米仔蘭一年生幼莖的發育解剖學研究
2022-09-09 08:56:28呂佳峻朱栗瓊招禮軍楊鳳蘭陳惠昕龍孟元
熱帶農業科學 2022年8期
關鍵詞:植物
呂佳峻 朱栗瓊 招禮軍 楊鳳蘭 陳惠昕 龍孟元
(廣西大學林學院 廣西南寧 530004)
米仔蘭(Aglaia odorata)是楝科(Meliaceae)米仔蘭屬(Aglaia)的小喬木、灌木,產于廣東、廣西,花黃色且芳香似蘭,不僅是優良的觀賞樹種,還是改善血液循環、治療跌打損傷等疾病的傳統中藥材[1-3]。目前學者們對于米仔蘭的研究有扦插繁殖、葉綠體基因組序列、化合物的除草和抗癌作用等[2,4-6]、關于米仔蘭莖的發育解剖學研究罕見報道。對植物莖進行解剖研究可為選育、栽培、分類和鑒定等提供依據[7-8],還可揭示植物的生長發育規律[9],其中導管分子形態特征對樹體生長起著重要指示作用[10]。另外,學者們研究植物莖的解剖結構特征與植物抗旱、抗寒和抗蟲害等抗逆性的相關性[11-14]。在莖的發育解剖學研究中,相關學者指出,莖發育過程一般包括原分生組織、初生分生組織、初生生長和次生生長4個階段[15-16],少部分植物有三生生長[17]。莖的初生結構有表皮、皮層和維管束,不同生長時期各類細胞的比例不同,隨著生長發育植物莖的各項功能逐漸完善[18-19]。自然界中雙子葉植物約有20萬個物種,分布廣泛,具有重要的經濟價值[20]。近年來關于莖的發育解剖學研究多見于單子葉植物[21-23],而雙子葉植物較少。本研究擬從米仔蘭一年生莖著手,按照發育順序,研究第1~6 節莖的解剖結構和導管分子特征,探究米仔蘭幼莖的發育規律及其與結構和功能相適應的方式,為雙子葉植物幼莖的發育解剖學研究提供理論依據。
1 材料與方法
1.1 材料
實驗材料于2021 年8 月采自廣西大學校園內生長健壯的5 株米仔蘭,植株為全光照下、株間相隔至少10 m。
1.2 方法
1.2.1 樣品采集與處理每株采集東西南北4 個方向,每個方向采集3 條一年生枝條,總共60 條。將頂芽下第1 節定為莖的第1 節,依次往下為莖的第2、3、4、5、6 節。于節處截斷,相同位置的歸為一類,分別置于6 個裝有70% FAA 液(70%酒精∶冰醋酸∶38%甲醛=90∶5∶5)的容器中固定48 h,待用。
1.2.2 制片
1.2.2.1 莖的石蠟切片各節隨機取30 段樣品的中段(長約1 cm),采用常規石蠟切片技術與叔丁醇脫水法相結合[24-25]。軟化:使用冰醋酸∶過氧化氫=1 ∶1(V/V)混合溶液浸泡樣品并抽真空0.5 h,處理3 h。脫水、透明:采用50%、70%、85%、95%叔丁醇依次各處理1.5 h,純叔丁醇處理0.5 h,重復2 次。浸蠟:用50%石蠟(叔丁醇∶石蠟=1∶1,V/V)、75%石蠟(叔丁醇∶石蠟=3∶1,V/V)將脫水后的材料依次處理6 h;在純石蠟溶液中處理12 h,期間每隔4 h 更換1 次石蠟溶液,共更換3 次。經過包埋、切片、展片、晾片后,進行以下操作:TO 脫蠟→梯度酒精復水→番紅染色→梯度酒精脫水→TO 透明;用中性樹膠進行封片,每段切片至少10 片;在光學顯微鏡下觀察并拍照記錄,在圖像測量軟件Image J 中進行數據測量,每一指標隨機選取30 個重復。觀察和測定指標如下:
表皮率=表皮橫切面積/莖段橫切面積×100%
皮層率=皮層橫切面積/莖段橫切面積×100%
初生韌皮部率=韌皮部橫切面積/莖段橫切面積×100%
初生木質部率=初生木質部橫切面積/莖段橫切面積×100%
髓率=髓橫切面積/莖段橫切面積×100%
1.2.2.2 導管離析制片按上述方法各節取6 段樣品,除去表皮,浸沒在冰醋酸∶30%過氧化氫=1:1(V/V)的混合離析液中,放入70℃烘箱中離析24 h;使用蒸餾水沖洗干凈、番紅染色,去浮色后,置于裝有蒸餾水的玻璃瓶中搖散;用滴管吸取樣液制成臨時玻片,在光學顯微鏡下觀察。每節制作10 個臨時裝片,每個取3 個視野,每個視野選取1 個典型的導管分子,總共30 個,并拍照記錄。在圖像測量軟件Image J 中測量每個導管分子的長度、寬度、尾端長度和端壁傾斜度,其中尾端長度和端壁傾斜度測量導管兩端2 個數據,取平均值。
1.2.3 數據處理與分析使用Excel 2016 對米仔蘭幼莖橫切結構以及導管分子特征參數等數據進行整理和作圖,采用SPSS 22.0 進行數據分析,采用LSD 作多重比較。
2 結果與分析
2.1 米仔蘭一年生幼莖橫切結構隨發育的變化
米仔蘭一年生幼莖的橫切結構如圖1-I 所示,其橫切面不規則,由外到內依次是:表皮、皮層、初生韌皮部、束中形成層、初生木質部和髓,共6 部分。由表1 可知,隨著莖的發育,其直徑不斷變粗,各橫切結構厚度均呈現上升趨勢。表皮由1 層排列緊密的細胞組成,在第1~2 節莖中多數為不規則的近圓形(圖1-C、1-G),發育至第3~4 節莖為近正方形(圖1-K、1-O),第5~6 節莖則是長方形狀的扁平細胞(圖1-S、1-W),在第1~6 節莖表皮細胞最外部均形成了角質膜。皮層從第1~6 節增厚了16.46 μm(表1),靠近表皮處的5~8 層細胞為厚角組織,細胞較小、排列緊密,且含有葉綠體,可進行光合作用;靠近韌皮部6~10 層細胞為薄壁組織,細胞較大、排列疏松,因此間隙發達,便于儲藏空氣。在發育過程中這兩部分組織的細胞層數增加不顯著,但細胞體積不斷增大,并且靠近韌皮部的薄壁細胞形狀由第1~2 節莖的近圓形,不斷被擠壓成不規則的扁平狀(圖1-B、1-F、1-J、1-N、1-R、1-V)。
米仔蘭一年生幼莖為外韌維管束,初生韌皮部在外,初生木質部在內,具有束中形成層。初生韌皮部第6 節厚度是第1 節的1.54 倍,初生韌皮部率增加了2%;初生木質部厚度第6 節是第1節的2.25 倍,初生木質部率提高了6.20%(表1)。解剖發現,各維管束形狀不同、間隔不明顯,髓射線較窄。隨著發育,維管束的形狀逐漸不易區分(圖1-B、1-F、1-J、1-N、1-R、1-V)。木質部中有木射線和導管,莖橫切面在有棱或凹槽處的初生木質部明顯增厚,但發育至第5 節莖后,增厚的特征開始消失(圖1-A、1-E、1-I、1-M、1-Q、1-U)。
髓位于莖中央,由較大的薄壁細胞組成,其直徑逐漸增大,第6 節是第1 節的1.46 倍;髓率先增后減,但最終增加了1.80%(圖1-A、1-E、1-I、1-M、1-Q、1-U,表1)。
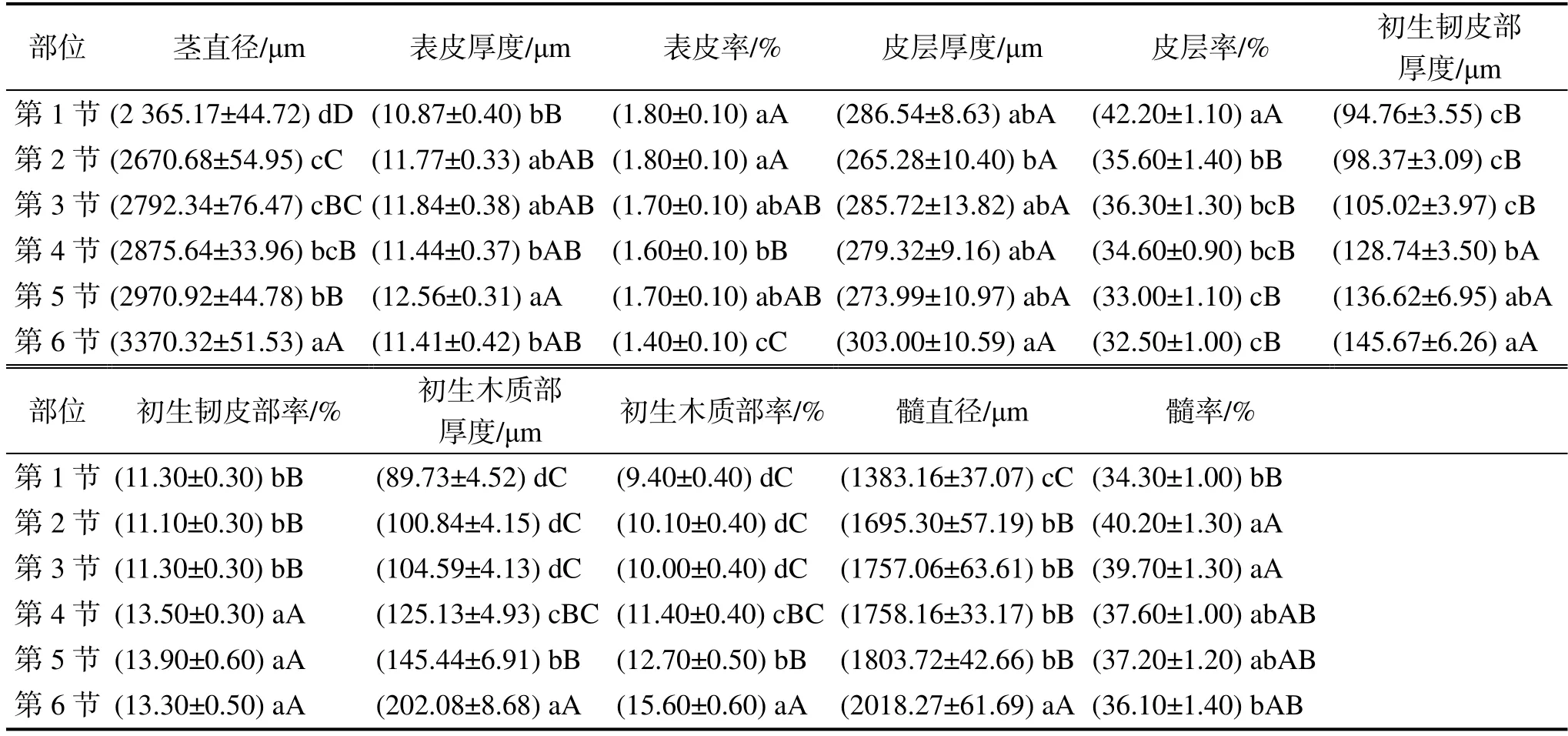
表1 米仔蘭一年生幼莖橫切結構多重比較
2.2 米仔蘭一年生幼莖導管分子隨發育的變化
按次生壁加厚方式分類,米仔蘭一年生幼莖導管分子類型有:環紋導管、梯紋導管、網紋導管和孔紋導管(圖2),其中環紋導管極少。各類導管的比例隨著發育,從第1~6 節莖,呈現由以梯紋導管為主向以網紋導管、孔紋導管為主的變化(圖3)。導管分子具尾情況有:兩端具尾、一端具尾一端無尾以及兩端無尾,其中兩端無尾在第3 節莖開始出現(如圖2、4)。導管分子尾端長度先增后減,從第 1~5 節莖逐漸上升,在第1~2 節莖時增加顯著,但第2~5 節莖的上升趨勢均不顯著,至第5~6 節莖發育時期則開始下降,且第6 節莖導管分子的尾端長度與第1 節差異不顯著(表2)。
由表2 可知,在發育過程中,導管分子的長度縮短,寬度增加,二者變化趨勢相反。第1~5節莖期間導管長度縮短不明顯,彼此差異不顯著;發育至第6 節莖時長度顯著下降,與先前時期導管分子的長度呈極顯著差異。導管寬度變化情況與長度類似,在第1~3 節莖寬度加大不明顯,差異不顯著;從第4 節莖開始呈顯著加大,發育至第5~6 節莖的導管寬度與第1~4 節莖均呈極顯著差異。
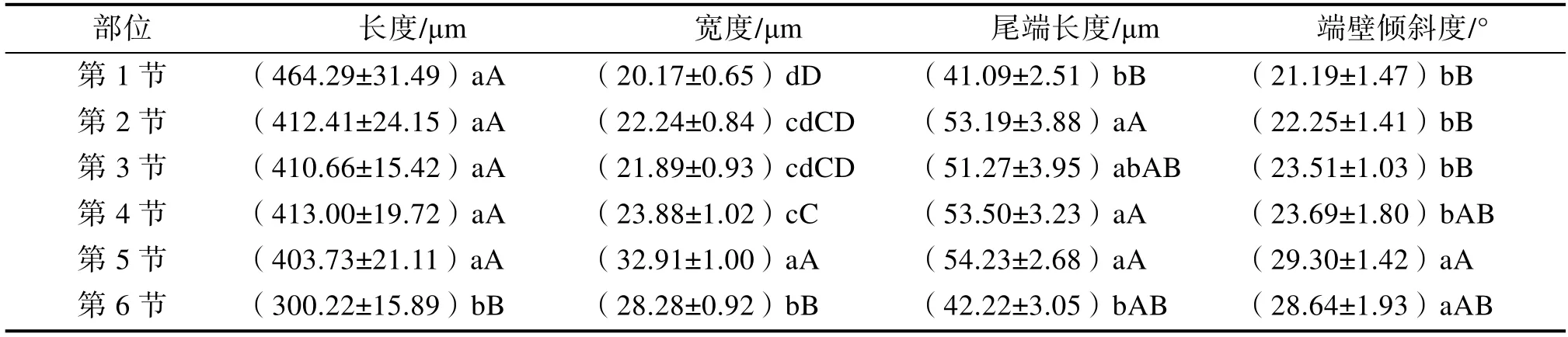
表2 米仔蘭一年生幼莖導管分子特征參數多重比較
隨著發育,導管分子的端壁傾斜度不斷增大,但第1~4 節莖傾斜度增加趨勢不顯著,發育至第5 節時傾斜度顯著上升。
3 討論與結論
3.1 米仔蘭一年生幼莖橫切面結構組成及發育規律
米仔蘭一年生幼莖橫切面形狀不規則,結構與一般雙子葉植物相同[26]。在米仔蘭第1~6 節莖的發育中表皮最外部均形成了角質膜,保護作用增強[27]。
本研究發現,米仔蘭幼莖皮層厚度以及薄壁細胞體積逐漸增大,說明隨著發育,位于皮層的薄壁細胞應對水分降低的能力不斷提高[28],雖然皮層率呈現下降的趨勢,但占比仍然在30%以上,與張敏等[29]對朝鮮越桔的研究結果類似。從第1~6 節莖,皮層薄壁細胞的形狀逐漸不規則,部分被擠壓成扁平狀,與陳存瑞[30]對牡丹的研究結果一致。因此,米仔蘭皮層發育規律與其它木本香花植物相同。
米仔蘭幼莖維管束為外韌型,但髓射線較窄、束狀外形不突顯,與一般雙子葉植物不同。初生韌皮部是植物有機物質的運送渠道[31],初生木質部是植物水分運輸的關鍵[32]。隨著發育,其初生韌皮部和初生木質部占比不斷提高,輸導能力逐漸增強。第1~5 節莖,初生韌皮部率大于初生木質部率,第6 節莖則相反,說明在第5~6 節莖發育時期,束中形成層的分化重心由初生韌皮部轉向初生木質部,幼莖木質化速度進一步提高。
髓中的薄壁細胞體積逐漸增大,是米仔蘭幼莖水分儲存能力提高的表現,同時是莖直徑加大的原因之一[33-34]。
3.2 米仔蘭導管分子發育規律及其系統學意義
導管分子特征有:次生壁加厚方式、寬度、長度、具尾類型、尾端長度以及端壁傾斜度等[35]。本研究發現,米仔蘭一年生幼莖導管分子的各項特征在個體發育過程中的演化規律不同。
3.2.1 導管類型導管分子次生壁加厚方式有5種,按照進化順序依次是環紋導管、螺紋導管、梯紋導管、網紋導管和孔紋導管[36],次生壁木質化加厚程度增加,管腔加大,輸導效率增強。環紋和螺紋導管出現在莖發育早期,梯紋、網紋和孔紋導管出現在莖發育較成熟時期[37-38]。米仔蘭一年生幼莖發育后期,網紋和孔紋是次生壁加厚的主要方式。導管寬度越大,輸導效率越高,但寬度加大也更容易栓塞化[39-40],降低導管分子的長度可以抵御栓塞的形成[41],同時可增強莖的機械支持力[42]。本研究中,導管長度隨發育縮短,寬度則增加。
3.2.2 導管具尾及端壁傾斜狀況導管具尾類型有:兩端具尾、一端具尾一端無尾、兩端無尾[43]。本研究中,從發育初期到發育后期,類型由兩端具尾、一端具尾一端無尾2 種類型增至3 種類型,且前2 種具尾類型的比例提高。導管無尾較具尾進化,但有學者認為,尾端的形成以及伸長有利于提高水分運輸效率和速率,即原始特征更利于不良環境[44];也有研究指出,同種植物導管分子的尾端長度不受環境因素的影響[45]。米仔蘭導管分子的尾端長度先增后減,發育初期和后期的尾端長度最小。
導管分子端壁傾斜度的變化是導管分子的演化特征之一[46]。本研究發現,從第1~6 節莖,導管分子端壁傾斜度逐漸增大,說明米仔蘭莖導管分子的端壁傾斜度特征較原始[47]。有學者認為,端壁傾斜度增大,上下兩個導管分子接觸的面積加大,輸導效率越高[44];另有學者持不同意見,認為端壁傾斜度的增加會導致運輸阻力上升,輸導效率降低[38]。因此,筆者認為,不同的植物類型結果不同,要視具體情況而定,可通過測定導水率得到結論。
綜上所述,米仔蘭一年生幼莖的發育結構呈現一定規律,與大部分雙子葉植物幼莖相似,同時結構特征與其功能性適應,保障了植株正常生長的需要。本研究從發育解剖學的角度研究米仔蘭幼莖,比較了米仔蘭與一般雙子葉植物幼莖發育的異同,但關于楝科植物解剖學的研究鮮有,缺乏與同科植物的比較。因此,需要對其它楝科植物的幼莖進行發育解剖研究,更進一步地對其進行比較和分析,以得出更加準確的結論。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
