谷子幾丁質酶基因家族鑒定及在白發病菌脅迫下的表達分析
2022-09-09 09:17:28韓海麗董夢迪馬芳芳韓淵懷韓彥卿
激光生物學報 2022年4期
關鍵詞:分析
韓海麗,王 鶴,董夢迪,馬芳芳, 韓淵懷,韓彥卿*
(1. 山西農業大學 a. 植物保護學院;b. 創新創業學院;c. 農學院,太谷 030801;2. 雜糧種質創新與分子育種山西省重點實驗室,太原 030031)
幾丁質酶是一類與植物病害相關的蛋白質[1-2],在高等植物生長發育過程和抗病過程中起著重要的作用[3-5]。Tabei等[3]將水稻的幾丁質酶基因導入黃瓜后,轉基因黃瓜對灰霉病的抗性顯著提高。Yamamoto等[4]將水稻的Ⅰ類幾丁質酶基因導入葡萄中提高了葡萄對白粉病的抗性。近年來,幾丁質酶基因已經被廣泛地轉入到不同作物種中,如水稻[5-6]、棉花[7]、大豆[8-9]、花生[10]、煙草[11-12]等,以提高植物的抗病性。關于植物幾丁質酶的研究比較常見,而且在真菌抗病研究中也有相關報道[13]。該基因最早是在菜豆中研究的[14],相關研究人員將菜豆幾丁質酶載入油菜、煙草中,發現菜豆幾丁質酶基因表達上升,死苗率下降,發病率顯著降低[15]。Rohini等[16]將煙草幾丁質酶基因導入花生中,發現幾丁質酶的基因表達量顯著上升,花生褐斑病的發病率顯著下降。2002年,Kishimoto等[17]將水稻幾丁質酶基因RCC2導入煙草中,發現其對灰霉病的抗性顯著提高。王果萍等[18]通過對西瓜植株進行接菌,并對轉基因西瓜進行分析,發現幾丁質酶基因顯著表達,且對西瓜枯萎病的發生有顯著的抑制作用。劉偉華等[19]通過對小麥的愈傷組織進行接菌,并對轉基因植株進行分析,發現幾丁質酶基因成功表達,與對照植株相比,幾丁質酶活性顯著增強,對白粉病的發生起到了很強的抑制作用。
幾丁質酶根據酶活結構域差異分為糖苷水解酶家族 18(glycoside hydroase family 18,GH18)和糖苷水解酶家族 19(glycoside hydroase family 19,GH19)2個亞家族。GH18在細菌、真菌、植物、哺乳動物、昆蟲和病毒中廣泛存在,而GH19亞家族僅出現在植物和鏈霉菌屬中,且其成員具有幾丁質結合域(chitin-bind domain,CBD)結構,可與幾丁質結合,增強植物體的抵抗能力[20-22]。
谷子是我國北方特色雜糧作物[23],不僅耐脊薄、耐貯藏、適應性強、水分利用率高[24],而且營養豐富、可糧草兼用[25],具有重要的經濟價值,也是禾本科作物抗逆基因挖掘的模式植物[26-28]。但隨著谷子品種的單一化種植,谷子白發病逐漸成為其主要病害之一。該病害的發生嚴重影響了谷子的生長發育,對谷子產量和品質構成了極大的威脅[29]。本研究通過谷子響應白發病菌侵染的轉錄組數據分析獲得了30 個幾丁質酶基因,并對谷子抗病相關幾丁質酶家族基因的基本信息、理化性質、染色體定位、基因結構、系統發育樹、蛋白基序等進行分析,擬明確幾丁質酶基因的基本特征和潛在功能,研究結果可為幾丁質酶基因的深入抗病機制解析及分子育種提供一定的研究基礎。
1 材料與方法
1.1 谷子幾丁質酶基因染色體定位、基因結構和保守結構域分析
首先從谷子的基因庫中獲得它們的染色體位置信息,接著利用TBtools軟件分析30個幾丁質酶基因的染色體位置[30],隨后對其進行串聯重復分析。判斷串聯重復的原則為:1)相鄰基因間的距離小于100 kb ;2)基因間相識度大于70%[31]。利用MEGA 7.0軟件對30個幾丁質酶基因的保守基序及保守結構域進行分析,得出Motif示意圖,進而用TBtools軟件對其進行可視化作圖。
1.2 谷子幾丁質酶基因家族結構及系統發育分析
利用 MEGA 7.0對30個幾丁質酶基因的蛋白序列進行比對,進而構建進化樹,校驗參數Bootstrap設置為1 000,其余參數為默認值。
1.3 谷子幾丁質酶基因家族成員蛋白理化性質分析及亞細胞定位
結合網站phytozome V12.1(https://phytozome.jgi.doe.gov/pz/portal.html V12.1),從在線公共網站上下載30個幾丁質酶基因的基本信息,再利用Expasy在線網站(https://www.genscript.com/psort.htm)對幾丁質酶基因家族成員進行理化性質分析。
1.4 谷子幾丁質酶基因表達模式分析
本課題組前期以抗病品種JG42及感病品種JG21為試驗材料,通過RNA的提取及公司測序后,我們獲得了12、24和48 h的轉錄組數據。經分析發現,幾丁質酶基因在抗感谷子品種中表達差異顯著。從谷子基因數據庫下載30個幾丁質酶基因的數據,用TBtools軟件繪制基因表達熱圖。
1.5 谷子幾丁質酶基因在干旱脅迫下的表達分析
選擇抗旱谷子(AN04)和干旱敏感谷子(豫谷1號)為材料,以不同光照條件(早:中等光照;中:強光照;晚:弱光照)為試驗組,正常未脅迫植株作為對照組,取其葉片進行RNA的提取,送公司進行二代測序后,利用TBtools軟件繪制基因表達熱圖。
1.6 差異表達基因的實時熒光定量PCR分析
分別對感病品種JG21及抗病品種JG42 兩葉一心時期的谷子進行孢子囊懸浮液噴灑接菌,接菌后12、24、48 h對抗感品種的葉片進行取樣,提取總RNA,反轉錄合成cDNA后進行實時熒光定量PCR(quantitative real-time PCR,qRT-PCR)驗證基因表達水平。采用Primer Premier設計基因特異引物(表1)。使用TaKaRa實時熒光定量試劑盒,以谷子β-Actin(Seita.3G265400.1)為內參基因,反應體系為10 μL,包括5 μL 2×PCR混合酶、1 μL 上下游引物混合液(10 μmol/L)、1 μL cDNA、3 μL無菌水。PCR程序為 :95℃ 30 s;95℃ 5 s,60℃ 30 s,39個循環 ;95℃ 1 min ;60℃ 30 s。試驗設3次重復,計算谷子幾丁質酶基因在白發病侵染下的相對表達量[32]。

表1 本試驗所用PCR擴增引物Tab. 1 Primers used in this study
2 結果與分析
2.1 谷子幾丁質酶基因家族的基因定位分析
通過染色體定位分析發現,谷子的30個幾丁質酶基因定位在除6號染色體外的其余8條染色體上(圖1)。其中,7號染色體上定位到的基因最多,為8個,其次是5號染色體,共定位到6個基因。1號染色體和3號染色體上的幾丁質酶基因最少,僅1個。2號、4號、8號和9號染色體上定位到的基因分別有4、2、3和4個,而其中Seita.J0579300定位在尚未組裝完整的scaffold上。進一步分析發現,在5號、7號、8號、9號染色體上,10個串聯重復涉及到14個基因(紅色矩形方框表示),如Seita.9G263100和Seita.9G263200屬于一個串聯重復,可推測所有形成的幾丁質酶基因串聯重復屬于同一亞家族。
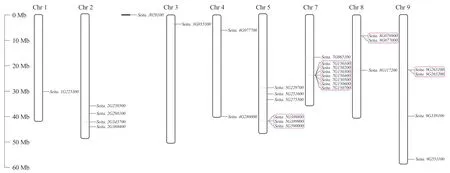
圖1 谷子幾丁質酶基因的染色體定位Fig. 1 Chromosome mapping of chitinase genes in foxtail millet
2.2 谷子幾丁質酶系統進化分析
為明確幾丁質酶蛋白基因的進化關系,本文利用MEGA 7.0軟件對查找到的30個幾丁質酶基因構建系統發育樹。根據幾丁質酶基因的功能可將其分為3大亞類(圖2)。第一大亞類含有2 個基因,第二大亞類含有11個基因,第三大亞類含有15個基因。第一大亞類具有相同的結構域,其2個基因都含有GRAS結構域,為赤霉素信號傳導的主要參與者,修飾或者結合小分子物質,能夠參與調節植物生長發育的各個方面;第二大類的11個基因主要為GH18,具有GH18的保守結構域;第三大亞類的15個基因為GH19,GH19家族幾丁質酶主要存在于植物中,作為真菌細胞壁的主要成分參與植物抗逆等過程[33]。
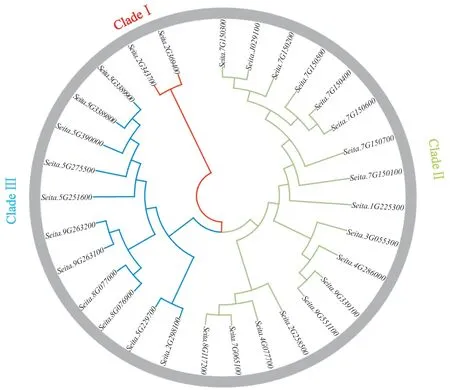
圖2 谷子幾丁質酶基因系統進化樹Fig. 2 Phylogenetic tree of the chitinase gene family of foxtail millet
2.3 谷子幾丁質酶基因蛋白理化性質分析
通過理化性質分析發現,該基因家族的氨基酸等電點(pI)為4.28(Seita.5G275500)~9.86(Seita.4G286000),相對分子質量為24 369.27(Seita.J0579300)~64 469.26 kD(Seita.2G343700)。蛋白質的不穩定指數(instability index,II)為19.17(Seita.7G150200)~50.91(Seita.2G298100),其中有12個幾丁質酶基因不穩定指數大于40,屬于不穩定蛋白。脂肪系數(aliphatic index)為53.80(Seita.4G286000)~84.49(Seita.5G229700),平均為72.71,說明它們的熱穩定性較為一致。依據平均疏水指數介于-0.5~0.5的為兩性蛋白的原則,這些幾丁質酶基因都為兩性蛋白,其中Seita.2G298100最大(0.250),Seita.2G343700最小(-0.482)(表2)。
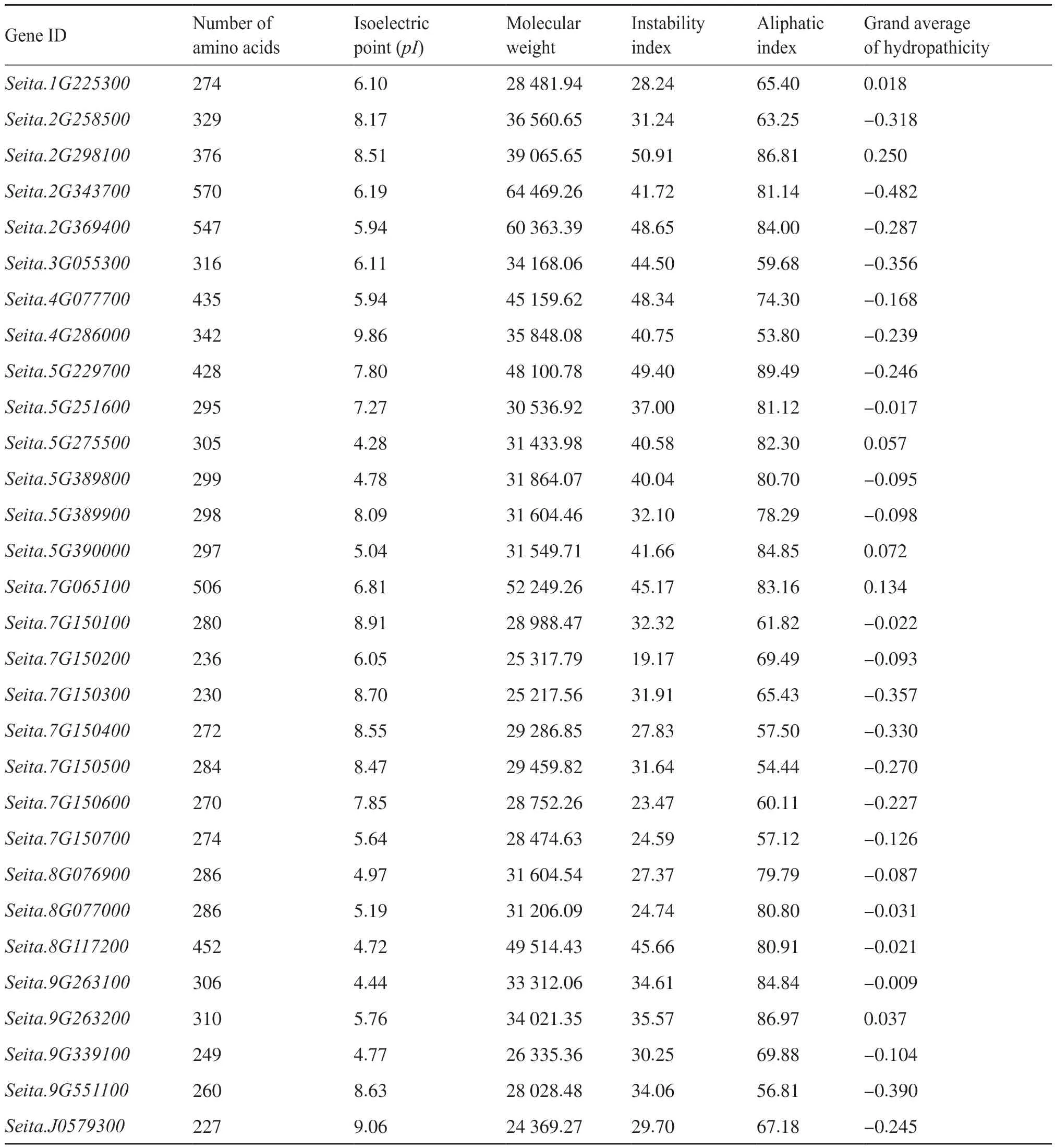
表2 谷子幾丁質酶蛋白基本理化性質Tab. 2 Basic physicochemical properties of chitinase in foxtail millet
2.4 谷子幾丁質酶蛋白二級結構預測和亞細胞定位
對30個谷子幾丁質酶蛋白的二級結構進行預測,發現該家族含有α-螺旋、延伸鏈、β-折疊和無規則卷曲。其中,延伸鏈占7%~19%,β-折疊在8% 左右,無規則卷曲和α-螺旋占二級結構總量的20%~40%。亞細胞定位預測表明,谷子幾丁質酶基因全部定位在細胞外(extracelluar),屬于分泌蛋白(表3),其所有基因表達蛋白質均在細胞外發揮作用。
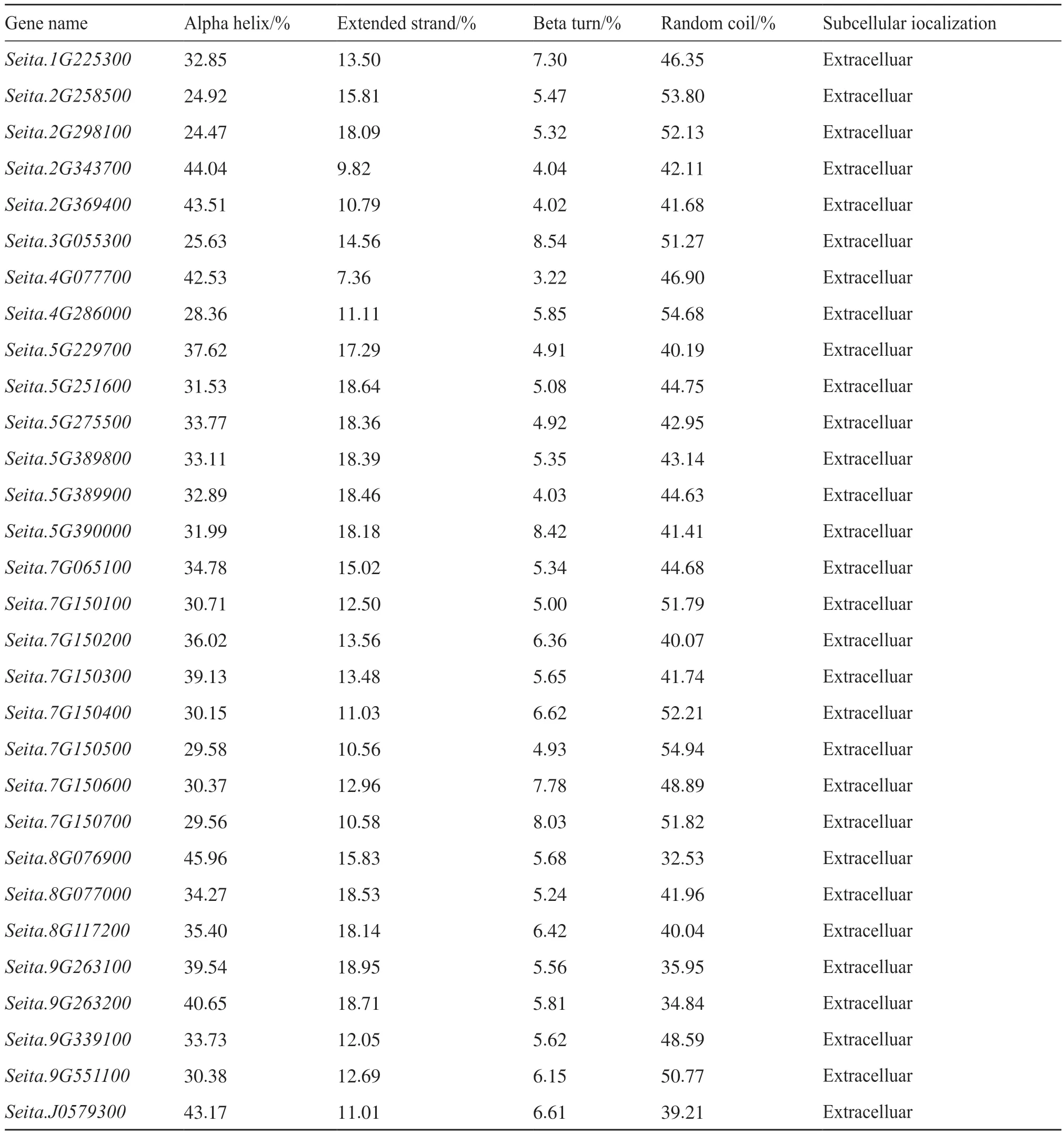
表3 谷子幾丁質酶蛋白二級結構預測和亞細胞定位Tab. 3 Secondary structure prediction and subcellular localization of chitinase foxtail millet
2.5 谷子幾丁質酶基因結構和保守結構域分析
對30個幾丁質酶基因家族基因結構分析和蛋白基序分析發現:它們含有10類蛋白基序(Motif 1~Motif 10),長度分別在15~50 個氨基酸之間(表4),其中,Seita.5G229700沒有鑒定到保守的Motif結構,Seita.2G369400只含有Motif 10 ;Motif 1、Motif 3 和 Motif 4 在 14 個蛋白中均被鑒定到 ;含有Motif 9 的蛋白最多,共19個,這19個蛋白被聚類在一起,說明這些蛋白可能發揮類似的功能。其次是Motif 2,在18個蛋白中被鑒定到。此外,Motif 8在11個蛋白中被鑒定到;Motif 7在9個蛋白中被鑒定到,且它們聚集在一起,推測具有相似的功能。Motif 5 和 Motif 6 分別在 7 個和 5 個蛋白中被鑒定到(圖3)。基因結構分析發現,30個幾丁質酶基因都含有上下游調控區(UTR),所有成員均包含外顯子、內含子,但在具體的數量和位置上存在差異(圖3)。
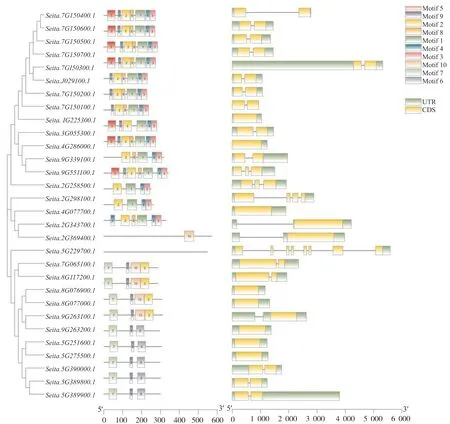
圖3 谷子幾丁質酶基因結構和保守結構域分析Fig. 3 Chitinase gene structure and conserved domain in foxtail millet

表4 谷子幾丁質酶基因家族成員10個蛋白基序分析Tab. 4 Protein motif analysis of ten members of chitinase gene family in foxtail millet
2.6 谷子幾丁質酶基因參與谷子逆境脅迫響應
對抗旱和干旱敏感兩個谷子品種的轉錄組數據進行分析,由圖4 可知,多數幾丁質酶基因在抗旱和敏感谷子品種中未見顯著表達差異。 其中,5個幾丁質酶在抗旱和敏感谷子品種中基因表達在不同時期存在差異。Seita.7G150200在干旱敏感品種,中等光照(早)中上調表達,而在強光照(午)、弱光照(晚)下調表達或幾乎不表達;Seita.7G150400和Seita.7G150500及Seita.7G150700有著相同的表達模式,在抗旱品種,中等光照(早)中上調表達,強光照(午)下調表達;Seita.5G390000和Seita.7G065100有著相同的表達模式,在干旱敏感品種弱光照(晚)中上調表達,而在中等光照(早)、強光照(午)中不表達或下調表達。這說明部分幾丁質酶的基因表達受到光照和品種的影響,在抗旱和干旱敏感的表達存在明顯差異(圖4)。
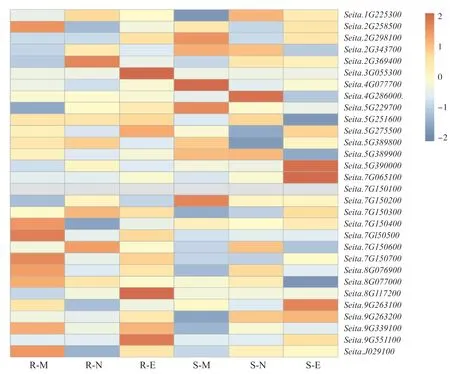
圖4 谷子幾丁質酶基因在干旱脅迫下的表達分析Fig. 4 Relative expression patterns of chitinase genes under drought stress in foxtail millet
2.7 谷子幾丁質酶基因響應白發病菌侵染的表達分析
為明確谷子幾丁質酶應對白發病菌侵染的響應,我們分析了抗感谷子在接種白發菌12、24和48 h后的基因表達模式。由熱圖可以看出:在抗病品種中,除Seita.2G369400在12 h表達明顯下調外,其他幾丁質酶基因大部分在抗病品種中12、24和48 h這3個時間段中均上調表達;在感病品種中,除了Seita.9G263100、Seita.8G117200和Seita.7G150400這3個基因在12 h表達上調外,其余大部分幾丁質酶基因在感病品種中3個時間段均表現為下調表達(圖5)。由此可推測,這些幾丁質酶基因在響應白發病菌侵染過程中發揮了重要的抗病調控作用。
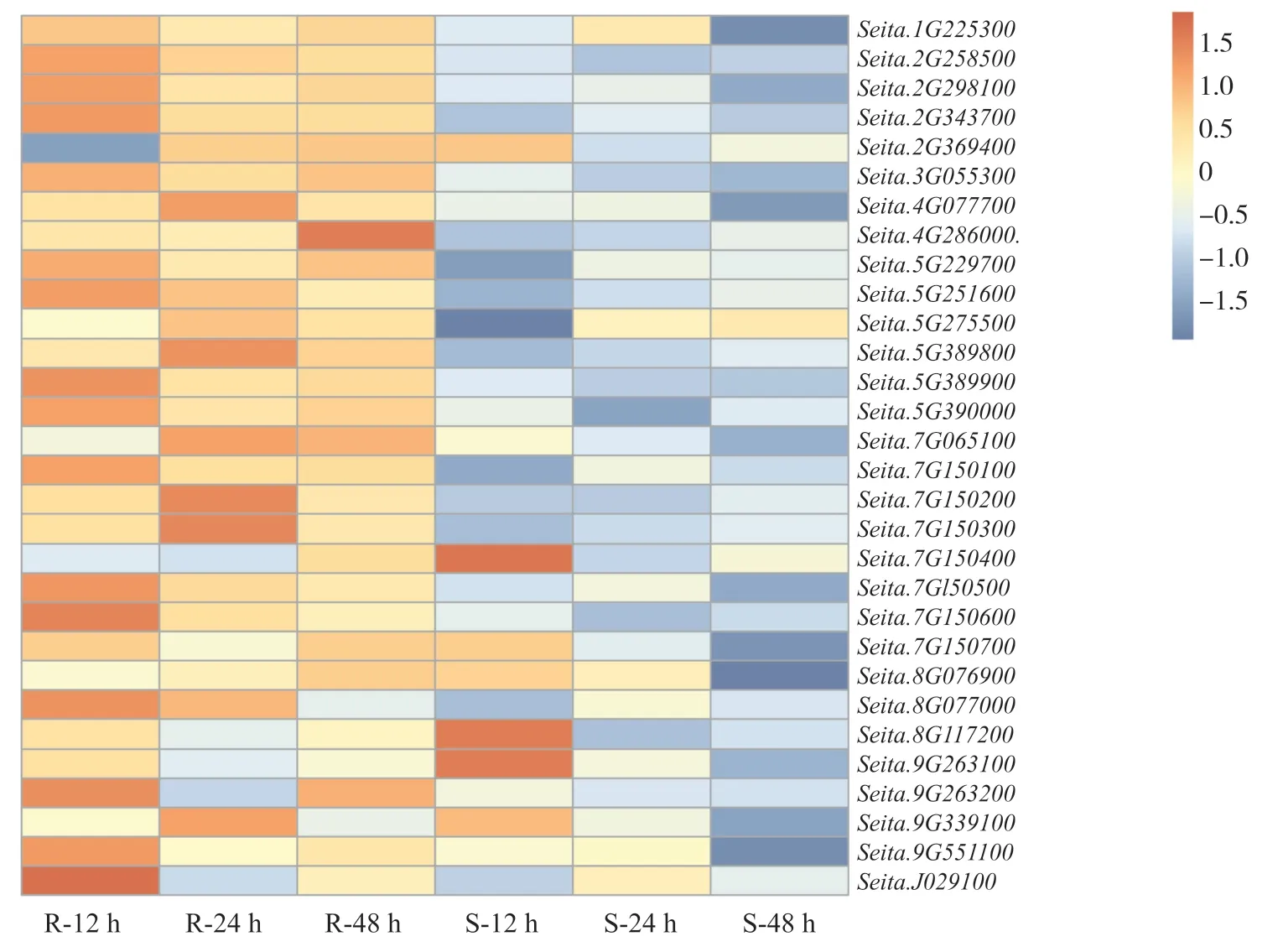
圖5 谷子幾丁質酶基因在抗感品種中3個時間段的表達分析Fig. 5 Expression analysis of chitinase gene in foxtail millet during three time periods in resistant cultivars
2.8 谷子幾丁質酶基因參與抗病響應的表達驗證
為進一步驗證幾丁質酶基因參與谷子抗白發病響應的表達差異,本文選擇Seita.1G225300、Seita.4G286000、Seita.5G389900、Seita.7G150600、Seita.7G150700和Seita.8G076900共6個基因進行實時熒光定量表達分析。通過驗證可知:Seita.1G225300在抗病品種中12、24和48 h 3個時間段均上調表達,而在感病品種中12 h時基因不表達;Seita.4G286000基因在抗病品種中48 h時表達量達到最高,而在感病品種中在這3個時間段內均呈現逐漸下降的趨勢;Seita.5G389900在抗病品種中3個時間段表達量上調,而在感病品種中表達量下調;Seita.7G150600在3個時間段內表達量明顯上調,而在感病品種中表達量較抗病品種均下調;Seita.8G076900基因在感病品種中48 h時顯著下調。
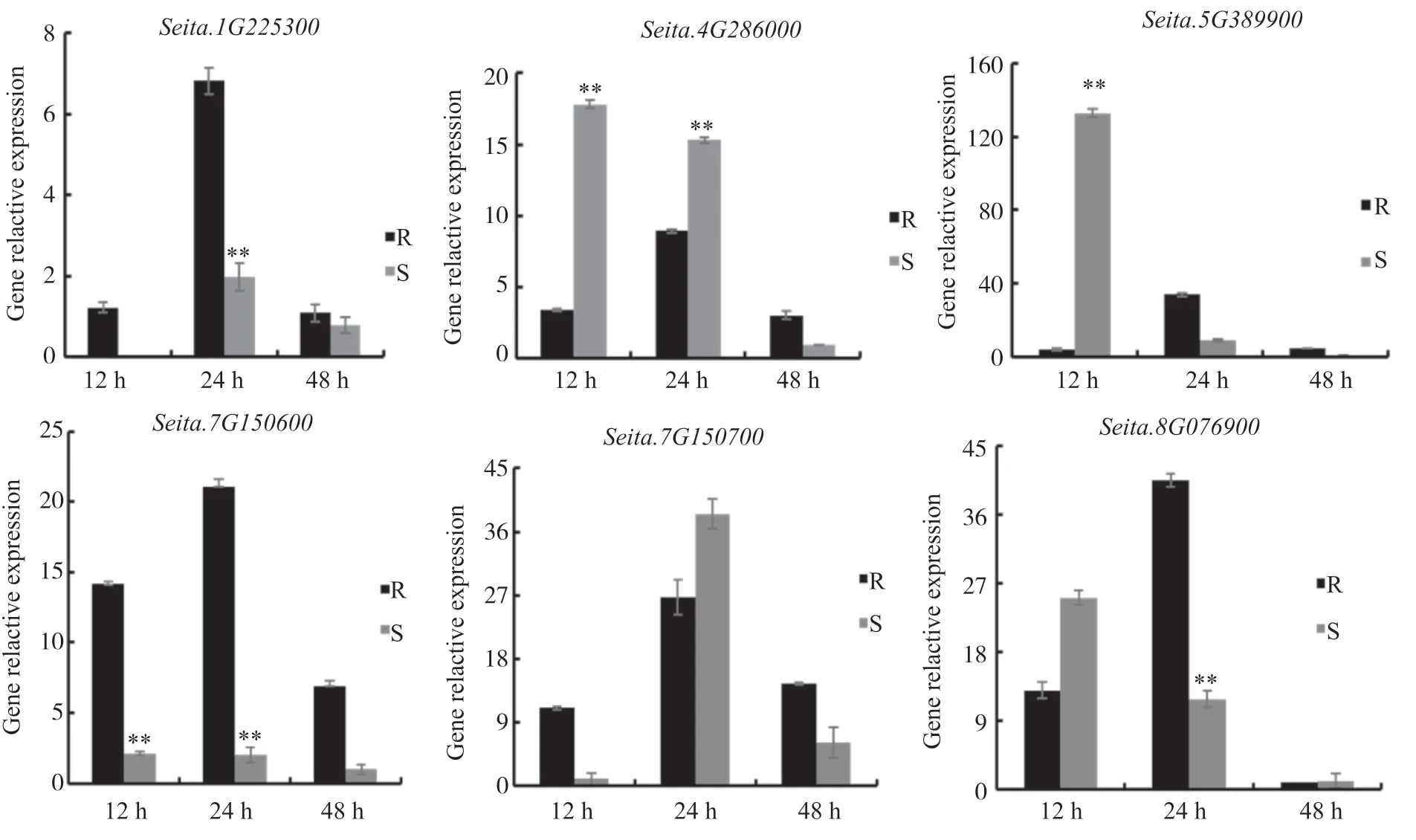
圖6 谷子幾丁質酶基因在白發病侵染后脅迫下的表達分析Fig. 6 Relative expression patterns of chitinase genes under adversity stress in foxtail millet
3 討論
幾丁質酶基因已在水稻[34]上被證明可參與調控植物的生長發育、抗逆性等過程。本研究對谷子中的幾丁質酶基因進行結構以及氨基酸保守結構域分析,發現其基因結構上表現出了一定的變異性,這與水稻和高粱中的結果相似[35]。對30個幾丁質酶基因進行染色體定位分析發現,分布在同一條染色體上的幾丁質酶基因具有相似的保守結構域。定位在第7條染色體上的幾個基因具有相似的Motif及相似的UTR及CDS區域。根據相似的結構域推測其具有相似的功能(圖3)。保守結構域分析顯示,幾丁質酶基因家族具有相同或相似的結構域,證明了幾丁質酶基因在進化過程中存在一定的保守性。
植物幾丁質酶基因的表達形式可以分為兩種:組成型表達和誘導型表達。植物幾丁質酶廣泛分布于植物的根、莖、葉、花及胚等部位。在外源因子的刺激下,植物體內的幾丁質酶大幅上調表達。本研究根據系統發育樹及染色體定位分析發現,同一條染色體上的基因在系統發育樹的分類相同,且有相似的保守結構域,結合熱圖分析可知,它們在抗病或抗旱過程中具有相似的調控方式(圖3~5)。幾丁質酶基因家族在植物抗病過程中發揮了重要作用[35]。在甘藍型油菜黑脛病和菌核病過程中,一些幾丁質酶基因家族成員被特異地誘導表達[36]。另外,荔枝的果皮幾丁質酶參與霜疫霉菌的抗病過程[37]。而本研究發現,Seita.7G150700和Seita.8G076900兩個基因共同參與抗旱和抗病過程。在這兩個過程中,抗病品種表達上調,而在感病過程中表達下調,且與轉錄組的結果一致,說明這兩個基因可能同時參與抗病和抗旱的過程。
本研究通過對其染色體定位、理化性質、基因結構與保守結構域進行分析,明確谷子幾丁質酶基因在抗感品種中的表達模式。其研究結果可為后期研究相關幾丁質酶基因對谷子的抗病性提供一定的理論基礎,為谷子分子育種發掘潛在的基因資源。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06