蘿卜硫素下調結直腸癌中eIF4F翻譯起始復合物的表達
2022-11-03 07:22:02周大臣喻宗繁
安徽醫科大學學報 2022年10期
邢 運,周大臣,賀 良,崔 笑,喻宗繁
結直腸癌(colorectal cancer, CRC)是指結腸和直腸黏膜上皮和腺體發生的惡性腫瘤。2018年的全球癌癥統計結果表明,CRC總發病人數185萬,占比10.2%,居全球癌癥發病率第3位。死亡人數88.1萬(9.2%),居第2位[1]。在細胞翻譯過程的起始階段,真核細胞啟動因子4F(eIF4F)翻譯起始-復合物作為限速步驟發揮著重要作用[2]。蘿卜硫素(sulforaphane,SFN)是一種異硫氰酸酯,天然存在于大多數十字花科蔬菜中。1992年首次從西蘭花中提取,并定義為II相解毒酶誘導劑[3]。SFN已被證實具有多種抗癌特性,特別是對CRC[4]。該研究從CRC患者組織標本中檢測eIF4F翻譯起始復合物相關蛋白的表達,進一步探討SFN影響CRC細胞增殖的具體分子生物學機制,為SFN治療CRC提供理論依據。
1 材料與方法
1.1 材料
1.1.1組織標本 收集12例安徽醫科大學第二附屬醫院普外科CRC患者的組織標本,分別收取腫瘤組織與癌旁組織,放入裝有RNA組織保存液的標本管中,并且所有標本在收集和運輸過程中均置于冰上,并于-80 ℃冰箱中長期保存,將收集到的12例組織樣本制作為組織芯片。
1.1.2細胞與動物 原代CRC細胞系HT-29購于上海生命科學院細胞典藏庫。健康裸鼠,4~6周齡,18~22 g,購自江蘇南京集萃藥康生物有限公司,動物生產許可證號:SCKK(蘇)2018-0008。飼養于20~25 ℃、40%~60%濕度、照明與黑暗各12 h交替環境中,自由進水、飲食,1周后行后續實驗。
1.1.3試劑和儀器 SFN、MTT購自美國Sigma公司;胎牛血清購自美國Gibco公司;青霉素/鏈霉素、BCA試劑盒和4%多聚甲醛購自上海碧云天生物技術有限公司;DMEM購自美國Hycolne公司;0.25%胰酶購自美國Wisen公司;β-actin和eIF4F復合物相關抗體購自美國Cell Signaling Technology公司;山羊抗兔或抗鼠二抗購自北京中杉金橋有限公司;二甲基亞砜(DMSO)和0.1%結晶紫購自北京索萊寶科技有限公司。細胞培養箱購自美國 Thermo Fisher公司,熒光倒置顯微鏡購自日本OLYMPUS公司,酶標儀購自上海科華生物工程股份有限公司。
1.2 方法
1.2.1細胞培養 原代CRC細胞系HT-29細胞,DMEM高糖培養基+10%血清+雙抗(青霉素、鏈霉素)中培養,細胞培養箱培養條件為:5%CO2,37 ℃。每3 d換液1次,每5 d傳代1次,傳代次數不超過10代,1 500 r/min離心5 min,取生長狀態良好的處于對數生長期的HT-29細胞進行實驗。
1.2.2細胞活力測定(MTT) 將細胞接種于密度為3×103個/孔的96孔板中。二甲基亞砜(DMSO,對照組)和不同濃度的SFN處理48 h。MTT比色法測定細胞活力。在每個時間點加入5 mg/ml 的MTT溶液10 μl,37 ℃孵育4 h。移除培養基并添加200 μl異丙醇以溶解甲醛沉淀物。在570 nm處測量細胞勻漿的光密度,620 nm作為參比濾光片。DMSO作為載體對照,對細胞活力無影響。
1.2.3Western blot實驗 處理后的HT-29細胞PBS洗2次,加入蛋白裂解液(RIPA ∶蛋白酶抑制劑 ∶磷酸酶抑制劑比例為1 ∶100 ∶100),待細胞裂解完全后吸取懸液,4 ℃、13 200 r/min離心30 min,吸取蛋白上清液,儲存于-80 ℃冰箱,用BCA法測蛋白濃度并進行蛋白質定量,加上樣緩沖液煮沸10 min變性,以蛋白質30 μg進行SDS-PACE電泳。轉膜后,用含5% BSA的TBST室溫慢搖封閉1.5 h,分別加入β-actin和相應一抗(1 ∶1 000),4 ℃孵育過夜,加入相應的辣根過氧化物酶標記的羊抗鼠/兔二抗孵育1 h,洗膜后用顯影儀檢測蛋白顯影。
1.2.4動物實驗(裸鼠成瘤) 12只健康適齡裸鼠隨機均分為兩組,將0.1 ml(細胞數約1×106)處于對數生長期的HT-29細胞沿皮下緩慢注入裸鼠腋窩。3 d后,實驗組每天腹腔注射溶解于DMSO的SFN(0.5 mg/kg),對照組每天腹腔注射等濃度的溶劑DMSO(0.5 mg/kg),共注射14 d。14 d后處死裸鼠,取出腫瘤,測量體積大小,將腫瘤標本制成石蠟切片進行蘇木精-伊紅(HE)染色和免疫組織化學染色。
1.2.5HE染色法 石蠟切片置于65 ℃恒溫箱中烘焙60 min,于二甲苯中脫蠟,依次在無水乙醇、95%、70%、50%乙醇溶液中脫蠟處理。將切片依次放入蘇木精、伊紅染色槽中,用自來水洗滌,再置于50%、70%、95%乙醇溶液、無水乙醇中脫水處理,干燥后用中性樹脂密封,鏡下觀察細胞結構與形態。
1.2.6免疫組織化學染色法 石蠟切片置于65 ℃恒溫箱中烘焙60 min,于二甲苯中脫蠟,依次在無水乙醇、95%、70%、50%乙醇溶液中脫蠟處理。再放入枸櫞酸鈉溶液中高溫下抗原修復10 min。用純水沖洗2次,每次5 min,油性筆畫圈,10%過氧化氫去離子水 ∶甲醇(1 ∶9) 孵育10 min,以滅活內源性過氧化物酶活性,TBST沖洗3次,每次5 min,破膜液(含0.25% Triton X-100的TBS) 10 min,TBST沖洗,滴加10%山羊血清封閉液,室溫孵育2 h,滴加一抗,4 ℃過夜。回收一抗,TBST沖洗,滴加反應增強液,清洗,滴加相應辣根過氧化物酶標記的二抗,純水沖洗,DAB顯色后自來水沖洗終止反應,蘇木精染核20 s,純水沖洗。再置于50%、70%、95%乙醇溶液、無水乙醇中脫水處理、固定,中性樹膠封片后鏡下觀察相關蛋白表達情況。
1.3 統計學處理使用GraphPad Prism 9.0軟件對實驗數據進行統計學分析。細胞相關實驗均分別重復3次以上。用ImageJ軟件測量蛋白灰度值以及免疫組化光密度值,并對光密度(optical density,OD)值、裸鼠成瘤中的腫瘤體積結果進行t檢驗分析,P<0.05表示差異有統計學意義。
2 結果
2.1 CRC患者標本中eIF4F復合物相關蛋白在腫瘤組織中的表達高于癌旁組織CRC患者標本的免疫組化染色結果顯示,eIF4F復合物相關蛋白在腫瘤組織中的表達明顯高于癌旁組織(圖1A)。OD值分析顯示:腫瘤組織4EBP1(t=4.235,P<0.001)、P-4EBP1(t=2.974,P<0.01)、eIF4E(t=3.820,P<0.001)、P-eIF4E(t=3.308,P<0.01)、eIF4G(t=4.252,P<0.001)表達均高于癌旁組織,差異有統計學意義(圖1B)。

圖1 CRC患者腫瘤組織與癌旁組織中eIF4F復合物相關蛋白的表達差異 ×200A:腫瘤組織與癌旁組織免疫組化染色結果;B:各組OD值差異分析;與癌旁組織比較:**P<0.01,***P<0.001
2.2 SFN可抑制HT-29細胞的增殖用MTT比色法檢測SFN作用48 h后HT-29細胞存活率的變化,結果顯示:SFN對HT-29細胞活性有抑制作用,并且呈劑量依賴性。SFN對HT-29細胞的半抑制濃度(half maximal inhibitory concentration, IC50)為15.85 μmol/L(圖2A)。裸鼠成瘤實驗結果顯示,實驗組腫瘤體積低于對照組,差異有統計學意義(t=3.645,P<0.05)(圖2B)。HE染色結果顯示,與對照組比較,實驗組組織細胞內空泡更多(圖2C)。
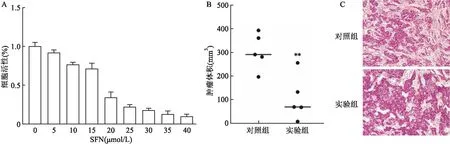
圖2 SFN抑制HT-29細胞的增殖 ×200A:不同濃度SFN處理HT-29細胞48 h后的細胞活性;B:裸鼠皮下移植瘤的體積比較;C:裸鼠皮下移植瘤的HE染色;與對照組比較:**P<0.01
2.3 SFN通過PI3K/AKT/mTOR/4EBP1信號通路影響HT-29細胞的增殖Western blot實驗結果顯示,SFN處理HT-29細胞后,eIF4F翻譯起始復合物相關蛋白的表達降低。作為其信號通路的上游,AKT和mTOR表達量也同時降低,P-AKT和P-mTOR的表達有所上升,而PI3K的表達量無明顯差異(圖3A)。裸鼠皮下移植瘤的免疫組織化學染色顯示,與對照組相比,實驗組中eIF4F翻譯起始復合物相關蛋白的表達量降低,AKT的表達降低,P-AKT與P-mTOR的表達增加(圖3B),這與Western blot檢測結果相吻合。

圖3 SFN降低相關通路蛋白及eIF4F復合物相關蛋白的表達 ×200A:SFN處理HT-29細胞后相關蛋白的Western blot結果;B:皮下移植瘤相關蛋白的免疫組化結果
3 討論
本研究顯示,eIF4F翻譯起始復合物在人類CRC組織中上調,這是翻譯起始的關鍵組成部分。同時,從十字花科蔬菜中提取的天然異硫氰酸酯SFN可通過抑制PI3K/AKT/mTOR/4EBP1信號通路,抑制原代CRC細胞中eIF4F復合物的上調。
mRNA翻譯(蛋白質合成)是基因表達中最耗能的步驟,并且受到高度調控。翻譯調控是細胞對環境的快速適應性反應,對維持細胞內蛋白質的穩態至關重要。因此,翻譯過程在人體生長發育中起著重要的作用,mRNA翻譯成蛋白質更是基因表達調控中的關鍵[5]。蛋白質翻譯包括起始、延伸、終止和核糖體回收。起始是限速階段,并受到eIF4F翻譯起始復合物合成速度的限制[6]。eIF4F由支架蛋白eIF4G、mRNA5′Cap結合亞單位eIF4E和RNA解旋酶eIF4A三個部分組成。在組成蛋白質翻譯起始復合物的所有起始因子中,由于eIF4E被認為是最不豐富的翻譯起始因子,所以,eIF4E 與eIF4G 結合形成eIF4F復合物的過程,也就成為了翻譯開始的限速步驟和控制蛋白質翻譯、表達的關鍵點[7]。
eIF4E:eIF4G的形成受到小的翻譯抑制因子4E結合蛋白(4EBPs)的嚴格調控,4EBPs與eIF4G競爭形成eIF4E:4EBP1復合物,并抑制eIF4E:eIF4G的形成[7]。eIF4E上4EBPs的結構結合位點與eIF4G重疊,從而使eIF4E能夠同時與eIF4G或4EBP1獨占結合。哺乳動物有3種4EBPs:4EBP1、4EBP2和4EBP3,其中4EBP1發揮著比其他因子更重要的生物學功能[8]。4EBP1與帽結合eIF4E的結合比與無帽eIF4E結合更快、更緊密。4EBP1和eIF4E之間的這種相互作用受到4EBP1磷酸化的嚴格調控,這反過來又會使4EBP1失活,并解離eIF4E:4EBP1相互作用。4EBP1的過度磷酸化在大多數人類癌癥中被觀察到,并且經常與腫瘤分級和不良預后相關。
文獻[9]報道,PI3K/AKT/mTOR通路在細胞增殖、凋亡、代謝和血管生成等方面都發揮著重要作用。另外,PI3K/AKT/mTOR信號通路與許多人類疾病密切相關,包括缺血性腦損傷、神經退行性疾病和腫瘤[10]。PI3K/AKT/mTOR分為兩部分:磷脂酰肌醇3-激酶(PI3K)及其下游分子絲氨酸/蘇氨酸蛋白激酶B(PKB,又稱AKT)。該通路被RTK和細胞因子受體激活。酪氨酸殘基被磷酸化,為PI3K轉位到膜上提供錨點,參與細胞外基質分子和細胞因子的轉導。這一信號通路對細胞也有重要的生物學效應,如促進細胞活性和延緩衰老和死亡,與各類生理活動相聯系。這一通路的功能障礙不僅與腫瘤的發生和發展有關,還與許多其他人類疾病有關,如白血病、糖尿病和精神分裂癥。雷帕霉素(TOR)是真核細胞絲氨酸/蘇氨酸激酶靶標,是細胞生長代謝的關鍵因子[11]。哺乳動物mTOR能感知細胞水平的代謝物(如氨基酸、葡萄糖、氧氣和生長因子),調節從蛋白質合成到自噬的一系列過程。并在不同的復合體中起作用,即mTOR復合體1(mTORC1)和mTOR復合體2(mTORC2)[12]。
本研究顯示,SFN可以通過抑制PI3K/AKT/mTOR/4EBP1信號通路來抑制eIF4F翻譯起始復合物的形成,其原因可能是mTOR磷酸化引起的信號反饋導致AKT過度磷酸化和4EBP1去磷酸化。反過來,4EBP1被SFN去磷酸化并激活,其與eIF4E穩定結合,導致蛋白停止翻譯。mTOR通過調節4EBP1的磷酸化在mRNA的翻譯過程中起著關鍵作用。然而,即使AKT磷酸化被激活,PI3K的表達仍無顯著變化。因此,可以發現mTOR磷酸化可誘導反饋,激活AKT磷酸化,影響其下游的4EBP1,從而影響蛋白質的翻譯過程。
