虹鱒幼魚(yú)對(duì)不同淀粉的利用
2022-08-30 02:01:34黨江雨蔡友旺張春燕曹凱林李小勤冷向軍
水生生物學(xué)報(bào) 2022年8期
黨江雨 蔡友旺 張春燕 曹凱林 李小勤 冷向軍
(上海海洋大學(xué)水產(chǎn)科學(xué)國(guó)家級(jí)教學(xué)示范中心, 農(nóng)業(yè)農(nóng)村部魚(yú)類營(yíng)養(yǎng)與環(huán)境生態(tài)研究中心,水產(chǎn)動(dòng)物遺傳育種中心上海市協(xié)同創(chuàng)新中心, 上海 201306)
魚(yú)類對(duì)糖的利用能力有限[1], 但是糖類作為一種廉價(jià)能量源, 不僅可以降低飼料成本, 而且對(duì)于魚(yú)類的生長(zhǎng)也有一定的促進(jìn)作用[2—4]。對(duì)虹鱒(Oncorhynchus mykiss)[5]、大西洋鮭(Salmo salar)[6]、青魚(yú)(Mylopharyngodon piceus)[7]、大黃魚(yú)(Larimichthys crocea)[8]和達(dá)氏鱘(Acipenser dabryanus)[9]等的研究表明, 淀粉等大分子糖類相較于葡萄糖等小分子糖類對(duì)于魚(yú)類的促生長(zhǎng)作用更明顯, 而且淀粉具有優(yōu)良的黏合性和可消化性。淀粉由直鏈淀粉和支鏈淀粉組成[10], 來(lái)源不同的淀粉, 在結(jié)構(gòu)組成存在差異, 其直/支鏈淀粉比例不同, 魚(yú)類對(duì)其利用程度也不相同[11—13]。在虹鱒[14]、羅非魚(yú)(Oreochromis mossambicus)[15]和草魚(yú)(Ctenopharyngodon idella)[16]上的研究表明, 這幾種魚(yú)類對(duì)支鏈淀粉的利用程度更高, 而條紋雜交鱸(Morone chrysops×M.saxatilis)[17]對(duì)直鏈淀粉的利用能力更高。
虹鱒屬鮭形目(Salmoniformes), 鮭科(Salmonidae), 大麻哈魚(yú)屬(Oncorhynchus), 是世界范圍內(nèi)廣泛養(yǎng)殖的冷水性魚(yú)類。研究表明, 虹鱒對(duì)于飼料中碳水化合物的適宜需要量為≤20%[1]; 在虹鱒對(duì)淀粉的利用方面也有一些報(bào)道, 例如不同種類淀粉對(duì)其消化率的影響[18]、淀粉和脂肪的比例對(duì)其生長(zhǎng)等的影響[19]和不同水平的淀粉和葡萄糖對(duì)其糖代謝等的影響[5]等, 但尚未見(jiàn)虹鱒對(duì)不同種類淀粉源利用比較的報(bào)道。
本試驗(yàn)分別以木薯淀粉、小麥淀粉、豌豆淀粉和玉米淀粉為淀粉源, 設(shè)置相同的淀粉水平, 考察不同種類淀粉對(duì)虹鱒生長(zhǎng)、魚(yú)體組成、血液生化指標(biāo)和肝臟糖代謝酶等的影響, 探討虹鱒飼料適宜的淀粉來(lái)源, 為虹鱒高效飼料的配制提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)設(shè)計(jì)與試驗(yàn)飼料
本試驗(yàn)分別以木薯淀粉、小麥淀粉、豌豆淀粉和玉米淀粉為淀粉源, 配制淀粉添加水平為15%的4組等氮等脂飼料。飼料原料經(jīng)粉碎、過(guò)篩(60目)后, 按配方比例稱取原料進(jìn)行混合, 用單螺桿擠壓機(jī)制成直徑為2 mm的沉性顆粒飼料[制粒溫度(85±5)℃], 制粒后的飼料用烘干機(jī)37℃烘至水分含量低于10%(各組保持一致的烘干時(shí)間), 室溫陰涼干燥保存?zhèn)溆谩T囼?yàn)飼料的配方組成及營(yíng)養(yǎng)成分含量見(jiàn)表 1。

表1 試驗(yàn)飼料配方組成及營(yíng)養(yǎng)成分含量(風(fēng)干基礎(chǔ), g/kg)Tab. 1 Ingredients and proximate composition of experimental diets (air-dry basis, g/kg)
1.2 試驗(yàn)魚(yú)和飼養(yǎng)管理
試驗(yàn)用虹鱒購(gòu)自昆明倘浩水產(chǎn)養(yǎng)殖有限公司。試驗(yàn)魚(yú)暫養(yǎng)馴化2周后, 饑餓24h, 挑選276尾大小均勻, 體格健壯的虹鱒(7.7±0.1) g, 平均分配到12個(gè)自動(dòng)充氣循環(huán)水桶內(nèi)(直徑1.0 m, 水深0.8 m,水體約為650 L), 試驗(yàn)共4個(gè)處理組, 每組設(shè)3個(gè)平行, 每個(gè)平行23尾魚(yú)。養(yǎng)殖期間每天投喂兩次(9:00和16:00), 日投喂量約為魚(yú)體重的2%—5%, 根據(jù)水溫和虹鱒攝食情況進(jìn)行適當(dāng)調(diào)整, 每次投喂以虹鱒不搶食為宜。每次于投喂2—3h后用虹吸法吸走桶底糞便, 每周換水3次, 換水量為循環(huán)系統(tǒng)的1/3。養(yǎng)殖期間水溫 12—16℃, 溶氧 6—7 mg/L, pH 7.24—7.78, 氨氮≤0.2 mg /L, 亞硝酸鹽≤0.1 mg /L。養(yǎng)殖試驗(yàn)于2020年冬季在上海海洋大學(xué)濱海養(yǎng)殖基地循環(huán)水車間進(jìn)行, 養(yǎng)殖試驗(yàn)共持續(xù)56d。
1.3 樣品采集
在養(yǎng)殖試驗(yàn)結(jié)束后, 魚(yú)體饑餓24h, 統(tǒng)計(jì)每桶中虹鱒數(shù)量并稱重, 計(jì)算增重率、飼料系數(shù)和成活率。每桶隨機(jī)取9尾魚(yú), 用MS-222麻醉(100 mg/L),取3尾魚(yú)裝袋后于–20℃保存, 用于全魚(yú)常規(guī)成分的測(cè)定; 另取3尾魚(yú), 測(cè)量體重、體長(zhǎng)、內(nèi)臟重和肝臟重, 用經(jīng)過(guò)肝素鈉潤(rùn)洗的注射器從尾靜脈處采血,3500 r/min離心10min, 取血清于–80℃保存; 取部分肝臟于波恩固定液中固定, 用于組織學(xué)觀察, 剩余肝臟于–20℃保存, 用于測(cè)定肝糖原含量和肝臟糖代謝酶活性; 其余3尾魚(yú), 抽血后(防止解剖時(shí)血液污染樣品)采集肝臟樣本于–20℃保存, 用于測(cè)定肝臟組成。
在采樣結(jié)束后, 將剩余虹鱒繼續(xù)投喂3d, 使其恢復(fù)正常的生理狀態(tài)。在饑餓24h后, 飽食投喂1次,于投喂前(0)及投喂后1h、3h、5h、7h和9h, 每桶隨機(jī)取1尾虹鱒(每處理組每個(gè)時(shí)間點(diǎn)為3尾魚(yú)), 用MS-222麻醉(100 mg/L), 使用經(jīng)肝素鈉潤(rùn)洗的注射器從尾靜脈處采血, 3500 r/min離心10min后取血清,于–80℃保存, 進(jìn)行血糖含量的測(cè)定。
1.4 測(cè)定指標(biāo)與方法
生長(zhǎng)性能與形體指標(biāo)成活率SR(%)=100×終末尾數(shù)(尾)/初始尾數(shù)(尾);
增重率WGR(%)=100×[終末體重(g)–初始體重(g)]/初始體重(g);
飼料系數(shù)FCR=采食量(g)/[終末體重(g)–初始體重(g)];
臟體比VSI=100×內(nèi)臟重(g)/體重(g);
肝體比HSI=100×肝臟重(g)/體重(g);
肥滿度CF(g/cm3)=100×體重(g)/體長(zhǎng)(cm)3;
蛋白質(zhì)沉積率PR(%)=100×魚(yú)體蛋白質(zhì)貯積量/攝入的蛋白質(zhì)總量;
脂肪沉積率LR(%)=100×魚(yú)體脂肪貯積量/攝入的脂肪總量。
全魚(yú)、肝臟與飼料組成全魚(yú)與飼料的水分、粗蛋白含量測(cè)定分別采用105℃常壓干燥法和凱氏定氮法(2300自動(dòng)凱氏定氮儀, FOSS, 瑞典), 粗脂肪含量測(cè)定采用氯仿-甲醇抽提法, 粗灰分測(cè)定采用550℃高溫灼燒法。
肝糖原含量測(cè)定將肝臟樣本于4℃解凍后,取0.1 g左右肝臟, 將肝臟與堿液以1﹕3的體積混合,在沸水浴中加熱20min, 得到水解液, 采用南京建成生物工程研究所試劑盒測(cè)定肝糖原含量。
血清生化指標(biāo)測(cè)定血清葡萄糖、甘油三酯和總膽固醇含量及谷草轉(zhuǎn)氨酶和谷丙轉(zhuǎn)氨酶活性采用南京建成生物工程研究所的試劑盒和酶標(biāo)儀(BioTeK Ynergy HTX, 美國(guó))測(cè)定。
肝臟糖代謝酶活性的測(cè)定取肝臟樣本于4℃解凍, 勻漿, 4℃離心(3000 r/min, 10min), 取上清液用于肝臟己糖激酶(HK)、磷酸果糖激酶(PFK)、丙酮酸激酶(PK)、磷酸烯醇式丙酮酸激酶(PEPCK)和葡萄糖-6-磷酸酶(G6Pase)活性測(cè)定。采用南京建成生物工程研究所的試劑盒和酶標(biāo)儀(BioTeK Ynergy HTX, 美國(guó))測(cè)定上述酶活性。
肝臟組織切片石蠟切片HE染色: 將肝臟樣品從固定液中取出, 用不同濃度的酒精、酒精和二甲苯混合液、二甲苯逐級(jí)脫水, 之后進(jìn)行浸蠟、包埋、切片、染色等工作, 染色劑為蘇木精-伊紅。染色后裝片, 待干燥后觀察肝臟的組織形態(tài),并拍照。
1.5 數(shù)據(jù)處理
試驗(yàn)數(shù)據(jù)以平均數(shù)±標(biāo)準(zhǔn)差表示, 采用SPSS25.0軟件進(jìn)行單因素方差分析, 結(jié)果用“平均值±標(biāo)準(zhǔn)誤”表示, 其中差異顯著者進(jìn)行Duncan多重比較, 差異顯著水平為P<0.05。
2 結(jié)果
2.1 生長(zhǎng)性能與形體指標(biāo)
在養(yǎng)殖過(guò)程中, 各組虹鱒存活率均為100%(表 2)。木薯淀粉組增重率最高(1049.3%), 飼料系數(shù)(0.83)最低; 與木薯淀粉組相比, 小麥淀粉組、豌豆淀粉組和玉米淀粉組的增重率顯著降低, 飼料系數(shù)顯著升高(P<0.05), 該3組在增重率和飼料系數(shù)上無(wú)顯著差異(P>0.05)。各組在肥滿度、臟體比和肝體比之間也無(wú)顯著差異(P>0.05)。
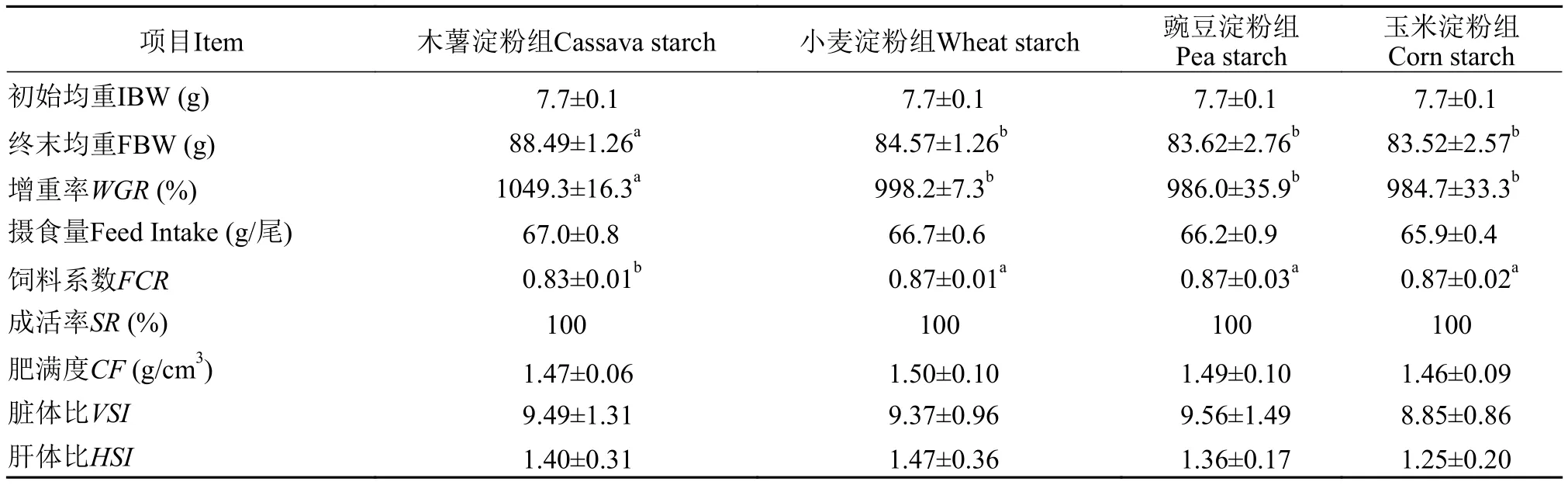
表2 飼料淀粉種類對(duì)虹鱒生長(zhǎng)性能和形體指標(biāo)的影響Tab. 2 Effects of dietary starches on growth and morphological indices of rainbow trout
2.2 魚(yú)體、肝臟組成
由表 3可見(jiàn), 各組在全魚(yú)水分、粗蛋白質(zhì)、粗脂肪和粗灰分含量上均無(wú)顯著差異(P>0.05)。在營(yíng)養(yǎng)物質(zhì)利用方面, 各組蛋白質(zhì)和脂肪沉積率無(wú)顯著差異(P>0.05)。肝臟水分、粗蛋白質(zhì)和粗脂肪含量也不受淀粉種類影響(P>0.05)。豌豆淀粉組肝糖原含量顯著低于其余各組(P<0.05); 木薯淀粉組、小麥淀粉組和玉米淀粉組的肝糖原含量無(wú)顯著差異(P>0.05)。

表3 飼料淀粉種類對(duì)虹鱒全魚(yú)和肝臟組成的影響(鮮重, %)Tab. 3 Effects of dietary starches on body and liver composition of rainbow trout (fresh weight, %)
2.3 血清生化指標(biāo)
由表 4可見(jiàn), 各組血清甘油三酯、總膽固醇含量不受淀粉種類影響, 谷草轉(zhuǎn)氨酶和谷丙轉(zhuǎn)氨酶活性也無(wú)顯著差異(P>0.05)。

表4 飼料淀粉種類對(duì)虹鱒血液生化指標(biāo)的影響Tab. 4 Effects of dietary starches on serum biochemical indices of rainbow trout
2.4 肝臟糖代謝酶活性
由表 5可見(jiàn), 各組丙酮酸激酶、葡萄糖-6-磷酸酶和磷酸烯醇式丙酮酸羧激酶活性均不受淀粉種類影響(P>0.05)。木薯淀粉組和小麥淀粉組己糖激酶活性顯著高于豌豆淀粉組和玉米淀粉組(P<0.05),磷酸果糖激酶活性顯著高于玉米淀粉組(P<0.05)。
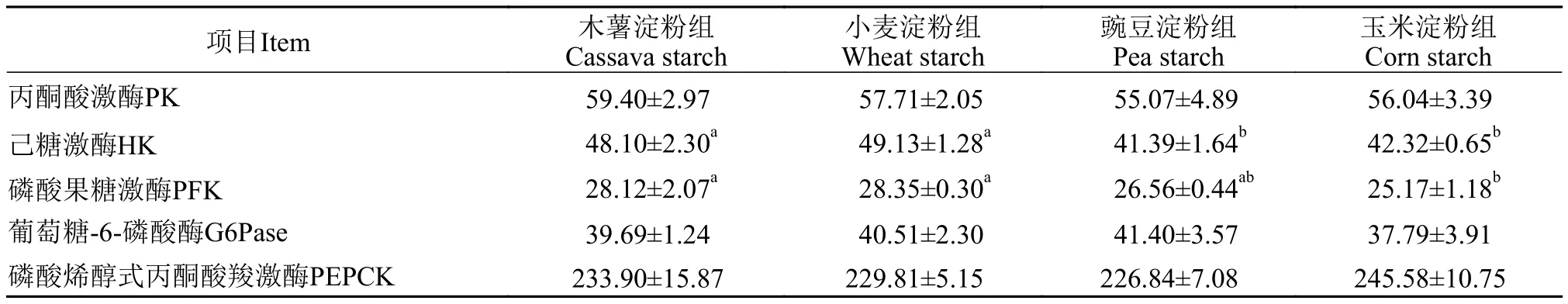
表5 飼料淀粉種類對(duì)虹鱒肝臟糖代謝酶活性的影響Tab. 5 Effects of dietary starches on activities of carbohydrate metabolic enzymes in the liver of rainbow trout (U/mg prot)
2.5 餐后血糖含量變化
由表 6和圖 1可見(jiàn), 各組虹鱒在攝食后, 血糖含量均呈現(xiàn)先上升后下降的趨勢(shì), 在攝食后7h血糖含量達(dá)到最高。木薯淀粉組在攝食后1—7h的血糖含量均顯著高于其余3組(P<0.05)。攝食9h時(shí), 各組血糖含量無(wú)顯著差異(P>0.05)。從雙因素分析來(lái)看,餐后時(shí)間和淀粉種類均對(duì)血糖含量存在顯著影響(P<0.05), 但二者交互作用無(wú)顯著影響(P>0.05)。
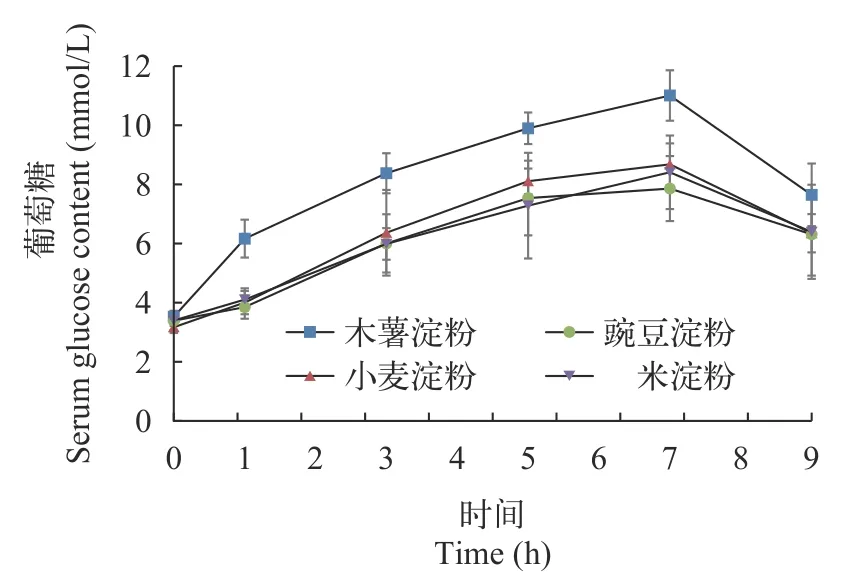
圖1 虹鱒餐后血糖含量變化Fig. 1 The serum glucose content of rainbow trout after feeding

表6 飼料淀粉種類對(duì)虹鱒攝食后血糖含量的影響Tab. 6 Effects of dietary starches on postprandial serum glucose content of rainbow trout (mmol/L)
2.6 肝臟組織學(xué)
由圖 2可見(jiàn), 各組肝臟組織形態(tài)正常, 未出現(xiàn)肝細(xì)胞腫脹、空泡化和細(xì)胞核位移等現(xiàn)象。

圖2 虹鱒肝臟組織切片F(xiàn)ig. 2 Tissue slices of rainbow trout liver (200×)
3 討論
3.1 不同種類淀粉對(duì)虹鱒生長(zhǎng)性能的影響
淀粉的來(lái)源廣泛, 不同種類的淀粉其直鏈淀粉和支鏈淀粉的含量、顆粒大小、分子結(jié)構(gòu)等方面均有不同, 直/支鏈淀粉的比例會(huì)影響魚(yú)類對(duì)淀粉的利用[20]。在本試驗(yàn)選用的4種淀粉中, 木薯淀粉的支鏈淀粉含量最高。從生長(zhǎng)性能來(lái)看, 木薯淀粉組的魚(yú)體增重率顯著高于小麥淀粉組、豌豆淀粉組和玉米淀粉組, 飼料系數(shù)也最低, 這表明虹鱒對(duì)支鏈淀粉含量高的淀粉源有較好的利用效果。在Preffer等[14]的研究中, 虹鱒對(duì)蠟質(zhì)玉米淀粉(1%直鏈淀粉和99%支鏈淀粉)的利用能力高于普通玉米淀粉(28%—30%直鏈淀粉和70%—72%支鏈淀粉),此外, 在草魚(yú)[16]和羅非魚(yú)[21]中也有類似的報(bào)道。但在大口黑鱸(Micropterus salmoides)[22]的研究中, 高直鏈/支鏈比的淀粉對(duì)魚(yú)類的生長(zhǎng)反而有促進(jìn)作用,其原因可能是過(guò)多的支鏈淀粉水解造成了攝食后血糖水平過(guò)高, 不利于大口黑鱸的正常生長(zhǎng)。茍仕潘等[23]的研究表明, 大口黑鱸[初始體重(28.36±0.10) g]飼料中適宜的可消化淀粉水平為9.22%—10.05%,而虹鱒[初始體重(28.4±3.2) g]飼料中的適宜淀粉水平分別為18%(15%粗脂肪)和27%(18%粗脂肪)[19],可見(jiàn), 虹鱒較大口黑鱸更能耐受高糖。
在本試驗(yàn)中, 各組肝臟組織學(xué)無(wú)顯著差異, 表明在15%的淀粉水平下, 肝臟均處于正常生理狀態(tài), 但豌豆淀粉組肝糖原含量顯著低于其余3個(gè)試驗(yàn)組, 原因可能是豌豆淀粉的直鏈淀粉含量高, 直鏈淀粉不易被消化分解, 從而降低了糖原在肝臟中的積累, 這與Song等[24]和徐祥泰等[25]在大口黑鱸上的研究結(jié)果類似。
3.2 不同種類淀粉對(duì)虹鱒魚(yú)體組成和血液生化指標(biāo)的影響
在本試驗(yàn)中, 各試驗(yàn)組在全魚(yú)和肝臟常規(guī)營(yíng)養(yǎng)組成方面沒(méi)有顯著差異, 并且血液的總膽固醇和甘油三酯含量也不受淀粉種類影響, 而這兩個(gè)指標(biāo)代表著魚(yú)類脂肪代謝情況, 表明不同種類的淀粉對(duì)蛋白質(zhì)和脂肪的轉(zhuǎn)化利用沒(méi)有產(chǎn)生顯著影響。任鳴春[5]采用小麥淀粉、木薯淀粉和玉米淀粉3種糊化淀粉源飼喂軍曹魚(yú)(Rachycentron canadum), 對(duì)全魚(yú)組成也未產(chǎn)生顯著影響; Song等[24]采用小麥淀粉、豌豆淀粉、木薯淀粉和高直鏈玉米淀粉為淀粉源飼喂大口黑鱸, 各組全魚(yú)組成之間也無(wú)顯著差異。
谷草轉(zhuǎn)氨酶和谷丙轉(zhuǎn)氨酶是反映肝功能的重要指標(biāo), 當(dāng)肝臟受損時(shí), 肝臟中的這兩種酶會(huì)釋放到血液中, 從而使血液中兩種酶活性升高。在本試驗(yàn)中, 各試驗(yàn)組血液中的谷草轉(zhuǎn)氨酶和谷丙轉(zhuǎn)氨酶活性沒(méi)有顯著差異, 在解剖過(guò)程中, 各處理組的肝臟從外觀上也沒(méi)有發(fā)現(xiàn)明顯差異。
3.3 不同種類淀粉對(duì)虹鱒糖代謝酶和餐后血糖水平的影響
本試驗(yàn)測(cè)定了己糖激酶(HK)、磷酸果糖激酶(PFK)、丙酮酸激酶(PK)、磷酸烯醇式丙酮酸激酶(PEPCK)和葡萄糖-6-磷酸酶(G6Pase)的酶活性, 前3種酶屬于糖酵解途徑, 后兩種酶存在于糖異生途徑。HK是糖酵解途徑的第一個(gè)限速酶。在本試驗(yàn)中, 木薯淀粉組HK活性顯著高于豌豆淀粉組和玉米淀粉組, PFK活性顯著高于玉米淀粉組, 并且餐后1—7h的血糖含量也顯著高于其余3組, 表明較高的血糖水平對(duì)HK和PFK產(chǎn)生了誘導(dǎo), 使其活性增加。小麥淀粉組的血糖水平并不高, 但HK和PFK活性較高, 其原因尚不清楚, 有待進(jìn)一步研究。PK是糖酵解最后一步的限速酶, 在本試驗(yàn)中,各組PK活性不受淀粉種類影響, 說(shuō)明其無(wú)法呼應(yīng)較高的血糖水平, Panserat等[26]曾報(bào)道, 虹鱒在攝食不含糖與含糖飼料后6h和24h, 其PK基因表達(dá)均無(wú)顯著差異。但在歐洲海鱸(Dicentrarchus labrax)[27]的研究表明, 飼料糖水平能夠調(diào)控PK的活性, 這可能是魚(yú)的種類和糖的種類不同造成的。PEPCK和G6Pase是糖異生途徑的代謝酶, 有研究發(fā)現(xiàn)[26], 虹鱒在攝食含糖和不含糖的飼料后, 其肝臟中PEPCK基因表達(dá)量均無(wú)變化, 說(shuō)明飼料中的糖對(duì)其沒(méi)有顯著影響; 飼料中糖種類對(duì)歐洲海鱸[28]G6Pase活力也無(wú)顯著影響。在本試驗(yàn)中, 各組PEPCK和G6Pase活性差異不顯著, 說(shuō)明這兩種酶活性不受淀粉種類的影響。
有研究表明, 當(dāng)海鯛(Sparus aurata)[29]和暗紋東方鲀(Takifugu obscurus)[30]長(zhǎng)期處于高血糖狀態(tài)時(shí), 會(huì)導(dǎo)致肝糖原異常積累, 對(duì)魚(yú)類的生長(zhǎng)不利, 在本試驗(yàn)中, 木薯淀粉組虹鱒的餐后1—7h血糖高于其余3個(gè)淀粉組, 但在9h后, 各組并無(wú)顯著差異, 木薯淀粉組在攝食表現(xiàn)和肝臟健康上都與其他各組無(wú)顯著差異, 且生長(zhǎng)性能最好, 這可能是虹鱒對(duì)糖的利用性和耐受性較高的緣故。Bergot[31]的研究表明, 攝食含30%葡萄糖組飼料虹鱒增重率、飼料效率和蛋白質(zhì)效率高于15%葡萄糖組, 也表明了在其耐受范圍內(nèi), 較高水平的糖類可能有利于提高虹鱒的生長(zhǎng)和飼料利用。
4 結(jié)論
在15%的淀粉水平下, 綜合考慮增重率、血液生化指標(biāo)、糖代謝酶和肝臟組織學(xué)等指標(biāo), 木薯淀粉較小麥淀粉、豌豆淀粉和玉米淀粉更適宜作為虹鱒飼料的淀粉來(lái)源。
