淡水養(yǎng)殖太平洋鮭饑餓后補(bǔ)償性生長效果的研究
2022-10-22 03:00:36王正梅
黑龍江水產(chǎn) 2022年5期
關(guān)鍵詞:生長
王正梅
(桐城市水產(chǎn)服務(wù)中心,安徽桐城 231400)
在自然界中動(dòng)物常因季節(jié)變化、食物種類、不同的空間分布和環(huán)境劇變等因素時(shí)常會(huì)遭受饑餓的脅迫[1]。有些動(dòng)物在遭受饑餓脅迫后的一段時(shí)間再恢復(fù)原先的攝食時(shí),其生長的速度將會(huì)超過之前正常攝食的個(gè)體,這種現(xiàn)象就稱為補(bǔ)償生長[2]。太平洋鮭(Oncorhynchusspp.)具有豐富的營養(yǎng)價(jià)值,含較高的蛋白質(zhì)和不飽和脂肪酸,是目前主要養(yǎng)殖的冷水性魚類中的一種[3]。太平洋鮭喜歡生活在清涼、流動(dòng)的無污染、含氧豐富的水域中,可以在淡水中養(yǎng)殖,也可以在海水中養(yǎng)殖[4],其最適宜的生長水溫為8℃~18℃[5]。中國沿海海水因溫度高于鮭魚的適宜生長溫度,不適合規(guī)模化的飼養(yǎng)該魚[6]。目前太平洋鮭已經(jīng)在中國的內(nèi)陸部分水溫較低的湖泊、江河及水庫中形成規(guī)模化養(yǎng)殖。其中,安徽省桐城市就是一個(gè)適合太平洋鮭養(yǎng)殖的地方,因?yàn)榧竟?jié)變換,在寒冷環(huán)境下很多水中的生物會(huì)減少,對(duì)于太平洋鮭來說只要水不結(jié)冰就不會(huì)影響其正常攝食,但是低溫環(huán)境下則會(huì)影響其食物的生存,使得太平洋鮭面臨饑餓脅迫,所以研究太平洋鮭饑餓脅迫之后正常采食是否能達(dá)到補(bǔ)償生長對(duì)安徽省桐城市的太平洋鮭養(yǎng)殖具有重要意義[7]。文章針對(duì)太平洋鮭饑餓脅迫后補(bǔ)償生長進(jìn)行研究,旨在為該地區(qū)太平洋鮭的養(yǎng)殖提供參考意見。
1 材料與方法
1.1 試驗(yàn)材料
試驗(yàn)用的太平洋鮭購于安徽某鮭魚養(yǎng)殖場,體重均為245g±5.5g的太平洋鮭魚共54尾。
1.2 試驗(yàn)方法
將購買的54尾太平洋太平洋鮭先在體積0.3m3的水族箱中暫養(yǎng)8d,然后隨機(jī)分為6組,對(duì)照組、處理1組、處理2組、處理3組、處理4組和處理5組,每組9尾,分3個(gè)水平,每個(gè)水平3尾,一個(gè)水平的太平洋鮭放于一個(gè)水族箱,每一組的投喂方案詳見表1;每個(gè)處理組的太平洋鮭均飼喂同一種基礎(chǔ)日糧,基礎(chǔ)日糧配方及營養(yǎng)組分詳見表2;整個(gè)飼養(yǎng)試驗(yàn)為期72d;試驗(yàn)過程記錄鮭魚的始末體重及每日的飼料消耗量。

表1 試驗(yàn)分組及投喂方案

表2 試驗(yàn)鮭魚基礎(chǔ)日糧配方及營養(yǎng)組份
1.3 飼養(yǎng)管理
試驗(yàn)太平洋鮭每天分別于上午9:00和下午16:00進(jìn)行飼喂,每天飼喂2次,飼喂的量根據(jù)投餌量減去飽食后20mins搜集的烘干殘餌量;每天對(duì)水箱進(jìn)行換水,換水量為水箱總水的 1/3,同時(shí)保證水溫在8℃~18℃,每天定時(shí)對(duì)水箱內(nèi)的水進(jìn)行充氣12hrs。
1.4 試驗(yàn)指標(biāo)測定及數(shù)據(jù)處理
試驗(yàn)太平洋鮭的特定生長率、攝食率、食物轉(zhuǎn)化率和蛋白效率比指標(biāo)的計(jì)算參見馮健[8]等人所述;得出的試驗(yàn)指標(biāo)數(shù)據(jù)經(jīng)過Excel表格初步整理之后,利于SPSS20.0軟件進(jìn)行統(tǒng)計(jì)分析,結(jié)果均采用平均值±標(biāo)準(zhǔn)誤來表示,顯著水平為P<0.05,極顯著水平為P<0.01。
2 結(jié)果
由表3可以看出在飼養(yǎng) 72d后每一組的太平洋鮭的存活率均為100%,同時(shí)試驗(yàn)結(jié)束后,處理1組、處理2組和處理3組的太平洋鮭體質(zhì)量和對(duì)照組無顯著性差異(P> 0.05),可以完全補(bǔ)償因饑餓對(duì)生長產(chǎn)生的影響;處理4組、處理5組的末期魚體質(zhì)量和特定生長率極顯著的低于對(duì)照組(P< 0.01),說明該兩組的饑餓處理的補(bǔ)償飼喂每辦法達(dá)到不對(duì)太平洋鮭的正常生長造成影響;同時(shí)處理1、處理2、處理3組的攝食率、食物轉(zhuǎn)化率、蛋白效率高于對(duì)照組,處理4組和處理5組的攝食率、食物轉(zhuǎn)化率、蛋白效率極顯著的低于對(duì)照組(P< 0.01),說明處理1、2、3組的完全補(bǔ)償是因?yàn)轷q魚的食物轉(zhuǎn)換率和攝食率同時(shí)提高而起到的補(bǔ)償作用。
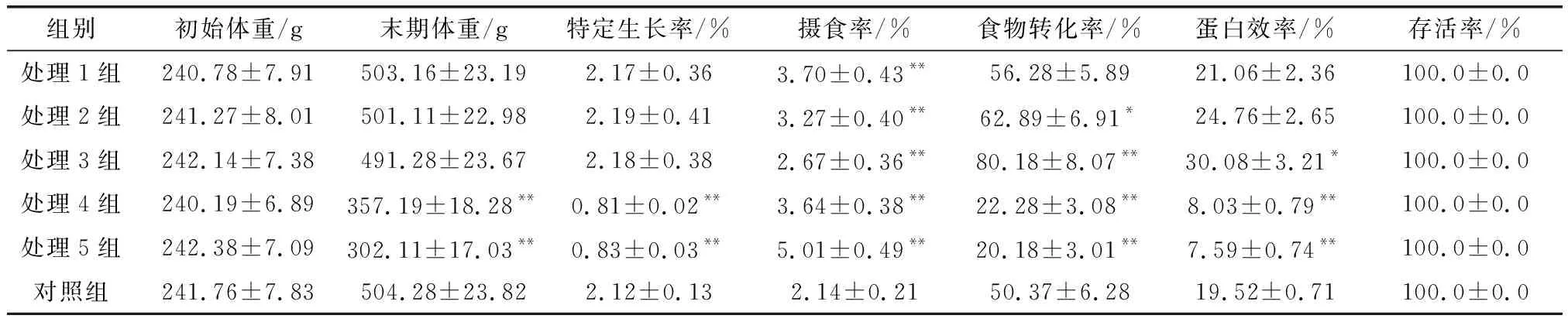
表3 太平洋鮭饑餓后補(bǔ)償對(duì)其生長效果的影響
3 結(jié)論
動(dòng)物產(chǎn)生補(bǔ)償生長主要是通過2個(gè)途徑,一是饑餓情況下使得動(dòng)物機(jī)體的代謝水平降低,但是在正常采食的時(shí)候,機(jī)體能維持一段時(shí)間的較低代謝水平,因?yàn)榇x支出的降低使動(dòng)物用于機(jī)體正常生長的能量增加,進(jìn)而提高了食物的轉(zhuǎn)化率,能夠?qū)?dòng)物進(jìn)行補(bǔ)償性生長;二是在饑餓后恢復(fù)正常采食,加大了動(dòng)物的食欲,一旦攝食水平提高了,動(dòng)物體內(nèi)就進(jìn)行大量的合成作用,完成補(bǔ)償性生長[9]。目前對(duì)于饑餓補(bǔ)償生長在魚類中的研究已經(jīng)很多,如鄭曙明等[10]研究發(fā)現(xiàn)對(duì)虎鯊饑餓處理4d、6d、8d時(shí),虎鯊部分出現(xiàn)補(bǔ)償生長,饑餓處理組雖然恢復(fù)生長后的生長速度高于對(duì)照組,但是其最終的體重不能趕上對(duì)照組。該研究在進(jìn)行72d的鮭魚養(yǎng)殖后,發(fā)現(xiàn)饑餓1d~6d在進(jìn)行補(bǔ)飼,太平洋鮭的生長速度明顯增快,且能趕上每天飼喂組的太平洋鮭體重,但是饑餓12d及以上在補(bǔ)飼,太平洋鮭的最終體重、特定生長率和對(duì)照組的相比表現(xiàn)極顯著的降低,說明饑餓12d及以上對(duì)太平洋鮭不能再進(jìn)行補(bǔ)償性生長,同時(shí)1d~6d饑餓組的最終體重低于對(duì)照組,趕不上對(duì)照組,該結(jié)論和上述論述相符;因此,在太平洋鮭養(yǎng)殖中, 應(yīng)盡量做到或者避免減少食物供應(yīng)不足本的情況發(fā)生;同時(shí)該研究還發(fā)現(xiàn)饑餓1d~6d循環(huán)補(bǔ)飼其攝食率和食物轉(zhuǎn)化率都顯著高于對(duì)照組,這說明完全補(bǔ)償生長的效果和是由攝食率、食物轉(zhuǎn)化率同時(shí)提高引起的該結(jié)論和Qian等[11]的銀鯽實(shí)驗(yàn)結(jié)論相符。
綜上所述,太平洋鮭飼養(yǎng)的時(shí)候盡量減少出現(xiàn)補(bǔ)償生長情況的出現(xiàn),避免不了的話,饑餓時(shí)間不要超過6d,以免對(duì)太平洋鮭的正常生長造成不可逆的影響。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
