雞皮膚毛囊生長差異基因的生物信息學分析
2022-09-22 05:15:54劉祎軒朱慧媛張朋威雷艷茹高超群李東華康相濤
中國畜牧獸醫 2022年9期
關鍵詞:信號
張 豪,劉祎軒,朱慧媛,張朋威,雷艷茹,高超群,李東華,康相濤
(河南農業大學動物科技學院,農業農村部畜禽資源(家禽)評價利用重點實驗室,鄭州 450000)
在現代商業生產中,家禽羽毛的數量和質量越來越受到消費者的關注,羽毛生長速度、質量和脫毛方式對高價值家禽胴體的生產具有重要意義。在家禽規模化生產中,15%的雞存在不同程度的羽毛發育問題,如脫毛和身體覆蓋不足等[1-2],這些問題嚴重影響了雞的能量代謝、活體外觀和胴體質量。羽毛生長不良不僅影響活體外觀,而且降低胴體質量和飼料轉化效率[3-4]。羽毛是一種起源于外胚層表皮細胞的角質化衍生物[5],其生長發育的調控機制目前仍不清楚。羽毛發育始于皮膚毛囊,它是一個由上皮(表皮)和間充質(真皮)成分組成的復雜微器官,通過上皮和間充質之間的關鍵分子相互作用調控羽毛的生長、替換及其形態結構的變化[6-7]。在羽毛形態發生前真皮細胞的分化過程并沒有被預先決定,胚胎皮膚間充質通過與發育中的表皮相互作用,使表皮基板下的間充質細胞采用出芽或芽間分化的方式形成真皮凝聚物,凝聚物在隨后的形態發生過程中再進一步形成毛乳頭[8-9]。在胚胎發育早期,皮膚上皮和間充質可以通過細胞的緊密排列和一系列基因表達的變化來相互作用形成羽毛原基,羽毛原基進一步分化,開始在羽毛芽的不同部位表達不同的信號、生長和細胞黏附分子,然后在毛囊內長成細絲,通過孵化產生羽絨[9]。因此,探究家禽羽毛形態發生發育過程中上皮和間充質的相互作用,了解家禽羽毛生長發育的分子機制,對于促進羽毛生長進而提高家禽養殖業的經濟效益具有重要意義。
在雞胚胎期前6 d,雞胚皮膚發育不全,表皮層非常薄,很難區分皮膚的表皮和間充質結構[10]。在雞胚胎期第7天,胚胎背部從肱到腰骶出現羽芽,腰骶水平處有約7排羽芽,腿部出現明顯的羽芽,而在尾巴的每個側邊上羽芽較模糊。在雞胚胎期第9天時,羽芽更加明顯,新出現的羽芽靠近中腹線,與胸骨相連,并延伸至腹部兩側[11]。在雞胚胎發育的第7~9天是雞皮膚上皮和間充質調控羽毛毛囊發育的關鍵時期。雞胚胎期第7天首次產生的背部羽毛在形態上被鑒定,第7~9天突出的毛囊建立了良好毛細血管供應,并滲透到初期毛囊的毛乳頭中[12],血管系統的建立對毛囊的后期生長至關重要。本研究通過下載GSE62882芯片表達數據集,經生物信息學分析篩選雞胚胎期第7~9天影響羽毛生長發育的關鍵基因及其參與的生物過程和信號通路,為進一步探究皮膚上皮和間充質在毛囊生長發育中的作用提供理論參考。
1 材料與方法
1.1 基因芯片數據分析
GSE62882芯片表達數據集以Affymetrix基因組陣列GPL3213平臺為基礎,從GEO數據庫(https:∥www.ncbi.nlm.nih.gov/geo/)中下載獲得,本研究中共有8個Cobb500肉雞樣本數據,其中胚胎期第7天背部皮膚上皮(E7 dorsal skin epithelium,E7ep)、胚胎期第9天背部皮膚上皮(E9 dorsal skin epithelium,E9ep)、胚胎期第7天背部皮膚間充質(E7 dorsal skin mesenchyme,E7me)和胚胎期第9天背部皮膚間充質(E9 dorsal skin mesenchyme,E9me)樣本數據各2個(表1)。將原始表達矩陣通過R語言中的limma包進行標準化分析,以log2|FoldChange|>1且P<0.05為篩選條件,篩選出E7ep和E9ep、E7me和E9me的差異基因,并繪制火山圖及熱圖。
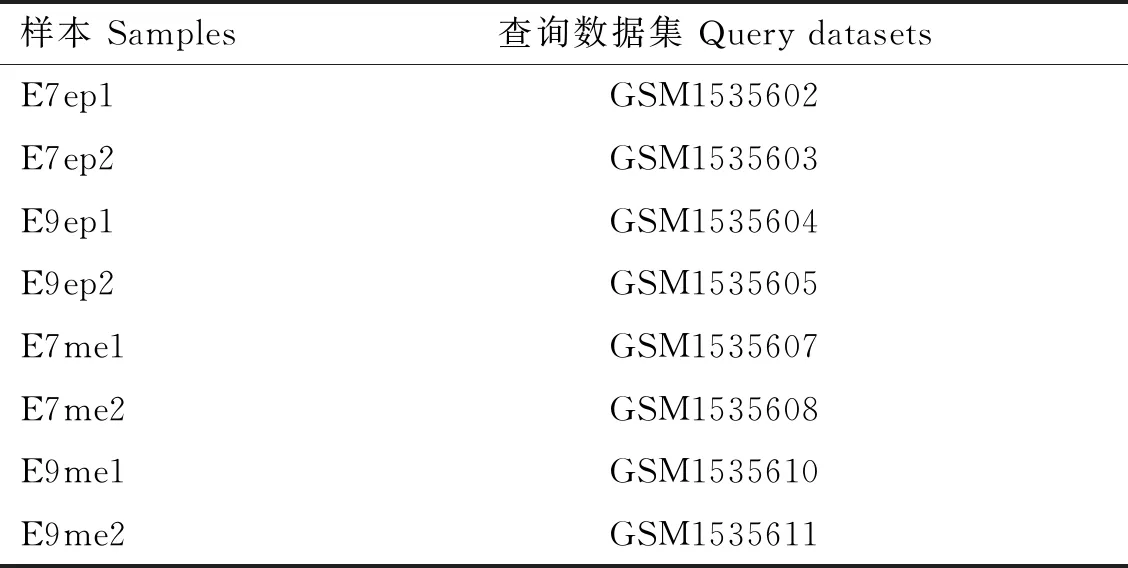
表1 GSE62882樣本數據
1.2 關鍵基因篩選
將E7ep和E9ep、E7me和E9me差異基因,以及E7ep/E9ep、E7me/E9me差異基因中共同上下調基因分別導入STRING version 11.5在線軟件(https:∥string-db.org/),得到蛋白互作(protein-protein interaction,PPI)網絡,同時將PPI網絡導入Cytoscape 3.8.2軟件進行可視化。運用STRINGAPP插件計算PPI網絡中各節點的拓撲參數,基于Degree選擇PPI網絡排名前十的關鍵基因,找出網絡中處于中心位置的關鍵節點,并構建PPI子網絡圖。
1.3 差異基因GO功能注釋
GO功能富集分析是大規模基因功能注釋的常用方法,本研究取E7ep和E9ep、E7me和E9me的差異基因運用R語言中clusterProfiler程序包進行功能注釋,獲取差異基因的GO條目。
1.4 差異基因的信號KEGG通路富集分析
KEGG是一個包含生物化學路徑的生物信息學數據庫。本研究將E7ep和E9ep、E7me和E9me的差異基因,以及表皮與間充質中共同上下調基因,利用R語言中clusterProfiler程序包進行差異基因KEGG通路富集分析,獲取差異基因和共同上下調基因富集的信號通路。
2 結 果
2.1 E7ep和E9ep差異基因的篩選和PPI網絡的構建
本研究共篩選出E7ep和E9ep差異基因626個,其中上調基因356個,下調基因270個(圖1)。選取前25個差異基因繪制熱圖,發現E7ep和E9ep差異基因包括SPINK5、LRRC17、GPIHBP1等(圖2);將差異基因導入STRING數據庫進行PPI網絡分析,經Cytoscape 3.8.2軟件構建PPI網絡的可視化結果,剔除孤立蛋白后,共得到119個節點及189條互作關系(圖3),基于Degree選擇PPI網絡排名前十的核心基因分別為WNT3A、FN1、WNT7A、FZD8、WNT3、WIF1、SHH、WNT9B、WNT4、WNT6(圖4)。
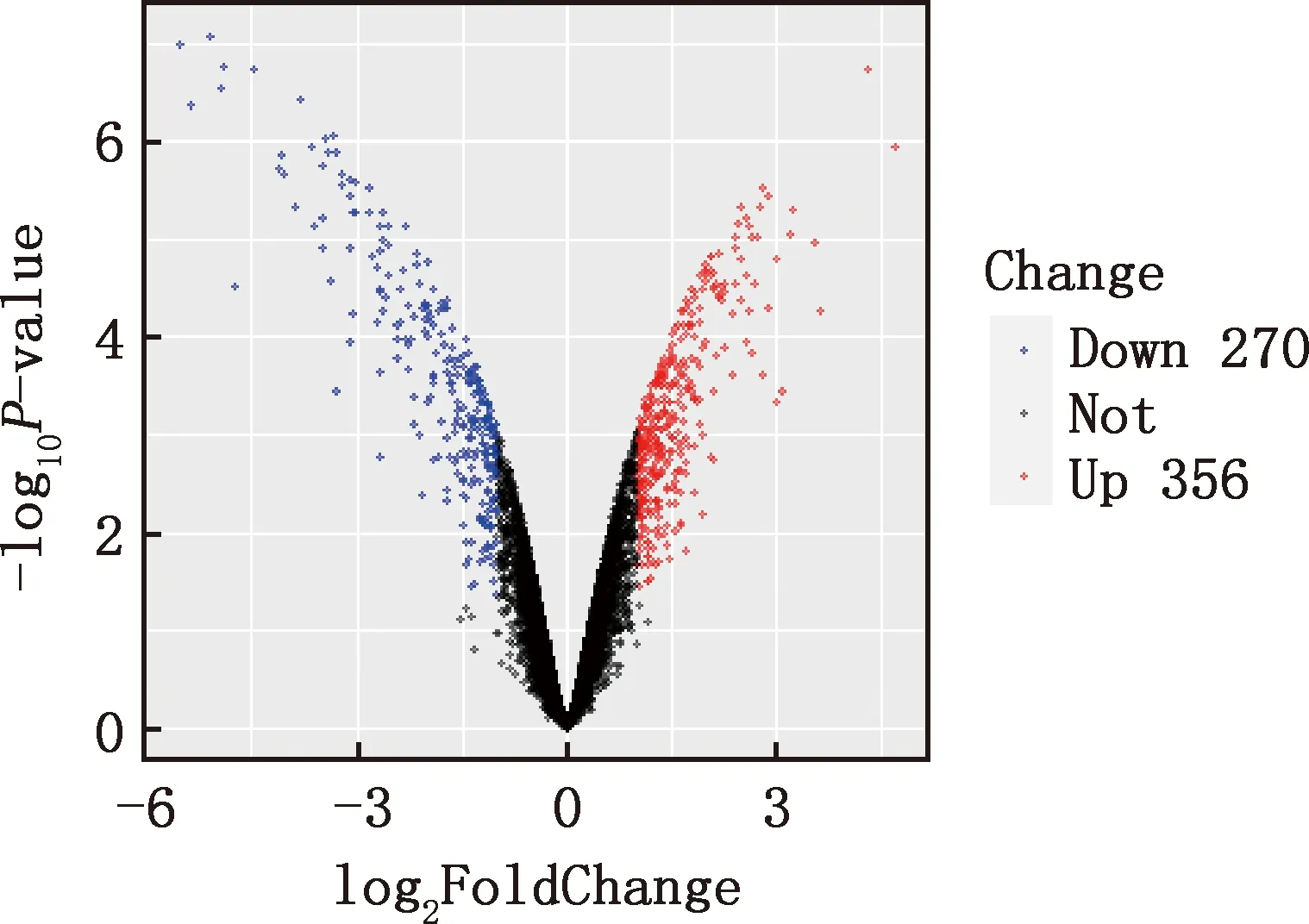
圖1 E7ep和E9ep差異基因的火山圖
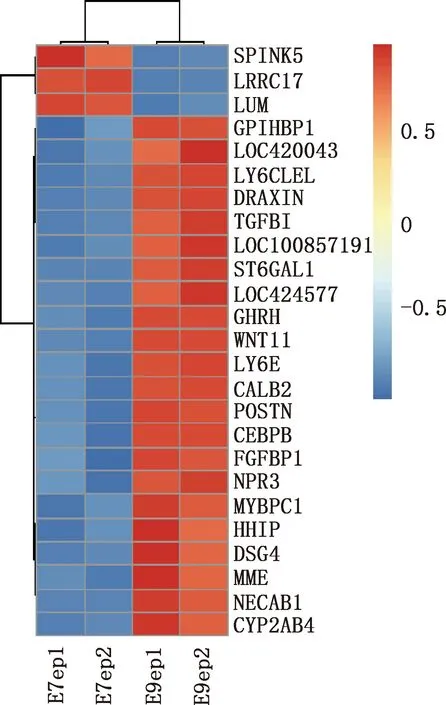
圖2 E7ep和E9ep差異基因的熱圖
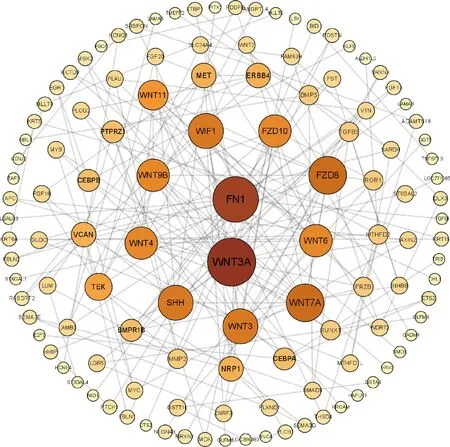
圖3 E7ep和E9ep差異基因的PPI網絡分析

圖4 E7ep和E9ep差異基因的PPI網絡樞紐模塊分析
2.2 E7ep與E9ep差異基因的GO功能注釋和KEGG通路富集分析
對E7ep和E9ep差異基因進行GO功能注釋和KEGG通路富集分析,結果顯示,差異基因參與細胞分化、解剖結構形態發生、神經系統發育、組織發育、神經形成、神經元的形成、神經分化、細胞黏著、生物黏附、多細胞有機體負調控等GO條目(圖5),參與Wnt信號通路、黑色素合成、TGF-β信號通路、mTOR信號通路等信號通路(圖6)。

圖5 E7ep和E9ep差異基因的GO功能注釋
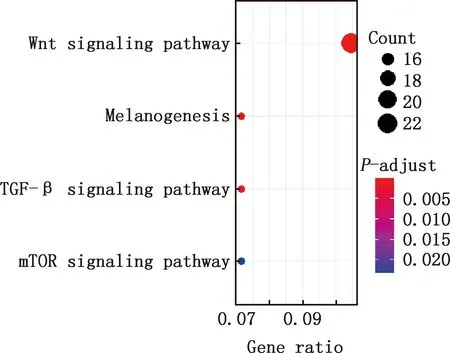
圖6 E7ep和E9ep差異基因的KEGG通路富集分析
2.3 E7me和E9me差異基因篩選和蛋白互作網絡構建
本研究共篩選出E7me和E9me差異基因690個,其中上調基因269個,下調基因421個(圖7)。選取前25個差異基因制備熱圖(圖8),發現E7me和E9me差異基因包括TAC1、FRK、PENK等;將差異基因導入STRING數據庫網站進行PPI網絡分析,經Cytoscape 3.8.2軟件構建PPI網絡的可視化結果,剔除孤立蛋白后,共得到206個節點及326條互作關系(圖9),基于Degree選擇PPI網絡排名前十的核心基因分別為FGF2、SDC1、ITGB3、BMP4、EGFR、ITGA9、FLT1、MYL10、RUNX2、PTPN11(圖10)。
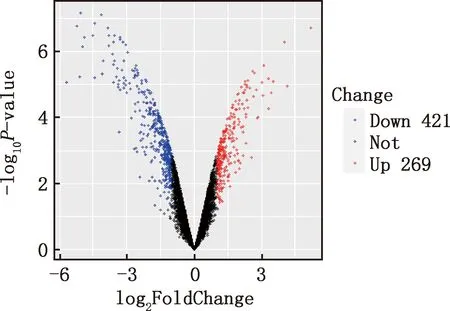
圖7 E7me和E9me差異基因的火山圖
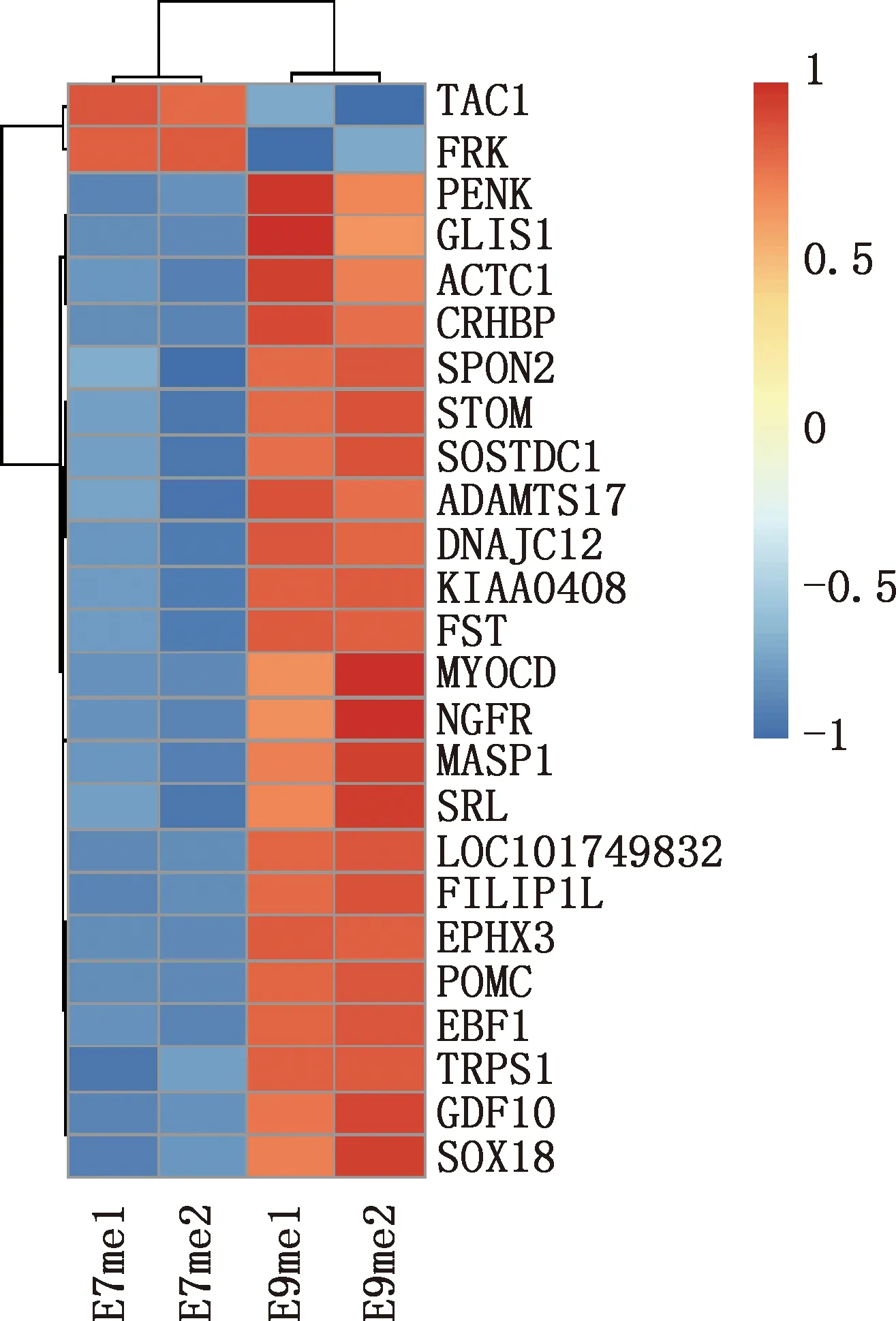
圖8 E7me和E9me差異基因的熱圖

圖9 E7me和E9me差異基因的PPI網絡分析
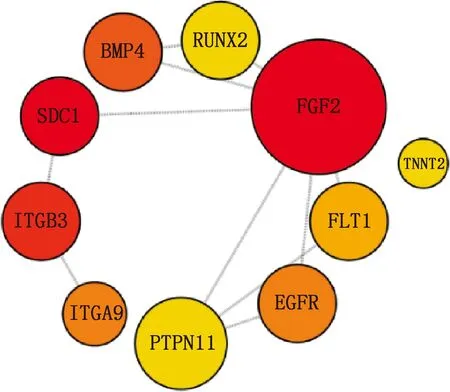
圖10 E7me和E9me差異基因的PPI網絡樞紐模塊分析
2.4 E7me和E9me差異基因的GO功能注釋和KEGG通路富集分析
對E7me和E9me差異基因進行GO功能注釋和KEGG通路富集分析,結果顯示,差異基因參與細胞分化、細胞發育過程、解剖結構的形態發生、動物器官發育、細胞表面受體信號通路、系統進程、循環系統的發展、解剖結構形態發生的調控、心臟發育、肌肉系統過程等GO條目(圖11),參與鈣信號通路、肌動蛋白骨架調控、EMC受體互作等信號通路(圖12)。
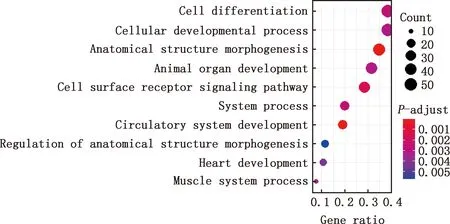
圖11 E7me和E9me差異基因的GO功能注釋
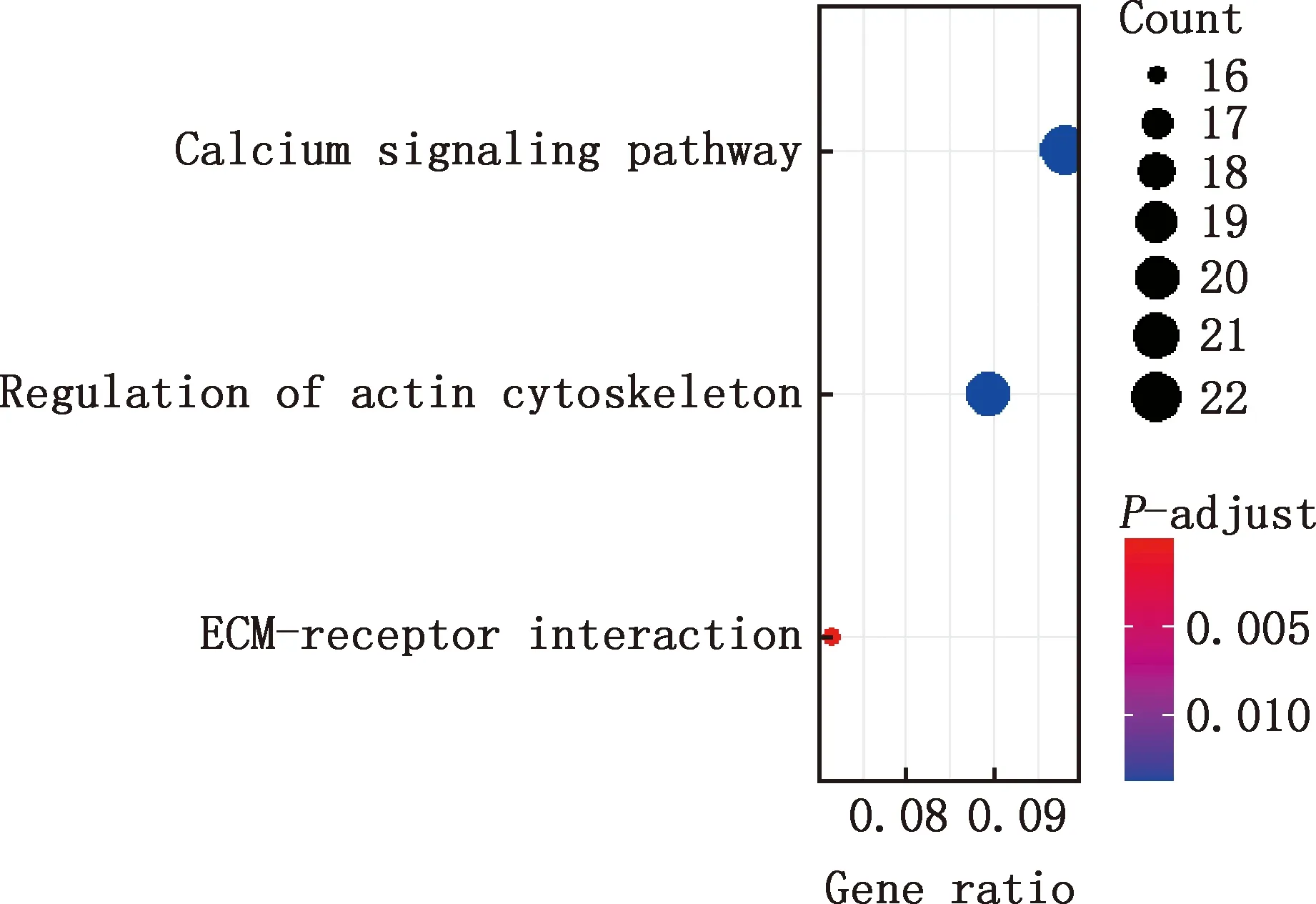
圖12 E7me和E9me差異基因的KEGG通路富集分析
2.5 皮膚上皮和間充質中共同上下調基因的生物信息學分析
篩選雞胚胎從第7和9天發育過程中背部皮膚上皮和間充質共同上下調基因,獲得共同上調基因26個(圖13A),共同下調基因48個(圖13B)。將共同上下調基因導入STRING數據庫進行PPI網絡分析,通過Cytoscape 3.8.2軟件構建PPI網絡的可視化結果。剔除孤立蛋白后,PPI網絡中共得到32個節點及34條相互作用關系,主要基因包括MYC、FGF18、RUNX2、JAG1、MSX2等(圖14)。將共同上下調基因進行KEGG通路富集分析,結果發現共同上下調基因主要富集在Wnt信號通路上(圖15)。
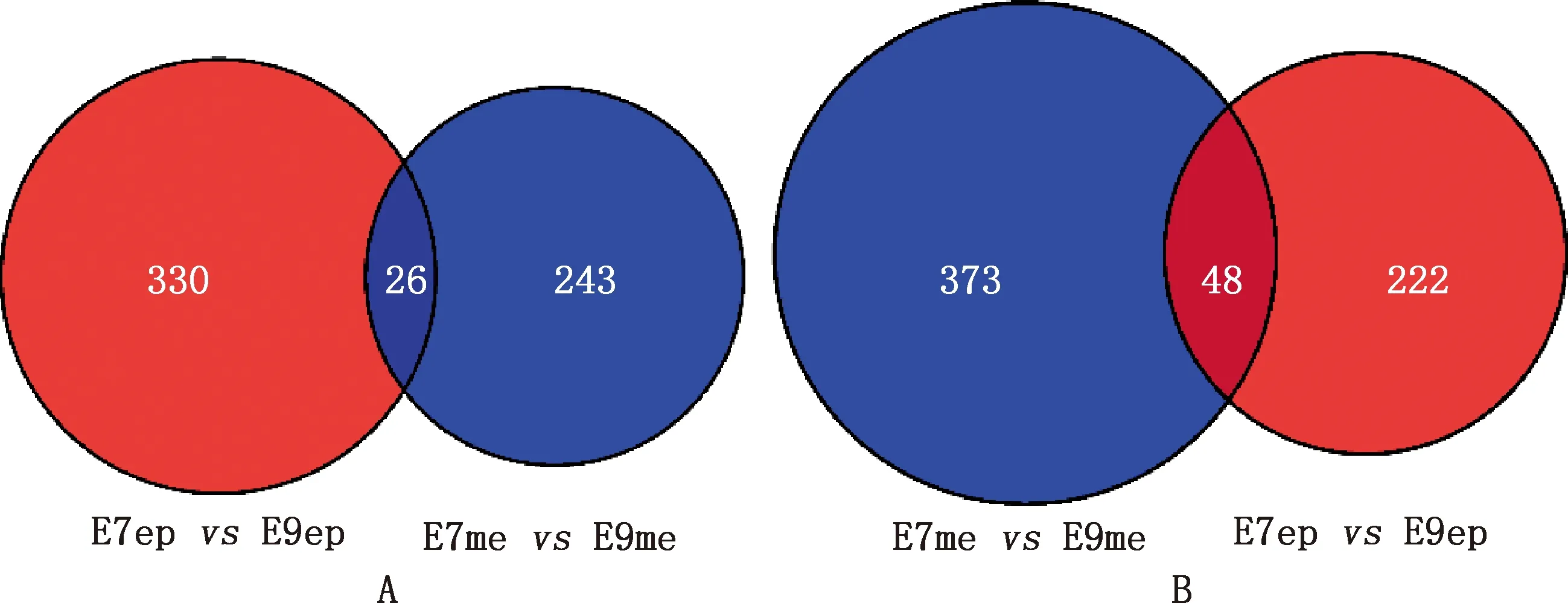
圖13 皮膚上皮與間充質共同上調(A)和下調(B)基因

圖14 共同上下調基因的PPI網絡分析

圖15 皮膚上皮與間充質共同上下調基因的KEGG通路富集分析
3 討 論
羽毛是雞的一個重要的外觀性狀,成雞羽毛的發育呈現從生長期、退行期到休止期的周期性生長,羽毛的生長性狀能夠反映雞的發育狀況,羽毛的正常生長和再生對提高家禽的福利和經濟價值具有重要意義[13-14]。羽毛是由皮膚上皮和間充質之間的一系列信號通路相互作用而形成的,上皮組織中的毛囊干細胞接收來自真皮間充質細胞的信號,并將信號傳回真皮間充質細胞,從而影響皮膚附屬物毛囊的生長發育[13]。目前關于雞胚胎皮膚毛囊發育的研究已有大量報道,但對早期毛囊發育過程中上皮和間充質間相互作用調控毛囊的生長發育的研究較少,本研究分析GSE62882數據集中胚胎期第7、9天的背部皮膚上皮和間充質的差異表達基因,篩選雞胚胎第7~9天發育過程中影響雞羽毛生長發育的關鍵基因、生物過程和信號通路,探究皮膚上皮和間充質協同作用對毛囊生長發育的影響。
本研究共獲得E7ep和E9ep差異基因626個,這些差異基因存在189條互作關系,基于Degree獲得PPI網絡排名前十的核心基因分別是WNT3A、FN1、WNT7A、FZD8、WNT3、WIF1、SHH、WNT9B、WNT4、WNT6,主要集中在Wnt信號通路上,通過GO功能注釋和KEGG通路富集分析發現,E7ep和E9ep差異基因主要富集在血管系統發育相關的生物過程和Wnt、TGF-β信號通路上。在個體皮膚附屬物中,WNT幫助形成局部生長區域,并形成發育中的羽毛,如WNT6過表達誘導不同羽毛區域的局部生長,WNT3A擴大了羽毛道的大小、促進了個體羽毛的形成[15-16]。TGF-β信號通路中最重要的成員是TGF-β超家族,該家族包含大量不同的多肽形態發生因子,包括TGF-β本身及BMP和GDF,能夠誘導毛囊進入退行期,如在小鼠中研究發現,TGF-β2通過激活IFN-γ和BDNF誘導毛囊進入退行期[17]。在毛囊生長發育初期,毛囊干細胞的激活需要抑制TGF-β信號通路中BMP的表達和激活Wnt信號通路中β-catenin的穩定表達共同完成。Wnt、TGF-β信號通路對血管的生成具有重要的調控作用[15,18]。在雞胚胎第7~9天的發育過程中,胚胎皮膚上皮開始有血管和神經的生成,毛囊周圍血管可提供毛囊生長所需的營養物質及各類細胞調控因子,進而調控毛囊細胞的生長分化[19]。
本研究共獲得E7me和E9me差異基因690個,基于Degree獲得PPI網絡排名前十的核心基因分別是FGF2、SDC1、ITGB3、BMP4、EGFR、ITGA9、FLT1、MYL10、RUNX2、PTPN11。活化的內皮細胞在FGF2的作用下開始增殖,并形成新的芽[18],使細胞間的接觸松動,導致毛細血管融合形成動脈和靜脈。FGF2可以通過誘導雞皮膚形成致密的真皮組織,致密真皮組織進一步發育成羽芽[20]。在表皮發育過程中,BMP信號傳導是皮膚上皮和間充質相互作用過程中的關鍵途徑,具有抑制毛囊生長發育的作用[13,21]。研究發現,EGFR抑制劑能夠縮短羽芽之間的距離并增加羽芽的數量[22];小鼠缺乏EGFR將不能長出毛發,小鼠皮膚中缺失EGFR將導致基質細胞有絲分裂活性增加、毛囊凋亡和形成毛發的上皮細胞系分化受損[23]。但EGFR在禽類皮膚毛囊生長發育過程中是否有相似的作用仍需進一步研究。E7me和E9me差異基因主要涉及肌肉發育、循環系統的發育和鈣信號通路,鈣信號通路能夠影響皮膚間充質細胞的運動方向,在加速傷口愈合、減少疤痕形成、傷口收縮和誘導毛發再生等過程中有廣泛的應用[24]。
本研究共獲得胚胎期第7和9天雞背部皮膚上皮和間充質中共同上調基因26個,共同下調基因48個,共同上下調基因PPI網絡中的主要基因包括MYC、FGF18、JAG1等。MYC在人和小鼠毛囊周期中已有報道,參與小鼠毛囊形態發生的不同階段及毛囊角化細胞的增殖或分化[25-26],c-Myc蛋白是β-catenin下游的轉錄因子,在雞胚胎期第8~12天表達量顯著增加,在調節毛囊干細胞分化中是必不可少的[27]。FGF18在小鼠毛囊中高表達,可誘導休止期毛囊再生,在雞羽毛生長中的作用目前仍不清楚[28]。JAG1是Notch的配體,通過Notch信號通路參與毛囊的生長調控[29-31]。Notch信號通路被認為是調節包括毛囊在內的多細胞生物干細胞穩態和發育的主要信號轉導通路。在毛囊發育過程中,Notch1的活性是決定細胞命運的關鍵。Notch1在早期基板的形成中并不發揮作用,但與真皮內陷毛乳頭的形成有關。Notch1敲除以后將會導致退行期提前,毛發逐漸變短[32-33]。對背部皮膚上皮和間充質共同上下調基因進行KEGG通路富集分析,結果發現共同上下調基因主要富集在Wnt信號通路上。Wnt/β-catenin信號通路在胚胎期第8~12天能夠促進羽毛的生長和發育[34-35]。Wnt信號通路在背部皮膚上皮和間充質互作中具有重要作用,在皮膚羽毛形成的誘導期,毛囊的形成依賴于單層上皮和真皮細胞濃縮物之間的一系列相互作用,上皮細胞接收來自底層間充質的指示信號,而間充質細胞需要表皮信號來形成毛乳頭。在毛囊基板形成之前,真皮間充質細胞均勻地接受廣泛的上皮發出的Wnt信號,真皮間充質細胞隨后產生第一個真皮Wnt信號,誘導表皮基底細胞定期聚集,導致毛發基板的形成,初始誘導后,發展中的基板產生Wnt配體,以誘導底層間充質細胞形成真皮凝聚物[36],最終形成皮膚毛囊的毛乳頭,而毛乳頭是毛囊生長發育的調控中心。在胚胎期第7~9天發育過程中,E7me和E9me差異基因在Wnt信號通路上并沒有富集,但是上皮和間充質共同上下調基因富集在Wnt信號通路上,這可能與分析過程中數據樣本的選取有關,并不代表E7me和E9me差異基因在Wnt信號通路上沒有富集。
4 結 論
本研究通過對GSE62882的數據集中部分數據進行生物信息學分析,結果表明在胚胎期第7~9天上皮和間充質共同上下調基因主要富集在Wnt信號通路上,表明上皮和間充質通過Wnt信號誘導皮膚毛囊的生長發育,為進一步探究皮膚上皮和間充質在毛囊生長發育中的作用提供理論參考。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
