越冬脅迫對草魚抗氧化能力及脂肪酸組成的影響
2022-08-30 02:01:36武文一
水生生物學報 2022年8期
關鍵詞:氧化應激
武文一 吉 紅
(西北農林科技大學動物科技學院, 楊凌 712100)
在水產養殖中, 越冬(尤其是北方地區)是一個特殊的階段, 魚類遭受多種越冬應激影響, 例如食物匱乏、水溫低和光照[1]。魚類的免疫反應隨著季節變化而變化, 冬季魚類的免疫功能受到抑制, 同時造成較高的氧化應激反應, 進而導致魚類高死亡率[2]。魚類動員機體儲備的能量物質, 以度過食物短缺時期, 例如在越冬期間[3]。這種動員不僅僅包括肝胰臟, 還包括肌肉和脂肪組織中的脂質分解;以肌肉中蛋白質為主要能量來源; 或者將肝糖原或肌糖原分解成葡萄糖作為能量底物[4]。根據本實驗室的前期的研究結果, 越冬期間, 脂質動員是作為魚類應對越冬脅迫的最重要能量供應方式之一, 自始至終從越冬開始到越冬結束維持魚體能量之穩態[5]。有研究表明, 魚類在長期營養匱乏脅迫下, 動員能量抵御不利應激的同時, 誘導機體產生了活性氧自由基(ROS), 長期累積造成氧化應激[6,7]。過多的ROS會破壞機體抗氧化系統的平衡以及穩定, 造成細胞的損傷、細胞核的凋亡以及脂肪酸過氧化[8,9]。尤其在越冬期間作為主要供能物質的脂肪酸, 其供能作用受到巨大威脅以及挑戰。最直接結果就是降低供能水平, 造成機體抗氧化力以及免疫力的進一步降低, 故而提高了死亡率[10]。越冬期間造成了氧化應激的過度產生, 提高了魚體死亡, 因此探索并解決越冬脅迫對魚體抗氧化力乃至免疫力之間的相互作用關系具有重要的意義。
草魚(Ctenopharyngodon idellus)作為我國主要經濟淡水魚之一, 其產量居2019年中國養殖魚類產量的榜首, 達553.3×107kg[11]。由于草魚生長迅速,繁育技術較為成熟, 價格親民, 營養豐富, 因此深受養殖者以及消費者的歡迎[12]。但是在越冬脅迫下面臨著魚體內ROS的過量生成, 誘導了氧化應激脅迫。而過度的氧化應激可能導致機體免疫力下降,誘發疾病甚至死亡率的提高, 不利于草魚水產養殖業的健康發展。為了建立有效的策略, 并指導管理決策以改善該草魚的越冬存活率, 因而需要更好地了解越冬驅動的高度氧化應激的相互作用。因此,本研究以草魚為研究對象, 探索越冬條件下氧化應激對草魚免疫和抗氧化能力的影響, 以及與機體脂肪酸組成變化之間的相互聯系, 為緩解草魚越冬期間產生過度的氧化應激的反應以及降低魚體死亡率提供相關的理論依據。
1 材料與方法
1.1 實驗材料
實驗用魚均取自西北農林科技大學安康水產試驗示范站室外培育池塘, 選擇同一批次養成商品規格的草魚, 挑選大小整齊以及健壯的個體作為實驗對象, 實驗魚體重(1053.33±16.11) g。同時在室外培育水泥池中馴化兩周后開始實驗, 馴化期間每日正常投喂商品飼料(粗蛋白28%, 粗脂肪6%)。
1.2 實驗條件和方法
實驗在室外培育水泥池中進行(4 m×4 m×1 m),選取8個實驗池, 中間用隔網隔開, 上面架設遮陽網,水深控制0.50 m左右, 水容量8 m3左右。經過2周投飼馴化后, 停食1天進行分組; 隨機分為7組, 每組3個重復, 每個重復15尾魚。當水溫自然下降到草魚停止攝食(水溫: 15℃)時, 實驗開始; 當水溫自然升高到草魚開始重新覓食(水溫: 15℃)時實驗結束,此過程即為越冬期。實驗共計7 組, 分別為饑餓0周(Week 0)、1周(Week 1)、2周(Week 2)、4周(Week 4)、8周(Week 8)、12周(Week 12)和16周(Week 16)。實驗用水為曝氣后井水, 流水環境, 每2周清污一次。每日定時監測水溫、水質, 同時觀察實驗魚魚體的健康狀況以及死亡狀況。實驗用水條件:pH 7.8—8.2, 溶解氧5—6 mg/L, 氨氮<0.1 mg/L, 亞硝酸鹽<0.01 mg/L, 硫化物<0.05 mg/L。
1.3 樣品采集
采樣程序經過西北農林科技大學動物保護與利用委員會批準, 同時按照動物福利與道德規范進行執行。在各實驗組饑餓處理結束后, 所有魚均用50 mg/L MS222麻醉后, 再對其進行取樣。每組實驗魚, 逐一測量體質量、體長和全長等指標,計算肥滿度。每個平行隨機抽取2尾魚進行尾靜脈采血, 4℃冰箱靜置2h后, 4℃離心(3000 r/min,15min), 上層澄清透明淡黃色的血清速凍于液氮中, 而后轉入–80℃冰箱保存, 用作血清抗氧化酶活性指標測定; 每個平行隨機抽取2尾魚在冰盤上進行解剖, 取其內臟團, 分離肝胰臟、腎、脾及腹腔脂肪等組織, 稱重, 同時量取腸道長度, 計算臟體比、肝體比、腎指數、脾指數、腹腔脂肪指數及腸體比等指標。隨后在上述分離組織中, 取部分肝胰臟、肌肉、前腸和腹腔脂肪組織速凍于液氮中, 隨后轉–80℃冰箱保存, 用作酶活性測量及脂肪酸的測定。
1.4 測定指標和方法
根據以下公式, 計算肥滿度、肝體比、臟體比、腸體比、腎指數、脾指數和腹腔脂肪指數:
肥滿度(Condition factor,CF, g/cm3)=魚體重×100/體長3;
肝體比(Hepatosomatic index,HSI)=肝胰臟重×100/魚體重;
臟體比(Viscerosomatic index,VSI)=內臟重×100/魚體重;
腸體比(Relative intestine length,RIL)=腸長×100/體長;
腎指數(Kidney index,KI)= 腎臟重×100/魚體重;
脾指數(Spleen index,SI)= 脾臟重×100/魚體重;
腹腔脂肪指數(Intraperitoneal fat body index,IPFI)=腹腔脂肪重×100/魚體重;
成活率(Survival rate,SR, %)=最終魚數×100/初始魚數。
肝胰臟、肌肉、前腸、脂肪組織和血清中的丙二醛(MDA)、過氧化氫酶(CAT)和總超氧化物歧化酶(SOD)活性/含量使用南京建成生物工程研究所試劑盒測定(貨號分別是: A003-1-2、A007-1-1和A001-3-2), 谷胱甘肽 S-轉移酶(GST)活性和超氧陰離子含量使用北京索萊寶科技有限公司試劑盒測定(貨號分別是: SN101和BC1295), 蛋白質濃度使用南京建成生物工程研究所考馬斯亮藍染色法試劑盒(貨號: A045)測定。
脂肪酸測定分析過程中, 脂肪提取參照Tian等[13]的方法。取組織樣品大約5 g, 放置于10 mL離心管;加入氯仿?甲醇混合液(體積比2﹕1)溶液5 mL, 高速勻漿后, 搖床震蕩2h使之充分混勻, 隨后再用定量濾紙過濾。加入4 mL 去離子水, 震蕩混勻后在離心機上進行3000 r/min離心5min, 棄上清液及中間組織蛋白層, 下層40℃水浴蒸干。蒸干后加入1 mL正己烷(色譜純)溶解提取出的油脂。油脂的皂化及甲酯化參照Tian等[13]的方法, 加入1 mL 氫氧化鉀/甲醇溶液(0.4 mol/L), 再進行30min的甲酯化。最終加入2 mL去離子水, 震蕩混勻后在離心機上進行3000 r/min離心5min, 取上層溶液–20℃保存。樣品首先皂化以及甲酯化后, 使用氣相色譜儀(Agilent7820a, Agilent Technologies, Santa Clara, CA, USA)進行測定。脂肪酸甲酯樣品, 通過與脂肪酸甲酯混合標準品(47015-U, Sigma-Aldrich, USA)進行比較鑒定。取1 μL混合物進行上機檢測。脂肪酸組成的相對值分析按面積歸一化法計算, 以總脂肪酸的百分比形式呈現[14]。
1.5 數據處理
所有數據均采用SPSS統計軟件(19.0版, Chicago, IL, USA)的單因素方差分析和Duncan’s多重比較檢驗進行分析。使用Origin Pro 2017C(Northampton,MA, USA)軟件, 采用協方差矩陣法(Covariance matrix)對不同組織抗氧化酶活性數據進行主成分分析(Principal component analysis, PCA)分析, 解釋不同越冬時間對不同組織抗氧化能力的相對影響和差異; 同時采用協方差矩陣法(Covariance matrix)對肝胰臟、肌肉和脂肪組織脂肪酸數據進行主成分分析分析[15], 解釋不同越冬時間對上述組織脂肪酸組成的相對差異。采用相關矩陣法(Correlation matrix)對肝胰臟、肌肉和脂肪組織脂肪酸與酶活性數據進行主成分分析分析[16], 解釋不同越冬時間處理對肝胰臟、肌肉和脂肪組織中的抗氧化能力與脂肪酸組成的相對差異。其中, 各組織脂肪酸數據首先進行正(余)弦或對數轉換, 使之方差齊性, 以適應PCA分析對線性關系的要求[17], 再進行單因素方差分析(One-way ANOVA)和Duncan’s多重比較檢驗; 主成分分析中的因子提取使用, 按特征值大于0.5的標準確定, 為了使因子易于解釋和命名, 采用方差最大正交旋轉法對其進行旋轉處理。所有數據用平均值±標準差(mean±SD)的方式表示。P<0.05為差異顯著。柱狀圖采用Prism 7 軟件(Graph Pad Software Inc., San Diego, USA)進行繪制。
2 結果
2.1 不同越冬時間對草魚生物學性狀的影響
隨著越冬時間延長, 越冬草魚體重、肝胰臟重量、肥滿度、肝體比、臟體比、腸體比和腹腔脂肪指數等指標均呈現顯著下降的趨勢(P<0.05); 而腎指數和脾指數呈現顯著上升的趨勢(P>0.05; 表 1)。
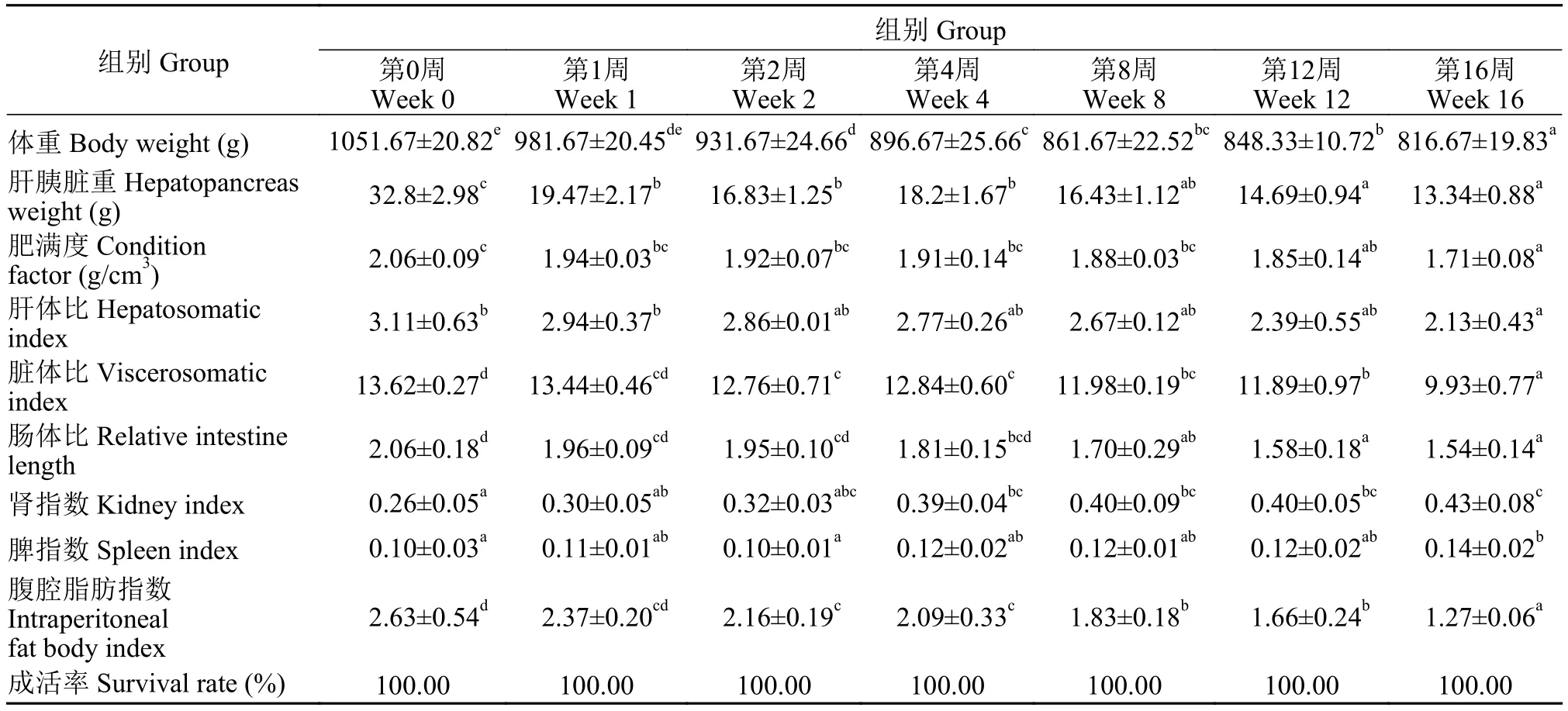
表1 不同越冬時間處理對草魚生物學性狀的影響Tab. 1 Effects of different overwintering time treatment on biometric parameters in grass carp (n=3)
2.2 不同越冬時間對草魚組織抗氧化指標的影響
如圖 1所示, 隨著越冬時間延長, 肝胰臟、肌肉、前腸、脂肪組織和血清中抗氧化能力相關指標均發生了顯著變化(圖 1A—E;P<0.05), 故而采用主成分分析法降低原有多個組織中抗氧化能力相關指標變量的維度, 用較少的獨立公因子反映原有變量的絕大部分信息。圖 2顯示了經PCA處理后形成的評分圖和載荷圖。載荷特征值如表 2所示, 只有特征值大于1.0才被視為數據差異的重要主成分(根據Kaiser規則)。前3個公因子的特征值均大于1.0, 表明這3個公因子對PCA解釋的累計方差有著重要貢獻。PC1和PC2一起解釋了98.96%的累計方差貢獻率(方差貢獻率分別為84.79%和14.17%),PC3只解釋0.96%方差貢獻率。因此, 3個公因子足以解釋不同越冬處理下, 不同組織中不同抗氧化能力相關指標的變化。在評分圖中, 不同顏色橢圓面積大小表示不同組織中抗氧化能力相關指標在不同越冬時間處理下的變化趨勢, 面積越大, 表示抗氧化能力相關指標在越冬時間處理下變化程度越大, 那么該組織在越冬時間內其抗氧化能力指標體系變化越劇烈。橢圓面積由大到小依次是: 脂肪組織>肝胰臟>肌肉>前腸>血清。顯示脂肪組織在越冬時間內其抗氧化能力指標體系變化程度最劇烈,間接體現脂肪組織在越冬期間所有氧化應激程度最大; 其次是肝胰臟和肌肉, 變化最小的是前腸和血清。而載荷圖顯示, SOD和CAT對總體抗氧化能力的貢獻中起主要影響(主成分載荷特征值絕對值大于0.5)。
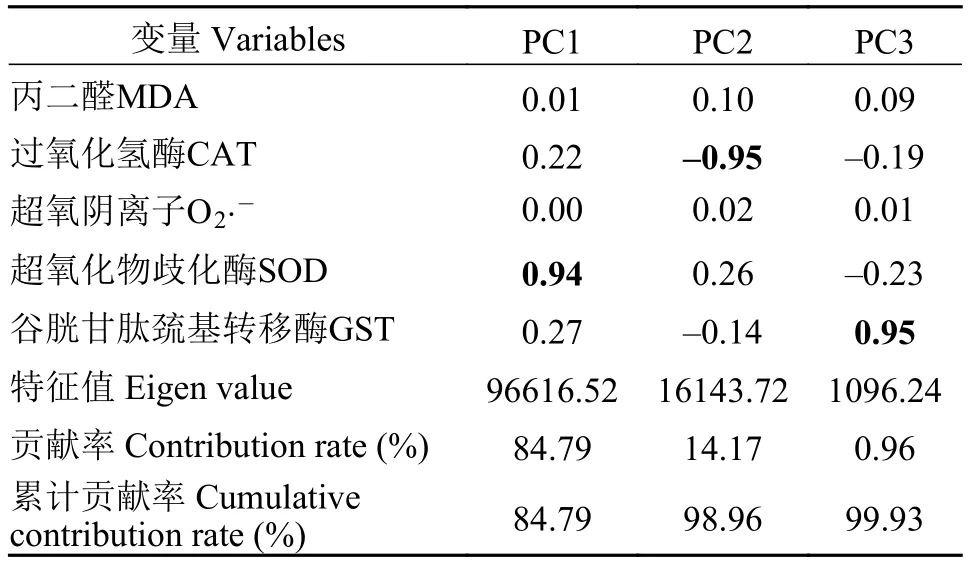
表2 協方差矩陣法下的顯著主成分載荷特征值分析Tab. 2 Eigen analysis of the covariance matrix loadings for significant principal components

圖1 不同越冬時間處理對草魚各組織抗氧化指標的影響Fig. 1 Effects of different overwintering time treatment on antioxidant indexes in tissues of grass carp (n=3)

圖2 通過分析不同越冬時間處理下草魚各組織中抗氧化能力, 根據選定的抗氧化酶活性含量變量(圖 1)生成PCA評分圖和載荷圖Fig. 2 PCA score plot and loading plot are generated based on selected antioxidant capacity variables (Fig. 1) obtained by analyzing the fatty acid composition in tissue and serum of grass carp under different overwintering time treatments
2.3 不同越冬時間對草魚肝胰臟、肌肉和脂肪組織脂肪酸組成的影響
根據上述結果, 不同越冬時間內肝胰臟、肌肉和脂肪組織中抗氧化體系變化最劇烈, 故而選定這3種組織進行脂肪酸比例測定。隨著越冬時間的增長, 肝胰臟、肌肉和脂肪組織中各類脂肪酸組成比例均產生了顯著差異(P<0.05; 表 3—5)。通過主成分分析, 圖 3中評分圖表示不同越冬時間處理下草魚肝胰臟、肌肉和脂肪組織中脂肪酸比例的變化趨勢, 各組織在不同越冬時間處理下, 均表現出了較大的差異性; 在肝胰臟中, 越冬第0、第1和第4周處理組在評分圖第一四象限, 越冬第2和第12周處理組在第二象限, 越冬第8和16周處理組在第三象限; 在肌肉中, 越冬第0、第1、第4和第8周處理組在評分圖第三四象限, 越冬第2周處理組在第一象限, 越冬第12和第16周處理組在第二象限; 脂肪組織中, 越冬第2周處理組在第一象限, 越冬第0、第8、第12和第16周處理組在評分圖第二三象限, 越冬第1和第4周處理組在評分圖第四象限。表 6中,主成分分析后, 對肝胰臟、肌肉和脂肪組織分析得出: 前3個公因子累計貢獻率均超過95%, 表明可以使用上述3個主因子(PC1、PC2和PC3)較好地代表18項脂肪酸變量指標。結合圖 3, 對肝胰臟、肌肉和脂肪組織載荷圖進行分析可得: 肝胰臟PUFA比例變化對總體脂肪酸比例變化產生主要影響(主成分載荷特征值大于0.5), 在越冬期間, PUFA比例變化呈現顯著上升(P<0.05); 肌肉C18:2n-6和C16:0比例變化對總體脂肪酸組成產生了主要影響, C18:2n-6比例變化在越冬期間呈現先顯著下降而后顯著上升, C16:0比例變化顯著下降(P<0.05); 腹腔脂肪PUFA、n-6PUFA、SFA和MUFA比例變化對總體脂肪酸比例產生了主要影響, 越冬脅迫期間, PUFA、n-6PUFA和MUFA比例變化呈現顯著下降后上升保持穩定, SFA比例變化呈現顯著上升后下降保持穩定(P<0.05)。

表6 協方差矩陣法下的顯著主成分載荷特征值分析Tab. 6 Eigen analysis of the covariance matrix loadings for significant principal components

圖3 通過分析不同越冬時間處理下草魚肝胰臟、肌肉和脂肪組織脂肪酸組成, 根據選定的脂肪酸變量(表 3—5)生成PCA評分圖和載荷圖Fig. 3 PCA score plot and loading plot are generated based on selected fatty acid variables (Tab. 3—5) obtained by analyzing the fatty acid composition in hepatopancreas, muscle and adipose tissue of grass carp under different overwintering time treatments
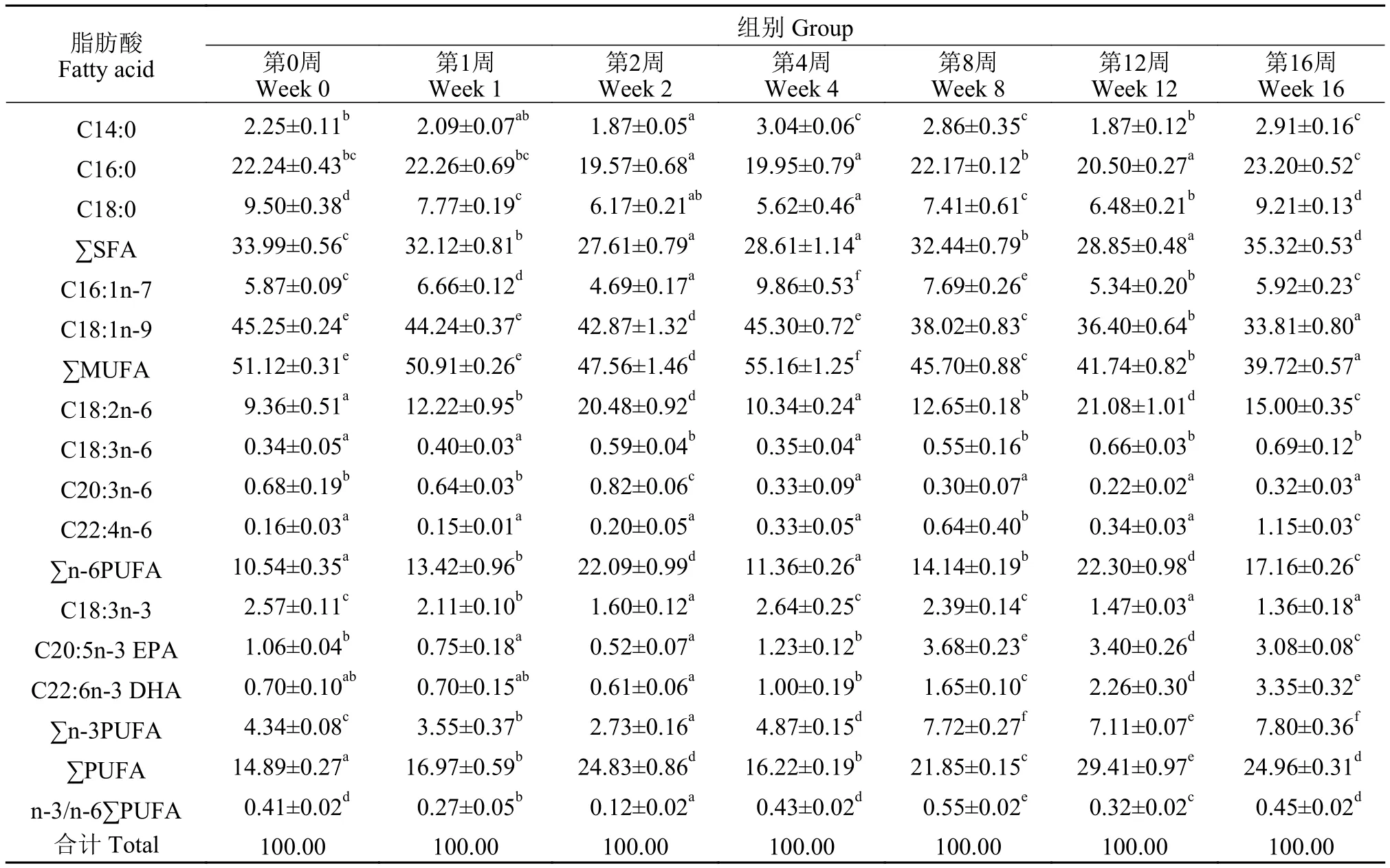
表3 不同越冬時間處理對草魚肝胰臟組織脂肪酸組成的影響Tab. 3 Fatty acid composition of hepatopancreas in grass carp under different overwintering time treatments (% total fatty acid; mean±SD;n=3)
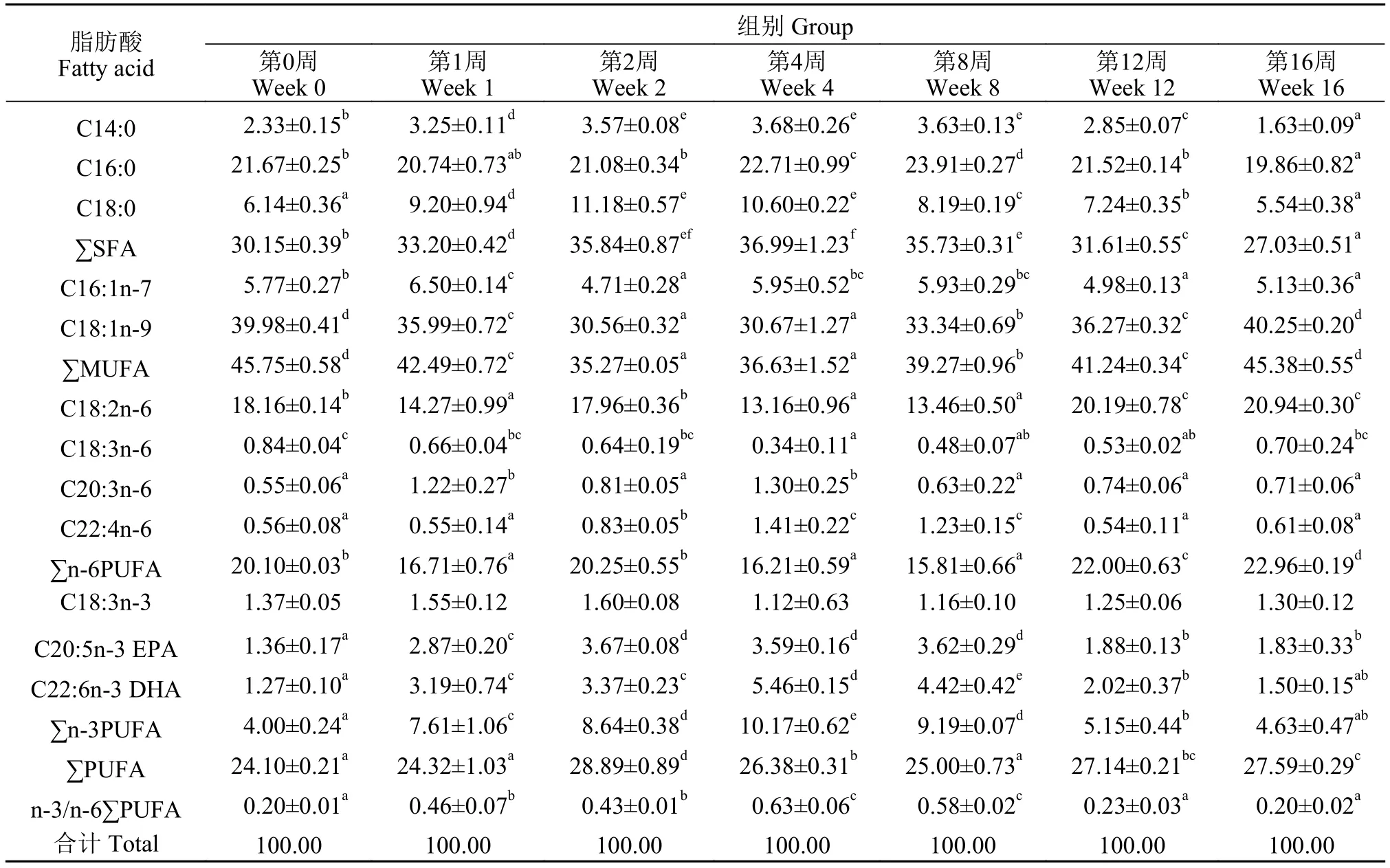
表4 不同越冬時間處理對草魚肌肉組織脂肪酸組成的影響Tab. 4 Fatty acid composition of muscle in grass carp under different overwintering time treatments (% total fatty acid; mean±SD; n=3)
2.4 不同越冬時間對草魚肝胰臟、肌肉和脂肪組織脂肪酸組成和抗氧化能力相關指標關聯性的影響
對越冬脅迫下, 草魚肝胰臟、肌肉和脂肪組織脂肪酸組成以及抗氧化能力的相關指標進行了相關矩陣法主成分分析之關聯性分析(圖 4)。載荷特征值如表 7可得: 當特征值大于1.0才會認為是數據差異的重要主成分(Kaiser規則)。由于前3個公因子特征值均大于1.0, 表明前3個公因子對PCA解釋累計方差貢獻率有著極其重要的貢獻。肝胰臟和肌肉PC1、PC2和PC3累計方差的貢獻率超過了80%, 脂肪組織累計方差貢獻率則超過了75%。但是載荷特征值表 7顯示, 沒有任何指標變量起到了主要影響(主成分載荷特征值絕對值大于0.5), 但是按高低排序, 發現其中最具有影響作用的3個變量,分別是肝胰臟中的MUFA、n-3PUFA和PUFA; 肌肉中的DHA、n-3PUFA和MUFA; 脂肪組織中的SFA、C16﹕1n-7和PUFA。圖 4中, 肝胰臟脂肪酸組成的比例和抗氧化能力指標位于得分圖的同一區域, 如CAT、SOD、GST和脂肪酸C18﹕2n-6、C18﹕3n-6、n-6PUFA、PUFA均在第二象限緊密聚集, MDA和脂肪酸C20﹕3n-6、C18﹕1n-9、MUFA均在第一象限緊密聚集, 表明存在正相關; 肌肉脂肪酸組成比例和抗氧化能力指標位于得分圖的同一區域, 如MDA、SOD、GST和脂肪酸C18﹕1n-9、MUFA均在第一象限緊密聚集, CAT和脂肪酸C16﹕0、C16﹕1n-7、C22﹕4n-6、DHA均在第二象限緊密聚集, 表明存在正相關; 脂肪組織脂肪酸組成比例和抗氧化能力指標位于得分圖的同一區域,如CAT、SOD、GST和脂肪酸C16﹕0、C18﹕0、C20﹕3n-6均在第四象限緊密聚集, MDA和脂肪酸SFA、MUFA、EPA均在第一象限緊密聚集, 表明存在正相關; 雖然這些變量對方差貢獻較小, 但從該分析中依然獲得了組織脂肪酸組成與抗氧化能力相關變量間較為積極的關聯性的結果。
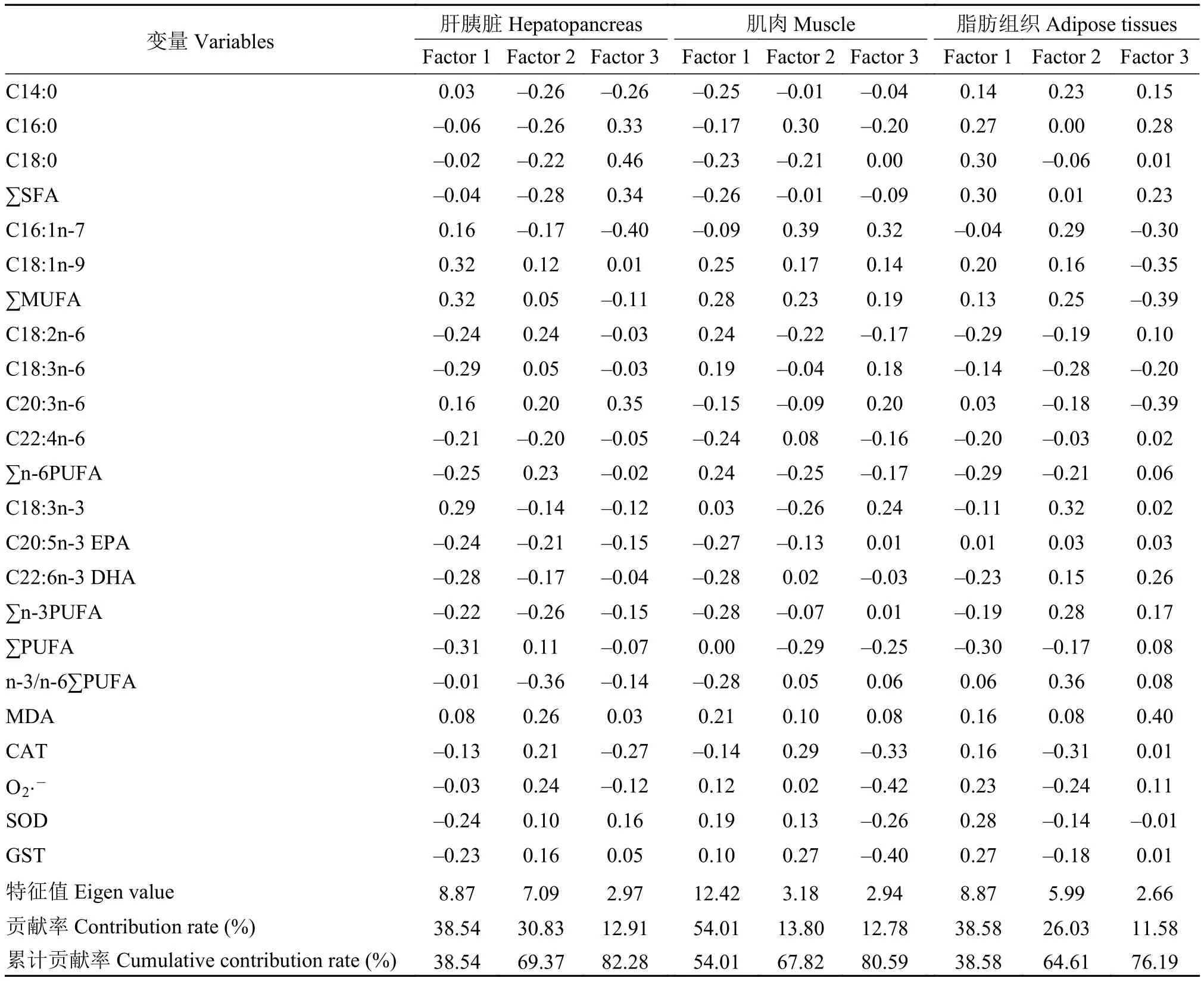
表7 相關矩陣法下的顯著主成分載荷特征值分析Tab. 7 Eigen analysis of the correlation matrix loadings for significant principal components

圖4 不同越冬時間處理對草魚肝胰臟、肌肉和脂肪組織脂肪酸組成及抗氧化能力的主成分分析Fig. 4 Principal component analysis (PCA) of fatty acid composition and antioxidant capacity variables in hepatopancreas,muscle, foregut, adipose tissue and serum of grass carp under different overwintering time treatments

表5 不同越冬時間處理對草魚腹腔脂肪組織脂肪酸組成的影響Tab. 5 Fatty acid composition of adipose tissue in grass carp under different overwintering time treatments (% total fatty acid; mean±SD;n=3)
3 討論
3.1 不同越冬時間對草魚生物學性狀的影響
魚類在自然界生長過程中, 經常面臨饑餓這一種自然生理脅迫。在饑餓脅迫影響下, 魚體往往只能依靠自身的營養物質的消耗來維持機體能量的代謝平衡[18]。生物學性狀是反映不同狀態下魚體營養儲備消耗情況最直接量化指標[19]。本研究結果表明, 在越冬脅迫下, 草魚的生物學性狀參數中,體重、肝胰臟重量、肥滿度、肝體比、臟體比、腸體比和腹腔脂肪指數均呈現顯著下降的趨勢, 但是腎指數和脾指數呈現顯著上升的趨勢; 相比較肝胰臟質量和腹腔脂肪指數, 則在第1和第2周就出現了顯著的下降。這表明在越冬期間, 肝胰臟和腹腔脂肪組織均在第一時間進行了能量動員, 供應魚體進行了能量消耗, 這也許是魚體為了適應氧化應激狀態下, 需要能量物質進行消耗進而來維持正常生理狀態的一種保護性手段[20]。
3.2 不同越冬時間對草魚組織抗氧化指標的影響
肝胰臟是魚類重要的器官, 具有多種代謝及分解功能[21]; 肌肉是魚類重要的運動組織[22], 前腸是重要的消化器官[23], 腹腔脂肪組織是作為草魚重要的儲能器官[24], 血清則作為一種組織液, 起維持機體內環境穩態的作用[25]。生物學性狀指標的變化,如: 肝胰臟質量、肝體比、腸體比和腹腔脂肪指數, 也表明了這些組織應對氧化應激所做出的適應性調整。在整個越冬脅迫期間, 對上述組織及血清抗氧化指標的測定后, 同時進行了PCA分析, 結果表明, 各個組織和血清在越冬脅迫期間所受的氧化脅迫的劇烈程度有所不同, 顯示: 脂肪組織>肝胰臟>肌肉>前腸>血清, 表明腹腔脂肪組織在越冬脅迫期間所受氧化應激的程度最大, 其次是肝胰臟和肌肉, 變化最小的則是前腸和血清。而SOD和CAT活性大小對總體抗氧化能力的貢獻中起主要影響[26]。內源性抗氧化防御系統是保護魚類組織和細胞免受氧化損傷的系統, 而SOD和CAT酶是該系統的主要反應元件之一。SOD可將細胞內的超氧陰離子轉化成H2O2, 同時CAT又將H2O2轉化成無毒無害的H2O以及O2, 進而降低組織內的氧化應激,維持細胞內的氧化還原平衡[27]。MDA水平的高低則間接反映了魚體遭受氧化自由基損傷后的嚴重性, 是反映魚體的細胞膜受到氧化破壞的重要指示物[28]。GST是組織中存在的較為豐富的抗氧化酶之一, 是抗氧化防御系統組成的關鍵部分[29]。腹腔脂肪組織作為草魚體內儲存能量物質最為豐富的器官[30], 在整個越冬脅迫期間自始至終保持著能量供應的狀態, 脂肪分解供能較其他組織活躍很多,脂肪酸的分解和氧化供能以及脂肪酸轉運也較其他組織活躍, 這可能就是草魚越冬期間脂肪組織受到較高氧化應激的原因之一。
3.3 不同越冬時間對草魚肝胰臟、肌肉和脂肪組織脂肪酸組成的影響
這里選擇越冬脅迫期間所受氧化應激程度最大的3個組織: 肝胰臟、肌肉和脂肪組織。在不同時間越冬脅迫處理下, 脂肪酸比例均有顯著不同。這反映出了不同時間越冬脅迫下, 脂質動員出現了組織特異性。PCA分析表明, 肝胰臟中PUFA比例變化, 肌肉中C18﹕2n-6和C16﹕0比例變化, 腹腔脂肪中PUFA、n-6PUFA、SFA和MUFA比例變化對總體脂肪酸比例產生了主要影響, 脂肪酸比例種類差異較為明顯[16,31]。脂肪組織中PUFA、n-6PUFA和MUFA比例顯著下降后上升保持穩定, SFA比例顯著上升后下降保持穩定; 肝胰臟PUFA比例顯著上升; 肌肉中C16﹕0和C18﹕2n-6比例顯著下降。尤其是功能性脂肪酸的代表之一PUFA, 在肝胰臟、肌肉和脂肪組織比例變化的趨勢更能體現越冬脅迫下, 組織應對越冬脅迫所表現的組織特異性, 脂肪組織中PUFA和n-6PUFA也可能進行了供能; 肝胰臟PUFA比例變化的上升, 表明魚體為了維持某些特殊生物學功能而間接保留PUFA; 肌肉PUFA比例的變化對總脂肪酸變化貢獻不大, 但C18﹕2n-6比例變化的下降可能預示著進行了供能。
3.4 不同越冬時間對草魚肝胰臟、肌肉和脂肪組織脂肪酸組成和抗氧化能力相關指標關聯性的影響
對肝胰臟、肌肉和脂肪組織中, 抗氧化能力的相關指標與脂肪酸比變化例間的相關性進行的分析結果表明, 脂肪酸比例變化與各組織抗氧化能力大小有著較強的相關性, 圖 5展示了脂肪酸組成在越冬脅迫下與機體抗氧化能力間關系的示意圖。草魚肝胰臟中抗氧化指標CAT、SOD、GST和脂肪酸變量C18﹕2n-6、C18﹕3n-6、n-6PUFA、PUFA有關聯作用, MDA和脂肪酸變量C20﹕3n-6、C18﹕1n-9、MUFA有關聯作用; 同時肝胰臟中脂肪酸變量MUFA、n-3PUFA和PUFA對關聯分析相關指標貢獻排名靠前, 表明肝胰臟中與氧化應激指標直接關聯脂肪酸變量為PUFA, 對機體損傷指標MDA直接關聯脂肪酸變量為MUFA。肌肉中抗氧化指標MDA、SOD、GST和脂肪酸變量C18﹕1n-9、MUFA有關聯作用, CAT和脂肪酸變量C16﹕0、C16﹕1n-7、C22﹕4n-6、DHA有關聯作用;同時肌肉中脂肪酸變量DHA、n-3PUFA和MUFA對關聯分析相關指標貢獻排名靠前, 表明肌肉中與氧化應激指標機體損傷指標MDA直接關聯脂肪酸變量為MUFA。脂肪組織中抗氧化指標CAT、SOD、GST和脂肪酸變量C16﹕0、C18﹕0、C20﹕3n-6有關聯作用, MDA和脂肪酸變量SFA、MUFA、EPA有關聯作用; 同時脂肪組織中脂肪酸變量SFA、C16﹕1n-7和PUFA對關聯分析相關指標貢獻排名靠前, 表明脂肪組織與氧化應激指標直接關聯脂肪酸變量為SFA中的C16﹕0, 對機體損傷指標MDA直接關聯脂肪酸為SFA。上述結果表明: 草魚腹腔脂肪組織中脂肪酸變量SFA在越冬脅迫期間供應能量的同時與氧化應激乃至機體損傷顯示主要正相關, 肌肉中脂肪酸變量PUFA、MUFA分別與氧化應激乃至機體損傷顯示主要正相關, 而肝胰臟中脂肪酸變量MUFA與氧化應激乃至機體損傷顯示主要正相關[32,33]。相關性分析為我們揭示了, 草魚在越冬脅迫期間, 受到氧化應激脅迫最大的3個組織與脂肪酸變量間產生的某種關聯, 為越冬脅迫期間草魚機體脂肪酸劇烈變化的同時, 將脂肪酸動員與氧化應激進而機體的損傷聯系在一起, 提供了一條新的思路和見解。

圖5 越冬脅迫下草魚組織中脂質過氧化與機體抗氧化能力間的關系示意圖Fig. 5 Relationship between lipid peroxidation and antioxidant capacity of grass carp under overwintering stress
4 結論
綜上所述, 草魚在越冬脅迫過程中, 生物學性狀變化的結果表明了草魚為了適應越冬脅迫下能量動員的需要采取了適應性調節, 同時, 我們證明了越冬期間所有氧化脅迫應激最大的3個組織分別是脂肪組織、肝胰臟和肌肉, 同時與越冬期間作為主要能量物質消耗的脂肪酸間進行了一種關聯性分析, 確定了幾種主要脂肪酸種類(肝胰臟MUFA, 肌肉PUFA、MUFA, 脂肪組織SFA)在越冬期間對氧化應激乃至機體損傷的直接相互聯系。本研究提供的基準信息, 可用于制定有效的越冬脅迫期之前的投喂策略, 同時在越冬脅迫期間和越冬脅迫后的恢復階段, 做出適當管理和精準的投喂決策, 以期改善草魚越冬脅迫后的存活率和生產的效率。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
