數(shù)<0.05的系}∩{株高≤75 cm的系}∩{抗倒性為1級的系}∩{無病害發(fā)生的系}∩{越冬抗凍性為1級的系}∩{分蘗率≥2000%的系}∩{有效分蘗率≥80%的系}∩{穗粒數(shù)≥35 的系}∩{千粒重≥45 g 的系}={21、46、63、106}。從后代生長一致、穩(wěn)定遺傳的118 個野生麥苗系中選出了‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’四個符合標準的野生麥苗抗穗發(fā)芽種質(zhì)系,其抗穗發(fā)芽效果如圖3所示。

圖3 野生麥苗抗穗發(fā)芽種質(zhì)系21、46、63、106抗穗發(fā)芽效果
2.4 野生麥苗抗穗發(fā)芽種質(zhì)系收集地塊以往種植情況調(diào)查
通過走訪調(diào)查收集抗穗發(fā)芽種質(zhì)資源系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’地塊的主人獲悉,從收集之日起前5~10年間,本地塊主要種植的小麥品種有‘濟麥22’、‘邯6172’、‘衡4399’。
2.5 野生麥苗抗穗發(fā)芽種質(zhì)系的SSR-PCR檢測結(jié)果

表5 發(fā)現(xiàn)野生麥苗抗穗發(fā)芽種質(zhì)系地塊以往種植情況調(diào)查
分析引物Xgwm493(檢測位點位于3B染色體)檢查結(jié)果(圖4A)、引物Xgwm538(檢測位點位于4B 染色體)檢查結(jié)果(圖4B)、引物Xgdm33(檢測位點位于1A、1D染色體)檢查結(jié)果(圖4 C),3對引物分別在4個抗穗發(fā)芽種質(zhì)系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’的基因組DNA中檢測到了和‘節(jié)節(jié)麥’一致而參考親本‘濟麥22’、‘邯6172’、‘衡4399’沒有的特異性條帶。引物Xgwm493在‘節(jié)節(jié)麥’、4個抗穗發(fā)芽種質(zhì)系的基因組DNA 中檢測到了約160 bp 大小(圖4A 中箭頭所示)的特異性條帶;引物Xgwm438 在‘節(jié)節(jié)麥’、4 個抗穗發(fā)芽種質(zhì)系的基因組DNA 中檢測到了約400 bp 大小(圖4 B 中箭頭所示)的特異性條帶;引物Xgdm33 在‘節(jié)節(jié)麥’、4 個抗穗發(fā)芽種質(zhì)系的基因組DNA中檢測到了約800、720、80 bp大小的特異性條帶(圖4 C 中箭頭所示);這充分證明了抗穗發(fā)芽種質(zhì)系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’和‘節(jié)節(jié)麥’的親緣關(guān)系。
引物Xgdm33(檢測位點位于1A、1D染色體)和引物Xgwm52(檢測位點位于3D 染色體)在4 個抗穗發(fā)芽種質(zhì)系的基因組DNA 中不僅檢測到了和‘節(jié)節(jié)麥’一致而參考親本‘濟麥22’、‘邯6172’、‘衡4399’沒有的特異性條帶,還檢測到了一些和全部或部分參考親本一致而‘節(jié)節(jié)麥’沒有的特異性條帶(圖4D、E 箭頭所示);引物Xgwm314(檢測位點位于3D染色體)檢測到了大量的和參考親本‘邯6172’、‘衡4399’一致而‘節(jié)節(jié)麥’沒有的特異性條帶(圖4E 箭頭所示)。這充分證明了4個抗穗發(fā)芽種質(zhì)系和‘濟麥22’、‘邯6172’、‘衡4399’的親緣關(guān)系。

圖4 SSR分子標記檢查結(jié)果
引物Xgdm33、Xgwm52、Xgwm314 在4 個抗穗發(fā)芽種質(zhì)系的基因組DNA中都檢測到了和‘邯6172’的親緣關(guān)系,因此,抗穗發(fā)芽種質(zhì)系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’是‘節(jié)節(jié)麥’和‘邯6172’天然雜交后代的可能性較大。
3 結(jié)論
本研究通過對大田野生麥苗集中移栽,遺傳穩(wěn)定性觀察與選擇,穩(wěn)定系的穗發(fā)芽抗性鑒定,篩選出了4 個對穗發(fā)芽有絕對抗性的小麥種質(zhì)資源系‘ZZX21’、‘ZZX46’、‘ZZX63’、‘ZZX106’,為小麥抗穗發(fā)芽育種提供了寶貴的種質(zhì)資源,豐富了小麥抗穗發(fā)芽育種遺傳的物質(zhì)基礎。
通過遺傳分析與分子標記檢測,從分子水平上初步證明了從大田野生麥苗中獲得的小麥高抗穗發(fā)芽種質(zhì)資源材料是當?shù)卮筇镏械囊吧?jié)節(jié)麥’和種植的普通小麥品種進行天然遠緣雜交的后代。
4 討論
4.1 前人研究結(jié)果
具有絕對抗性的高抗穗發(fā)芽種質(zhì)資源的匱乏是限制小麥抗穗發(fā)芽育種取得突破的主要障礙。獲得具有絕對抗性的高抗穗發(fā)芽種質(zhì)資源一直是小麥育種工作者努力的方向。首先對現(xiàn)有的大量小麥品種資源材料進行了抗穗發(fā)芽鑒定,以期篩選出抗穗發(fā)芽特性好的材料。中國農(nóng)業(yè)科學院品種資源研究所、西北農(nóng)林科技大學、西南大學生物科技學院、山西省小麥研究所等單位都在小麥抗穗發(fā)芽種質(zhì)資源的篩選鑒定方面做了大量的工作[3-8]。發(fā)現(xiàn)了一些穗發(fā)芽抗性與種子萌發(fā)生理上的關(guān)系,抗性與麥穗形態(tài)、穎殼、種皮顏色上的關(guān)系,也選出了一些抗性相對較好的材料。但是受現(xiàn)有小麥遺傳背景的限制并沒有從根本上找到抗穗發(fā)芽的抗源基因。
隨著生物技術(shù)的發(fā)展,基因工程技術(shù)應用于小麥抗穗發(fā)芽種質(zhì)資源的創(chuàng)制。美國農(nóng)業(yè)部農(nóng)業(yè)研究局和堪薩斯州立大學通過對小麥全基因組測序研究克隆成功了PHS 基因,轉(zhuǎn)PHS 小麥的穗發(fā)芽受到明顯抑制;國家小麥工程技術(shù)研究中心克隆的反義硫氧還蛋白基因(anti-trxs)轉(zhuǎn)入小麥后,獲得的00T89、01TY18 轉(zhuǎn)基因株系,穗發(fā)芽率較對照分別降低35.5%和62%[9-11]。基因工程技術(shù)拓寬了抗穗發(fā)芽育種遺傳的物質(zhì)基礎,具有廣闊的發(fā)展前景。出于轉(zhuǎn)基因安全性的考慮,目前轉(zhuǎn)基因技術(shù)在小麥這種主要糧食作物上的應用受到了國家的嚴格限制。
自然界中存在著和小麥同族不同屬的節(jié)節(jié)麥(分屬于小麥屬與山羊草屬)有著極強的高抗穗發(fā)芽特性[12,16-17]。利用節(jié)節(jié)麥的高抗穗發(fā)芽基因創(chuàng)造小麥高抗穗發(fā)芽種質(zhì)資源,一直是中國科技工作者研究的熱點。四川農(nóng)業(yè)大學的蘭秀錦[12]對節(jié)節(jié)麥的抗穗發(fā)芽基因進行了染色體定位研究,確定了節(jié)節(jié)麥的抗穗發(fā)芽基因是隱性單基因位于2D染色體上,以四倍體的地方小麥品種矮蘭麥做母本與采自河南的節(jié)節(jié)麥進行雜交,F(xiàn)1代用秋水仙素進行染色體加倍獲得了雙二倍體小麥RSP其穗發(fā)芽率僅為2.86%[14-15];河南大學的劉新浩[17]對節(jié)節(jié)麥的抗穗發(fā)芽特性進行了鑒定及遺傳多樣性研究,發(fā)現(xiàn)了生長于黃河中游的節(jié)節(jié)麥抗穗發(fā)芽特性都較國外品種好。河南大學的陳趁趁[18]對節(jié)節(jié)麥和小麥雜交后代的抗穗發(fā)芽特性進行了鑒定并對抗穗發(fā)芽基因所在的相關(guān)染色體片段分析,通過連續(xù)兩年對群體的穗發(fā)芽分析,篩選到20份高抗穗發(fā)芽的材料,5天發(fā)芽率均在10%以下,在5D和6D染色體上,存在著6 對與極抗穗發(fā)芽基因緊密連鎖的SSR 分子標記位點。以上研究都在節(jié)節(jié)麥和小麥的遠緣雜交后代中獲得了高抗甚至于極抗穗發(fā)芽材料,豐富了小麥抗穗發(fā)芽育種的遺傳背景,有效利用了節(jié)節(jié)麥中的極抗穗發(fā)芽基因,為小麥抗穗發(fā)芽育種提供了寶貴的種質(zhì)資源。
4.2 本研究較前人研究的先進性
本試驗研究了一種從自然存在的大田野生麥苗中收集、篩選小麥高抗穗發(fā)芽種質(zhì)資源的方法;并從分子水平上證明了大田野生麥苗中獲得的小麥高抗穗發(fā)芽種質(zhì)資源材料是當?shù)卮筇镏械囊吧?jié)節(jié)麥和大田種植的普通小麥品種進行天然遠緣雜交的后代,具有和節(jié)節(jié)麥相同的極強抗穗發(fā)芽特性。本技術(shù)方法簡單易行,成本低廉,有效地利用了節(jié)節(jié)麥中的遠緣極抗穗發(fā)芽基因資源,選出的小麥高抗穗發(fā)芽種質(zhì)資源的遺傳背景豐富多樣,為當前的小麥抗穗發(fā)芽育種提供了寶貴有絕對抗性的抗穗發(fā)芽種質(zhì)資源。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
